


Introduction
The hibernation of relatively small in size and body weight mammals includes two main components: prolonged numbness with a significant decrease in metabolism and body temperature and relatively short periods of the homeothermic state. While periodic awakenings, animals can either forage (hamsters and chipmunks), or do not eat at all (ground squirrels, marmots, hedgehogs), excrete from the body only liquid excrement, or both liquid and solid. There are a number of hypotheses with a certain degree of probability explaining the hibernation intermittence, however, the nature, role and ratio of endogenous and exogenous factors in the formation of hibernation rhythms have not been finally determined (Anufriev , 2008).
The range of external temperatures, within which hibernation is possible, is limited in hibernating mammals. For most hibernants, it ranges from -5 to 15°C (Slonim, 1971). For gophers, the upper limit of the environmental temperature at which hibernation is possible is at about 22°C; at higher temperatures, the hibernation state of does not occur. In marmots, this indicator is also about 22°C, in hamsters within 9–12°C, in bats it is above 22°C (Kalabuhov, 1929; Kayser, 1961; Pengelley, Fisher, 1957, 1963).
The duration of periods of hypothermia – bauts has intraspecific, interspecific and seasonal peculiarities (Strumwasser, 1959; Pengelley, Fisher, 1961; Ortmann, Heldmaier, 2000; Oklejewicz et al., 2001; Hut et al., 2002; Калабухов, 1985). For example, in a European hedgehog (Erinaceus europaeus), at the beginning of hibernation, the duration of bauts is 2–3 days, in the middle of hibernation – from two to three weeks. In gophers, the maximum duration of numbness periods is basically no more than two weeks, in marmots – two-three weeks, in smaller winter- hibernating rodents (hamsters, chipmunks) - within 7 days (Duboeus, 1898; Kayser, 1961; Galster, Morrison, 1970 ; Willis et al., 1971; French, 1985; Buck, Barnes, 2000; Oklejewicz et al., 2001). In the Arctic ground squirrel (Spermophilus parryii), the environmental temperature plays a leading role in the occurrence of hibernation among other external factors (Dresher, 1967). A relationship between hibernation depth and ambient temperature for copperheads, bats, four species of chipmunks of the genus Eutamias, European hedgehogs, European ground squirrels and alpine marmots was found (Kristoffersson, Soivio, 1964; Heller, Poulson, 1970; Ortmann, Heldmaier, 2000; Hut et al., 2002). The dependence of the duration of hypothermia periods on body temperature in hibernation was found in the copperhead (S. lateralis) in the temperature range from 2 to 22°C. With decrease in body temperature, the duration of numbness periods increased, the dependence was logarithmic (Twente, Twente, 1965, 1967).
The success of hibernation in hibernators and their survival depend on the depth and duration of the conditions with lowered body temperature during the winter period. At the same time, the mechanism of the influence of temperature on the formation of hibernation rhythms is not fully understood, since there are insufficient materials on the species-specific features of this phenomenon. The aim of this work was to study the course of hibernation of a ground squirrel under conditions close to natural, to identify the mechanisms of the influence of ambient temperature on the formation of hibernation rhythms.
Materials
The ground squirrels were caught in August, 80 km South of Yakutsk by filling holes by water. In September, the devices for long-term registration of body temperature (temperature drives, thermographs DS-1922L; a detailed description of the device and their main characteristics can be found on the websites: http://www.elin.ru, http://www.thermochron.ru/, http://www.ibdl.ru/) programmed to temperature measurements at a frequency of 1 time per 60 min. was implanted intraperitoneal. Wintering and hibernation of four individuals of ground squirrels (Spermophilus undulatus Pallas, 1778) from November to April took place in the underground congelation laboratory of the Institute of geocryology of the SB RAS (Yakutsk). The tunnel that was located at a depth of 16-18 m, an area was about 1 thousand m2, was used for long-term observations of the permafrost state. The gophers were transported to the tunnel in November and remained in it until early April. For five months, hibernation took place at a temperature close to natural, in absolute darkness and the absence of any noise effects, common during hibernation in vivarium.
Since statistical data from four animals was not enough, previously obtained materials on body temperature in hibernation of three individuals of ground squirrels, hibernation of which previously took place in the basement of the vivarium of IBPK SB RAS at ambient temperatures from 3-5 to minus 10°С (Anufriev , 2013).
To calculate the dependence of the duration of hibernation on the environmental temperature, data on the course of hibernation of eight gophers on the temperature change in the litter of their wintering nests were used and recalculated (Ануфриев, 2008). In all cases, the environmental temperature at which hibernation took place was measured separately. More than 60 thousand hours of observations of hibernation of gophers on body temperature and dynamics of temperature in the litter of nests of wintering animals were analyzed.
Results
In the natural environment, hibernation of gophers begins in late September-early October and ends in the first decade of April (Larionov, 1943, 1958). In the experiment, the first hibernating animals were observed in the third decade of September. However, most of the animals began the hibernation from mid-October to early November. Hibernation ended during the first – second decade of April. The timing of hibernation in the vivarium and the natural environment were close. The initial period of hibernation was characterized by several short dives into hypothermia, then a certain rhythm of hibernation was established in accordance with the ambient temperature. In the spring, before the end of hibernation, the frequency of awakenings increased (Fig. 1, table. 1).
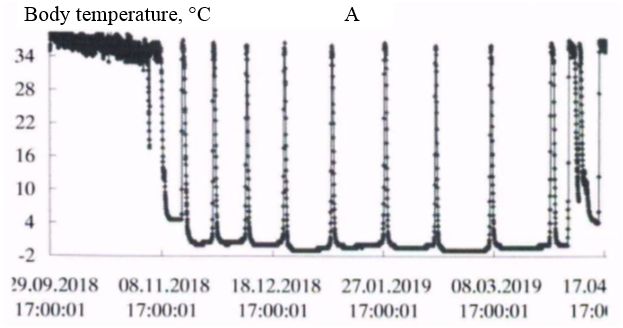
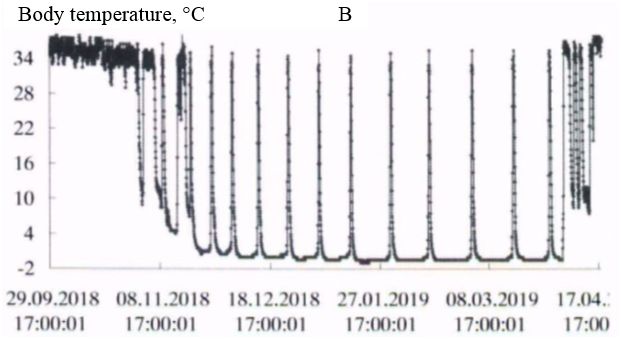
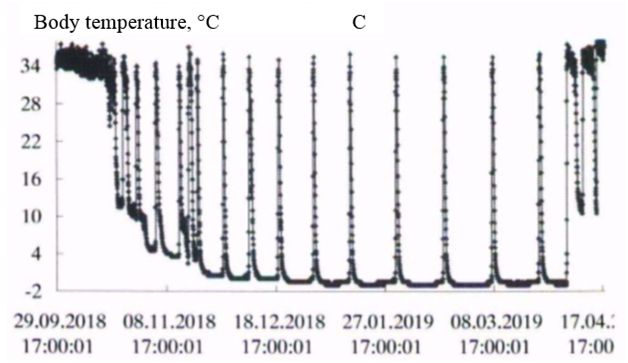
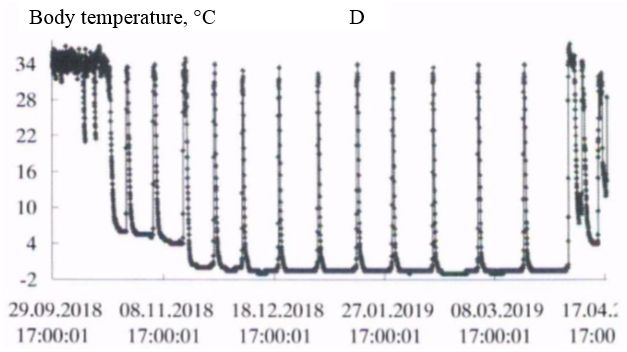
Fig. 1. The Dynamics of body temperature in long-tailed gophers in hybernation period: A - male 662; B – female 455; C - female 485; D – female 446
As the duration of hypothermia periods increased, the body temperature dropped more strongly with each immersion in hypothermia, in accordance with the ambient temperature. The ambient temperature in October was 222 (4.9 ± 0.01) / 4.5 + 6.0; in November – 720 (0.37 ± 0.14) / -5.0 + 5.5; in December – 744 (-5.4 ± 0.01) / -6.0 + -3.0; in January – 744 (-6.3 ± 0.01) / -7.0 + -4.5; in February – 672 (-6.5 ± 0.00) / -6.5 + -5.5; in March – 744 (-6.0 ± 0.01) / -6.5 + -4.5; in April – 349 (1.2 ± 0.22) / -6.5 + 5.5 °C.
Table 1. Body temperature of long-tailed gophers during winter hibernation
| Period / individual | ♂662* | ♀455 | ♀485 | |
| September | n(M±m) Min-Max | 31 (37.7 ± 0.16) 33.0–39.0 | 31(35.6 ± 0.20) 33.0–37.5 | 31(34.8 ± 0.12) 33.5–36.0 |
| October | n(M±m) Min-Max | 744 (36.1 ± 0.04) 33.0–36.0 | 744(34.9 ± 0.05) 38.5–38.0 | 744(27.9 ± 0.36) 8.0–37.5 |
| November | n(M±m) Min-Max | 744 (13.7 ± 0.6) 0.0–37.0 | 720(14.5 ± 0.5) 0.5–38.5 | 720(8.1 ± 0.36) 0.5–37.0 |
| December | n(M±m) Min-Max | 744 (1.99 ± 0.26) -1.0–36.5 | 744(2.8 ± 0.28) -0.5–35.5 | 744(2.1 ± 0.26) -0.5–35.5 |
| January | n(M±m) Min-Max | 744 (2.17 ± 0.29) -1.0–36.5 | 744(1.9 ± 0.27) -1.0–35.5 | 744(1.37 ± 0.26) -1.0–36.0 |
| February | n(M±m) Min-Max | 672 (0.4 ± 0.20) -1.0–36.0 | 672(1.4 ± 0.24) -0.5–35.5 | 672(0.87 ± 0.24) -1.0–35.5 |
| March | n(M±m) Min-Max | 744 (2.45 ± 0.32) -1.0–36.5 | 744(1.8 ± 0.27) -0.5–35.5 | 744(1.42 ± 0.27) -1.0–36.0 |
| April | n(M±m) Min-Max | 424 (16.6 ± 0.72) 0.0-37.0 | 424(21.1 ± 0.72) -0.5–38.5 | 424(24.02 ± 0.72) -1.0–39.0 |
Note. n – number of measurements; * - body weight after hibernation
In all experimental animals in the period of "deep" hibernation (December – March), the body temperature during periods of hypothermia decreased to values below 0°; at awakenings, during periods of normothermia, it reached values of 35.0 ÷ 36.5°C. In ground squirrels before and after hibernation the body temperature changed in the range 37-39°C, characteristic for homoiothermy.
Long-tailed gophers in a state of hypothermia during hibernation are highly resistant to cold. Thus, when wintering in the permafrost tunnel of IMZ SB RAS animals were in a state of hypothermia with a body temperature from 0 to minus 1°C from 58 to 68 % of their hibernation time (table. 2). The average duration of bauts was 225-331 hours, awakenings – from 15 to 23 hours. The maximum duration of the hypothermic period in hibernation in an adult male – 501 hours (about 21 day; fig. 1, table. 3).
The total duration of hibernation ranged from 3.5 to 4 thousand hours.
Table 2. Duration of hypothermia in long-tailed gophers in hibernation with body temperature 0 °C and below (h), 2018/19
| ♂662 | ♀455 | ♀485 | |
| November | - | - | - |
| December | 547 | 253 | 547 |
| January | 632 | 590 | 632 |
| February | 584 | 591 | 584 |
| March | 574 | 580 | 574 |
| April | 84 | 70 | 84 |
| Total, h/%* | 2331.0/66.1 | 2084.0/58.2 | 2421.0/61.3 |
Note. % * from the total hibernation time
Table 3. Characteristics of hibernation rhythms of long-tailed gophers, 2018/19
| Animal/mass, g, after hibernation | ♂662 | ♀455 | ♀485 |
| Mean duration of hypothermia, h,
*n(M±m) Min-Max |
10(331.1 ± 40.8) 122.0–501.0 | 15(225.0 ± 25.3) 70.0–362.0 | 16(231.9 ± 31.7) 48.0–41123.0 |
| Total duration of hypothermia, h/% | 3313.0/94.1 | 3375.0/94.04 | 3711.0/94.07 |
| Mean duration of awakenings, h,
n(M±m) Min-Max |
9(23.4 ± 1.8) 19.0–36.0 | 14(15.3 ± 0.5) 12.0–18.0 | 15(15.6 ± 0.6) 12.0–19.0 |
| Total duration of normothermia, h/% | 211.0/5.9 | 214.0/5.96 | 234.0/5.93 |
| Total duration of hibernation, h/days | 3524/146.8 | 3589.0/149.5 | 3945.0/164.4 |
Note. *n – number of periods of hibernation or awakening
Previously, such a duration of bauts in long-tailed gopher was not noted, the maximum duration was 17.8 days. (Anufriev, 2008). In addition, the animal, in which such a long period of hypothermia was registered, to the beginning of the process was a one-year-old male weighing 920 g, during 3.5 thousand hours of hibernation loss up to 258 g of body weight (about 1.7 g/day).
Long-tailed gophers are excellent hibernators, during hibernation, the proportion of time that animals spend in a hypothermia state is 94-95 % of the total budget of hibernation time. In all individuals, wintering in an underground laboratory at 16 m depth in permafrost conditions, the maximum duration of bauts was from 15 to 20 days., the entire hibernation included from 10 to 16 episodes of hypothermia. The period of "deep" hibernation, with a duration above 10 days., consisted of 8-10 bauts. The dependence of the body temperature on the environmental temperature, according to the form of a graphical curve, is approximated close to exponential and linear dependencies. The body temperature below zero in long-tailed gophers is noted at ambient temperatures minus 5-6°C and below (Fig. 2).
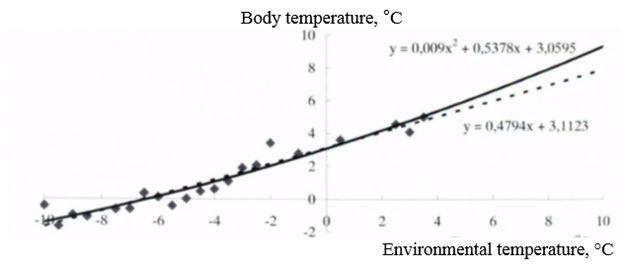
Fig. 2. Dependence of body temperature of long-tailed gopher during winter hibernation on ambient temperature
Since most of the hibernation of the long-tailed gopher in the natural environment takes place at near- and below-zero temperatures, during the experiments the environmental temperature for most part of the hibernation was below 0°C. The long-tailed gopher has a dependence of the duration of hypothermia periods on the ambient temperature. The longest periods of hypothermia were observed in the temperature range from 3-4°C to minus 4-5°C. The duration of hypothermia periods in this interval was about 8-15 days (Fig. 3).
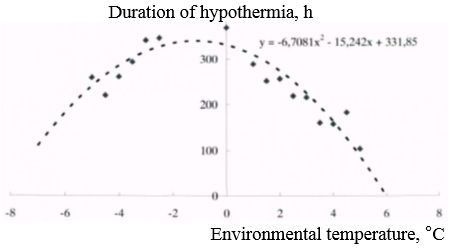
Fig. 3. Dependence of duration of hypothermia periods in long-tailed gopher on ambient temperature
The body temperature of the long-tailed gopher in hibernation depends on the ambient temperature, and the duration of periods of hibernation - from the ambient temperature, so it is natural that the duration of hibernation is different at different body temperatures (Fig. 4). The dependence is approximated by a logarithmic graphical curve. The longest periods of hypothermia were observed at near-zero body temperatures and up to minus 1°C. The duration of periods of hibernation in this case was about and more than 300 hours.
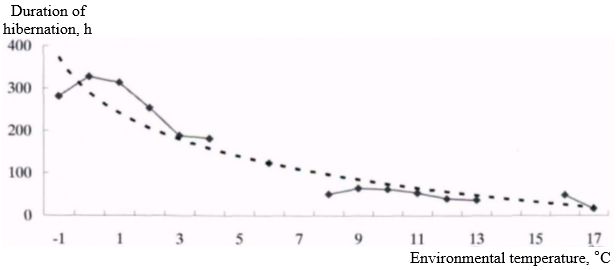
Fig. 4. Dependence of duration of hibernation periods on body temperature in long-tailed gopher in hibernation
Discussion
With the transition of winter-sleeping animals to the life in burrows and the loss of communication with the soil surface, the number of external factors affecting them, synchronizing endogenous rhythms with exogenous ones, significantly decreases. The periodizm of illumination, temperature and humidity disappears. Only temperature and, to a lesser extent, atmospheric pressure and soil moisture remain the main factors allowing to synchronize endogenous rhythms with seasonal changes in the environment (Anufriev, 2008). The long-tailed ground squirrel is among the species, along with the arctic ground squirrel (Spermophilus parryii), the marmot (Marmota camtschatica) and the hedgehogs (Erinaceus roumanicus and E. europaeus), in which the body temperature during hibernation can be 0°C and below (Anufriev , 2008; Anufriev, Ohlopkov, 2015; Rutovskaya et al., 2019b).
Inhabiting the most Northern part of the species' range, the long-tailed ground squirrel perfectly adapted to wintering conditions in negative ambient temperatures. Previously, the zone of optimal for hibernation temperatures of the environment was noted for this species (Anufriev , 2008). In this temperature range, the daily loss of body weight was minimal, the metabolic rate of oxygen consumption was also significantly lower than outside this temperature range. The optimal body temperature for hibernation in the long-tailed gopher has near-zero values, as in the white-breasted hedgehog (Rutovskaya et al., 2019a). The copperhead hibernation periods are longest at body temperatures of about 2°C (Twente, Twente, 1965, 1967), and hypothermia is noted to a body temperature of 22°C, in the white-breasted hedgehog to 22°C, in the long-tailed gopher to 17°C. In all three species, the graphical dependence was approximated by a logarithmic curve. A decrease in body temperature in long-tailed gophers to minus 1°C, as well as an increase to 2-4°C, caused a decrease in the duration of hypothermia periods, respectively, an increase in energy consumption in hibernation. The dependence of the body temperature of the long-tailed gopher on the ambient temperature is clearly expressed. Its graph is rather closer to the exponent, although in some places it is almost linear. Before us similarities and differences in hibernation of the long-tailed gopher with the Arctic gopher and black-capped marmot in Yakutia are considered (Anufriev, Ohlopkov, 2015). Most of the processes occurring in hibernation in the compared species were similar.
More recently, reports of hibernation of the white-breasted (E. roumanicus) and European (E. europaeus) hedgehog, including those with body temperatures below 0°C (Rutovskaya et al., 2019a, b), have been published. In the white-breasted hedgehog in a hypothermia state the presence of an optimal for hibernation temperature zone, which includes a temperature interval with positive and negative ambient temperatures was also revealed. The optimal for hibernation body temperature was also in the range of near-zero temperatures, and the minimum body temperature in hypothermia was minus 1.3°C, in the long-tailed gopher minus 2°C (Anufriev , 2008). In these phylogenetically quite remote species, white-breasted hedgehog and long-tailed gopher, there are a number of similarities in the overall organization of hibernation and parts of its components. In the initial period of hibernation, the intervals of hypothermia are relatively short, from November to March they reach the maximum duration, in hedgehogs up to 17, in gophers up to 18-20 days. Before the end of hibernation, in late March – early April, awakenings become more frequent. These similar in body mass species are characterized by a relatively rapid increase in temperature during spontaneous awakenings. In about five hours, animals with near-zero positive or negative body temperatures turn into homoiotherms with a body temperature about 35°C. The mechanisms of influence of environmental temperature on the formation of hibernation rhythms are similar. Both species are capable of hibernation under negative ambient temperatures, with body temperatures below 0°C (Rutovskaya et al., 2019a, b).
There are specific features of hibernation and hibernation of the compared species. White-breasted hedgehogs lie in hibernation much earlier than the long-tailed ground squirrels. From the first decade of September, short-term, gradually increasing periods of hypothermia were noted in hedgehogs, respectively, the hibernation season in these animals is longer. In the total budget of hibernation time, the proportion of homoiothermy in hedgehogs is higher, from 7 to 20 %, than in gophers. The average duration of the hypothermia periods in white-breasted hedgehogs were significantly lower than the long-tailed ground squirrels. The decrease in body temperature after spontaneous awakening is 30-40 % longer than in long-tailed ground squirrels.
Conclusions
The long-tailed ground squirrel in the North of its area demonstrates a high level of adaptation to environmental conditions during hibernation. The environmental temperature directly affects and is a regulator in the formation of hibernation rhythms. The temperature optimum for hibernation includes positive and negative temperatures corresponding to the winter course of temperatures of the location of wintering nests in the soil, the main habitats of animals. The body temperature of long-tailed gophers in hibernation depends on the environmental temperature, and the optimal body temperature for hibernation is in the range of near-zero values. A number of hibernation mechanisms identified for long-tailed ground squirrels are not unique to the family Sciuridae species, but is noted in representatives of the fam. Erinaceus.
References
Anufriev A. I. Ohlopkov I. M. Hibernation of three species of SCIURIDAE with body temperature below zero, Ekologiya. 2015. No. 1. P. 1–9.
Anufriev A. I. Ecological mechanisms of temperature adaptations of mammals and wintering birds of Yakutia. Novosibirsk: Izd-vo SO RAN, 2013. 220 p.
Anufriev A. I. Mechanisms of hibernation of small mammals of Yakutia. Novosibirsk: Izd-vo SO RAN, 2008. 157 p.
Buck P. L., Barnes B. M. Effects of ambient temperature on metabolic rate, respiratory quotient, and torpor in an arctic hibernator, Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000. Vol. 279. No. 1. P. 255–262.
Drescher J. W. Environmental influences on initiation and maintenance of hibernation in the Arctic ground squirrel Citellus undulatus, Ecology. 1967. Vol. 48. No. 6. R. 962–966.
Duboeus R. Physiological compared (Etude sur le mechanism de la thermogenèse of dusommeil chez mammifères). Paris, 1898. 286 p.
French A. R. Allometries of the duration of torpid and euthermic intervals during mammalian hibernation: a test of the theory of metabolic control of the timing of changes in body temperature, J. Comp. Physiol. (B). 1985. Vol. 156. P. 13–19.
Galster W. A., Morrison P. Cyclic changes in carbohydrates concentrates during hibernation in the Arctic ground squirrel, Amer. J. Physiol. 1970. Vol. 218. No. 4. P. 1228–1232.
Heller H., Poulson T. L. Circanian rhythms. II. Endogenous and exogenous factors controlling reproduction and hibernation in chipmunks (Eutamias) and ground squirrel (Spermophilus), Biochem. and Physiol. 1970. Vol. 33. P. 357–383.
Hut R. A., Barnes B. M., Daan S. Body temperature patterns before, during, and after semi-natural hibernation in the European ground squirrel, J. Comp. Physiol. (B). 2002. Vol. 172 (1). P. 47–58.
Kalabuhov N. I. Hibernation of mammals. M.: Nauka, 1985. 264 p.
Kalabuhov N. I. Summer hibernation of ground squirrels (C. fulvus and C. pygmaeus), Trudy laboratorii eksperimental'noy biologii Moskovskogo zooparka. 1929. T. 5. P. 163–176.
Kayser Ch. The physiology of natural hibernation. NY: Pergamon Press, 1961. 325 p.
Kristofersson R., Soivio A. Hibernation of the hedgehog (Erinaceus europaeus L.) The periodicity of hibernation of undisturbed animals during the winter in a constant ambient temperature, Ann. Sci. Fenn. 1964 (A). IV. Biol. Vol. 122. P. 1–22.
Larionov P. D. Ecological observations of the Yakut long-tail gopher, Zoologicheskiy zhurnal. 1943. T. 22. Vyp. 2. P. 234–246.
Larionov P. D. The range of habitat and the number of Yakut long-tailed ground squirrel, Uchenye zapiski Yakutskogo gosudarstvennogo universiteta. 1958. Vyp. 3. P. 35–49.
Oklejewicz M., Daan S. Strijkstra A. M. Temporal organization of hibernation in wild-type and tau mutant Syrian hamsters, J. Comp. Physiol. (B). 2001. Vol. 171. P. 431–439.
Ortmann S., Heldmaier G. Regulation of body temperature and energy requirements of hibernating Alpine marmots (Marmota marmota), J. Physiol. Regulatory Integrative Comp. Physiol. 2000. Vol. 278. No 3. P. 698–704.
Pengelley E. T., Fisher K. C. Onset and cessation of hibernation under constant temperature and light in the golden-mantled ground squirrel, Citellus lateralis, Nature. 1957. Vol. 180. P. 1371–1372.
Pengelley E. T., Fisher K. C. The effect of temperature and photoperiod on the yearly hibernating behavior of captive golden-mantled ground squirrel (Citellus lateralis), Canadian. J. Zool. 1963. Vol. 41. P. 1103–1120.
Pengelley E. T., Fisher K. C. The effect of temperature and rhythmical arousal ground squirrels (Citellus lateralis), Can. J. Zool. 1961. Vol. 39. No 1. P. 105–120.
Rutovskaya M. V. Diatroptov M. E. Kuznecova E. V. Anufriev A. I. Feoktistova N. Yu. Surov A. V. Body temperature dynamics of the white-breasted hedgehog (Erinaceus roumanicus Barrett-Hamilton, 1900) during hibernation, Zoologicheskiy zhurnal. 2019a. No. 5. P. 556–566.
Rutovskaya M. V. Diatroptov M. E. Kuznecova E. V. Anufriev A. I. Feoktistova N. Yu. Surov A. V. The phenomenon of lowering body temperature to negative values in the genus Erinaceus hedgehogs during hibernation, Zhurnal evolyucionnoy biohimii i fiziologii. 2019b. No. 4. (v pech.).
Slonim A. D. Ecological physiology of animals. M.: Vyssh. shk., 1971. 448 p.
Strumwasser F. Factors in the pattern, timing and predictability of hibernation in the squirrel, Citellus beecheryi, Am. J. Physiol. 1959. Vol. 196. P. 8–14.
Twente J. W., Twente J. A. Regulation of hibernation periods by temperature, Proc. Natl. Acad. Sci. USA. 1965. Vol. 54. P. 1058–1061.
Twente J. W., Twente J. A. Seasonal variation in the hibernating behavior of Citellus lateralis, Mammalian hibernating 111. Edinburg-London: Oliver and Boyd, 1967. P. 47–63.
Willis L. S., Goldman S. S., Foster R. F. Tissue K-concentration in relation to the role of kidney in hibernation and in the cause of periodic arousal, Comp. Biochem. and Physiol. 1971. Vol. (A) 39. No. 3. P. 437–445.

 © 2011 - 2026
© 2011 - 2026