



Введение
Зимняя спячка относительно небольших по размерам и массе тела млекопитающих включает два основных компонента: продолжительные оцепенения со значительным снижением метаболизма и температуры тела и относительно короткие периоды гомойотермного состояния. При периодических пробуждениях животные могут либо принимать пищу (хомяки и бурундуки), либо вообще обходиться без нее (суслики, сурки, ежи), выводить из организма только жидкие экскременты или и жидкие, и твердые. Существует ряд гипотез, с определенной долей вероятности объясняющих прерывистость спячки, тем не менее природа, роль и соотношение эндогенных и экзогенных факторов в формировании ритмов зимней спячки окончательно не определены (Ануфриев, 2008).
Интервал внешних температур, в границах которого возможно протекание спячки, у зимоспящих млекопитающих ограничен. Для большинства гибернантов он находится в пределах от -5 до 15 оС (Слоним, 1971). Для сусликов верхняя граница температуры среды, при которой возможна спячка, расположена около 22 оС, при более высоких температурах состояние спячки не наступает. У сурков этот показатель также находится около 22 оС, у хомячков в пределах 9‒12 оС, у летучих мышей выше 22 оС (Калабухов, 1929; Kayser, 1961; Pengelley, Fisher, 1957, 1963).
Длительность периодов гипотермии ‒ баутов (от англ. bout ‒ период) имеет внутривидовые, межвидовые и сезонные особенности (Strumwasser, 1959; Pengelley, Fisher, 1961; Ortmann, Heldmaier, 2000; Oklejewicz et al., 2001; Hut et al., 2002; Калабухов, 1985). Например, у европейского ежа (Erinaceus europaeus) в начале спячки продолжительность баутов составляет 2‒3 суток, в середине спячки – две-три недели. У сусликов максимальная продолжительность периодов оцепенения в основном не более двух недель, у сурков – две-три недели, у более мелких зимоспящих грызунов (хомячки, бурундуки) ‒ в пределах 7 суток (Duboeus, 1898; Kayser, 1961; Galster, Morrison, 1970; Willis et al., 1971; French, 1985; Buck, Barnes, 2000; Oklejewicz et al., 2001). Из внешних факторов в возникновении спячки у арктического суслика (Spermophilus parryii) температура среды играет ведущую роль (Dresher, 1967). Выявлена зависимость между глубиной спячки и температурой окружающей среды у золотистых сусликов, летучих мышей, у четырех видов бурундуков рода Eutamias, у европейских ежей, европейских сусликов и альпийских сурков (Kristoffersson, Soivio, 1964; Heller, Poulson, 1970; Ortmann, Heldmaier, 2000; Hut et al., 2002). Зависимость продолжительности периодов гипотермии от температуры тела в спячке выявлена у золотистого суслика (S. lateralis) в интервале температур от 2 до 22 оС. С понижением температуры тела увеличивалась продолжительность периодов оцепенений, зависимость была логарифмической (Twente, Twente, 1965, 1967).
Успешность зимовки гибернантов и выживаемость в спячке зависят от глубины и продолжительности состояния с пониженной температурой тела на протяжении зимнего периода. Вместе с тем механизм влияния температуры на формирование ритмов спячки до конца не понятен, поскольку недостаточно материалов о видовых особенностях этого явления.
Целью настоящей работы было изучение хода зимней спячки длиннохвостого суслика в условиях, близких к естественным, выявление механизмов влияния температуры среды на формирование ритмов зимней спячки.
Материалы
Длиннохвостых сусликов отлавливали в августе в 80 км южнее г. Якутска заливкой нор. В сентябре внутрибрюшинно имплантировали приборы длительной регистрации температуры тела (температурные накопители, термографы DS-1922L; подробное описание прибора и основные характеристики можно найти на сайтах: http://www.elin.ru, http://www.thermochron.ru/, http://www.ibdl.ru/), запрограммированные на измерение температуры с частотой 1 раз в 60 мин. Зимовка и спячка четырех особей длиннохвостых сусликов (Spermophilus undulatus Pallas, 1778) с ноября по апрель проходила в подземелье мерзлотной лаборатории Института мерзлотоведения СО РАН (г. Якутск). Штольня расположена на глубине 16‒18 м, площадь около 1 тыс. м2, используется для многолетних наблюдений за состоянием вечномерзлого грунта. Суслики были перевезены в штольню в ноябре и находились в ней до начала апреля. В течение пяти месяцев спячка проходила при температурном режиме, близком к естественному в природе, в абсолютной темноте и отсутствии каких-либо шумовых воздействий, обычных при зимовке в условиях вивария.
Поскольку статистических данных от четырех животных было недостаточно, использованы и пересчитаны ранее полученные материалы по температуре тела в спячке трех особей длиннохвостых сусликов, спячка которых ранее проходила в подвале вивария ИБПК СО РАН при температурах среды от 3‒5 до минус 10 оС (Ануфриев, 2013).
Для расчетов зависимости продолжительности спячки от температуры среды использованы и пересчитаны данные о ходе спячки восьми длиннохвостых сусликов по изменению температуры в подстилке их зимовочных гнезд (Ануфриев, 2008). Во всех вариантах отдельно измерялась температура среды, при которой проходила спячка. Суммарно проанализировано свыше 60 тыс. часов наблюдений за ходом зимней спячки длиннохвостых сусликов по температуре тела и динамике температуры в подстилке гнезд зимующих животных.
Методы
Рассчитывалась ежемесячная усредненная температура тела, бюджет времени на части, составляющие спячку (гипотермия и пробуждения), время нахождения зверьков в гипотермии с температурой тела ниже 0 оС. Определялась зависимость температуры тела зверьков в гипотермии от температуры среды, зависимость продолжительности гипотермии от температуры тела и зависимость продолжительности периодов гипотермии от температуры среды.
Зависимость температуры тела от температуры среды рассчитывали так: на цифровой записи температуры тела животных на протяжении спячки усреднялась температура тела за каждый интервал гипотермии. Усреднение проводили после окончания остывания животного и до начала следующего пробуждения. За тот же период времени усреднялась и температура среды. Затем составлялись таблицы, в которые заносились данные по температуре тела в соответствии с усредненной температурой среды. По каждому шагу температуры получалось от нескольких до 10–15 измерений. В сообщение включены суммированные данные по семи животным.
Рассчитывали зависимость продолжительности периодов спячки от температуры тела животных. У каждого зверька по цифровым данным температуры тела рассчитывали усредненную температуру тела, на протяжении каждого интервала спячки, и длительность этого состояния в часах. Затем готовили электронную таблицу, отражавшую зависимость продолжительности интервалов гипотермии от температуры тела, с шагом 0.5 оС, в которую заносили полученные данные. Продолжительности гипотермии при одной температуре суммировались, и рассчитывалось среднее, затем строились графики. Использованы данные также от семи животных.
Зависимость продолжительности спячки от температуры среды рассчитывалась следующим образом. В данных по температуре тела, или температуре в подстилке гнезда, определялась продолжительность гипотермии за каждый интервал гипотермии, а в материалах по температуре окружающей среды рассчитывалась средняя температура за эти периоды. Таблица заполнялась также с шагом 0.5 оС, и после расчета усредненных величин по каждой температуре строили графики. Зависимость длительности баутов от температуры среды рассчитана по цифровым данным от 15 особей.
Начало периода гипотермии при залеганию в спячку считали с момента, когда температура тела понижалась до 25 оС. Начало периода нормотермии (разогрев) считали с устойчивого повышения температуры тела со скоростью не менее 0.05 оС за 20 мин. В описательной статистике использовали средние ± ошибку средней.
Работу с животными проводили с учетом рекомендаций национального стандарта, по принципам надлежащей лабораторной практики Российской Федерации ГОСТ З 53434-2009.
Результаты
В условиях естественной среды зимняя спячка длиннохвостых сусликов начинается в конце сентября – начале октября и заканчивается в первой декаде апреля (Ларионов, 1943, 1958). В эксперименте первые залегшие в спячку зверьки наблюдались в третьей декаде сентября. Вместе с тем основная масса животных залегала в спячку с середины октября до начала ноября. Заканчивалась спячка на протяжении первой – второй декады апреля. Сроки зимней спячки в виварии и естественной среде близки. Начальный период спячки характеризовался несколькими непродолжительными погружениями в гипотермию, затем в соответствии с температурой окружающей среды устанавливалась определенная ритмика спячки. Весной, перед завершением спячки частота пробуждений возрастала (рис. 1, табл. 1).
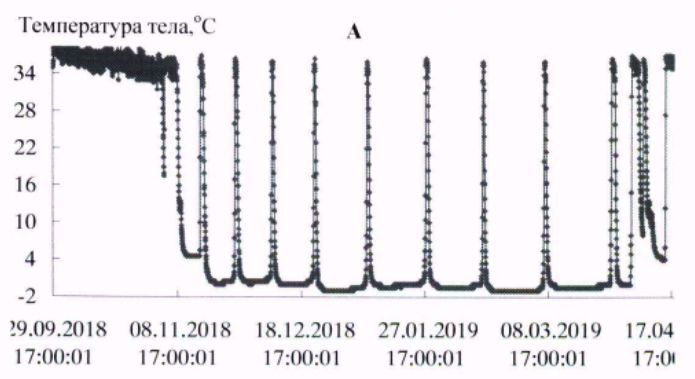
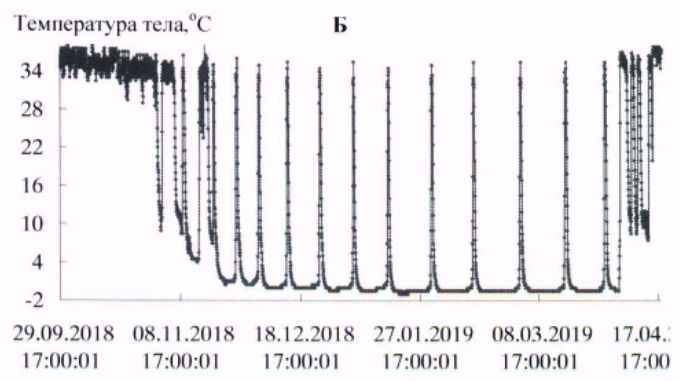
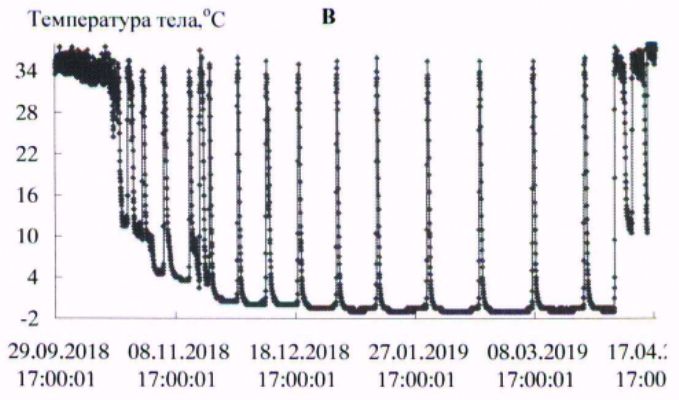
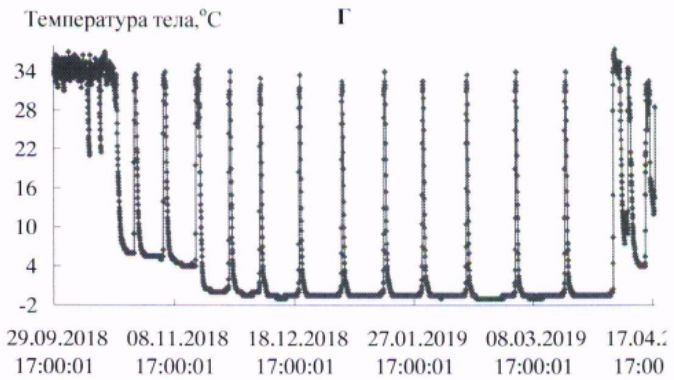
Рис. 1. Динамика температуры тела у длиннохвостых сусликов в период зимней спячки: А- самец 662, Б - самка 455, В - самка 485, Г - самка 446
Fig. 1. The Dynamics of body temperature in long-tailed gophers in hybernation period: A - male 662; B – female 455; C - female 485; D – female 446
С нарастанием продолжительности периодов гипотермии температура тела с каждым погружением в гипотермию понижалась сильнее, в соответствии с температурой окружающей среды. Температура среды в октябре была 222(4.9 ± 0.01) / 4.5 ÷ 6.0; в ноябре – 720(0.37 ± 0.14) / -5.0 ÷ 5.5; в декабре – 744(-5.4 ± 0.01) / -6.0 ÷-3.0; в январе – 744(-6.3 ± 0.01) / -7.0 ÷ -4.5; в феврале – 672(-6.5 ± 0.00) / -6.5 ÷ -5.5; в марте – 744(-6.0 ± 0.01) / -6.5 ÷ -4.5о; в апреле – 349(1.2 ± 0.22) / -6.5 ÷ 5.5 оС.
Таблица 1. Температура тела длиннохвостых сусликов в период зимней спячки
Table 1. Body temperature of long-tailed gophers during winter hibernation
| Период / особь | ♂662* | ♀455 | ♀485 | ♀446 | |
| Сентябрь | n (M ± m) Min-Max | 31(37.7 ± 0.16) 33.0–39.0 | 31(35.6 ± 0.20) 33.0–37.5 | 31(34.8 ± 0.12) 33.5–36.0 | 31(34.69 ± 0.21) 32.0–36.5 |
| Октябрь | n (M ± m) Min-Max | 744(36.1 ± 0.04) 33.0–36.0 | 744(34.9 ± 0.05) 38.5–38.0 | 744(27.9 ± 0.36) 8.0–37.5 | 744(24.9 ± 0.45) 5.5–37.0 |
| Ноябрь | n (M ± m) Min-Max | 744(13.7 ± 0.6) 0.0–37.0 | 720(14.5 ± 0.5) 0.5–38.5 | 720(8.1 ± 0.36) 0.5–37.0 |
720(6.4 ± 0.33) 0.0–35.0 |
| Декабрь | n (M ± m) Min-Max | 744(1.99 ± 0.26) -1.0–36.5 | 744(2.8 ± 0.28) -0.5–35.5 | 744(2.1 ± 0.26) -0.5–35.5 | 744(1.16 ± 0.21) -1.0–35.5 |
| Январь | n (M ± m) Min-Max | 744(2.17 ± 0.29) -1.0–36.5 | 744(1.9 ± 0.27) -1.0–35.5 | 744(1.37 ± 0.26) -1.0–36.0 | 744(2.08 ± 0.27) -0.5–35.0 |
| Февраль | n (M ± m) Min-Max | 672(0.4 ± 0.20) -1.0–36.0 | 672(1.4 ± 0.24) -0.5–35.5 | 672(0.87 ± 0.24) -1.0–35.5 | 672(0.24 ± 0.17) -1.0–35.5 |
| Март | n (M ± m) Min-Max | 744(2.45 ± 0.32) -1.0–36.5 | 744(1.8 ± 0.27) -0.5–35.5 | 744(1.42 ± 0.27) -1.0–36.0 | 744(1.19 ± 0.22) -1.0–35.5 |
| Апрель | n (M ± m) Min-Max | 424(16.6 ± 0.72) 0.0-37.0 | 424(21.1 ± 0.72) -0.5–38.5 | 424(24.02 ± 0.72) -1.0–39.0 | 426(14.15 ± 0.62) -0.5–37.5 |
Примечание. n – число измерений; * – масса тела после окончания спячки
У всех экспериментальных животных в период «глубокой» спячки (декабрь – март) температура в полости тела в периоды гипотермии понижалась до значений ниже 0о, при пробуждениях, в периоды нормотермии, достигала значений 35.0 ÷ 36.5 оС. У сусликов до и после спячки температура тела изменялась в диапазоне 37–39 оС, характерном для гомойотермии.
Длиннохвостые суслики в состоянии гипотермии в период зимней спячки обладают высокой устойчивостью к воздействию холода. Так, при перезимовке в мерзлотной штольне ИМЗ СО РАН зверьки от 58 до 68 % бюджета времени спячки находились в состоянии гипотермии с температурой тела от 0 до минус 1 оС (табл. 2). Средняя продолжительность баутов составляла 225–331 ч, пробуждений – от 15 до 23 ч. Максимальная продолжительность гипотермного периода в спячке у взрослого самца – 501 ч (около 21 сут.; рис. 1, табл. 3).
Таблица 2. Продолжительность гипотермии у длиннохвостых сусликов в спячке с температурой тела 0 оС и ниже (ч), 2018/19 г.
Table 2. Duration of hypothermia in long-tailed gophers in hibernation with body temperature 0 °C and below (h), 2018/19
| Животное / месяц | ♂662 | ♂455 | ♀485 | ♀446 |
| Ноябрь | – | – | – | 147 |
| Декабрь | 547 | 253 | 547 | 635 |
| Январь | 632 | 590 | 632 | 588 |
| Февраль | 584 | 591 | 584 | 600 |
| Март | 574 | 580 | 574 | 638 |
| Апрель | 84 | 70 | 84 | 82 |
| Общее, ч/%* | 2331.0/66.1 | 2084.0/58.2 | 2421.0/61.3 | 2690/67.7 |
Примечание. %* – от общего времени спячки
Общая продолжительность спячки составляла от 3.5 до 4 тыс. часов.
Таблица 3. Характеристика ритмов зимней спячки длиннохвостых сусликов, 2018/19 г.
Table 3. Characteristics of hibernation rhythms of long-tailed gophers, 2018/19
| Животное / масса, г, после спячки | ♂662 | ♂455 | ♀485 | ♀446 |
| Средняя длит. гипотермии, ч, *n(M ± m) / Min-Max |
10(331.1 ± 40.8) 122.0–501.0 | 15(225.0 ± 25.3) 70.0–362.0 | 16(231.9 ± 31.7) 48.0–412.0 | 13(289.5 ± 20.5) 137.0–381.0 |
| Общая длительность гипотермии, ч/% | 3313.0/94.1 | 3375.0/94.04 | 3711.0/94.07 | 3796/94.74 |
| Средняя длит. пробуждений, ч, n(M ± m) / Min-Max | 9(23.4 ± 1.8) 19.0–36.0 | 14(15.3 ± 0.5) 12.0–18.0 | 15(15.6 ± 0.6) 12.0–19.0 | 12(17.4 ± 0.6) 14.0–21.0 |
| Общая длит. нормотермии, ч/% | 211.0/5.9 | 214.0/5.96 | 234.0/5.93 | 209.0/5.26 |
| Общее время спячки, ч /сут. |
3524/146.8 | 3589.0/149.5 | 3945.0/164.4 | 3972.0/165.5 |
Примечание. *n – число периодов спячки или пробуждений
Ранее подобной продолжительности баутов у длиннохвостого суслика не отмечалось, максимальная продолжительность составляла 17.8 сут. (Ануфриев, 2008). Добавим, что зверек, у которого зарегистрирован такой продолжительный период гипотермии, к началу процесса – годовалый самец массой тела 920 г, за 3.5 тыс. ч спячки потери массы тела составили 258 г (около 1.7 г/сут.).
Длиннохвостые суслики – превосходные гибернаторы, во время зимней спячки доля времени, которое зверьки проводят в состоянии гипотермии, составляет 94–95 % от общего бюджета времени зимней спячки. У всех особей, зимовавших в подземной лаборатории на глубине 16 м в условиях вечномерзлого грунта, максимальная продолжительность баутов была от 15 до 20 сут., вся спячка включала от 10 до 16 эпизодов гипотермии. Период «глубокой» спячки, с продолжительностью выше 10 сут., состоял из 8–10 баутов. Зависимость температуры тела от температуры среды, по форме графической кривой, аппроксимируются близкими экспоненциальной и линейной зависимостями. Температура тела ниже нуля у длиннохвостых сусликов отмечена при температурах среды минус 5–6 оС и ниже (рис. 2).
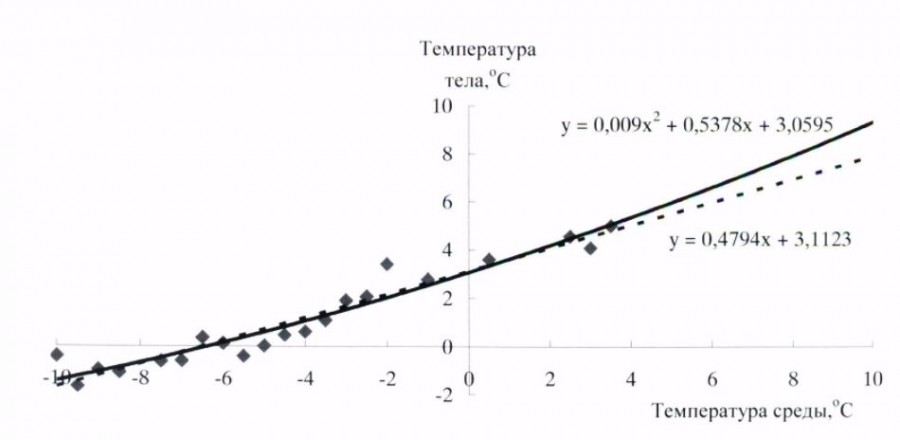
Рис. 2 Зависимость температуры тела от температуры среды у длиннохвостого суслика в период зимней спячки
Fig. 2. Dependence of body temperature of long-tailed gopher during winter hibernation on ambient temperature
Поскольку большая часть зимней спячки у длиннохвостого суслика в естественной среде проходит при околонулевых и температурах ниже 0 оС, при проведении экспериментов температура среды на протяжении большей части спячки была ниже 0 оС. У длиннохвостого суслика имеется зависимость продолжительности периодов гипотермии от температуры окружающей среды. Наиболее продолжительные периоды гипотермии отмечены в температурном диапазоне от 3–4 оС до минус 4–5 оС. Продолжительность периодов гипотермии в этом интервале составляла около 8–15 сут. (рис. 3).
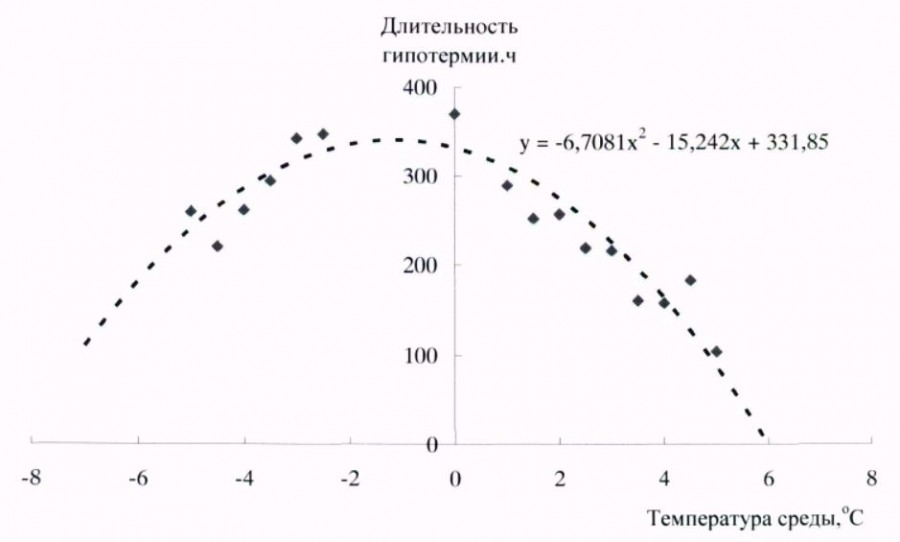
Рис. 3. Зависимость продолжительности периодов гипотермии от температуры среды у длиннохвостого суслика в период спячки
Fig. 3. Dependence of duration of hypothermia periods in long-tailed gopher on ambient temperature
Температура тела у длиннохвостого суслика в спячке зависит от температуры окружающей среды, а продолжительность периодов спячки – от температуры среды, поэтому вполне закономерно, что и продолжительность спячки различна при различных температурах тела (рис. 4). Зависимость аппроксимируется логарифмической графической кривой. Наиболее продолжительные периоды гипотермии отмечены при околонулевых температурах тела и до минус 1 оС. Продолжительность периодов спячки в этом случае составляла около и более 300 ч.
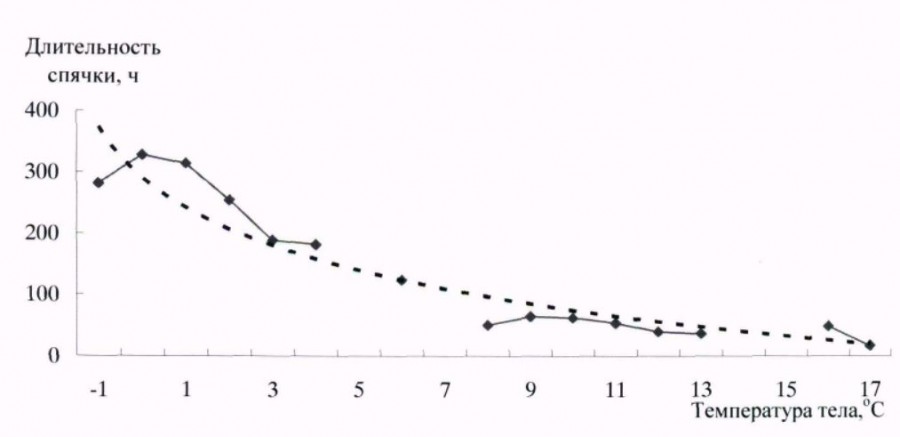
Рис. 4. Зависимость продолжительности периодов спячки от температуры среды у длиннохвостого суслика в спячке
Fig. 4. Dependence of duration of hibernation periods on body temperature in long-tailed gopher in hibernation
Обсуждение
С переходом зимоспящих животных к норной жизни и потерей связи с поверхностью почвы количество воздействующих на них внешних факторов, синхронизирующих эндогенные ритмы с экзогенными, значительно снижается. Исчезает периодизм освещенности, температуры и влажности воздуха. Основными факторами, позволяющими синхронизировать эндогенные ритмы с сезонными изменениями внешней среды, остаются лишь температура и, в меньшей степени, атмосферное давление и влажность почвы (Ануфриев, 2008). Длиннохвостый суслик входит в число видов, наряду с арктическим сусликом (Spermophilus parryii), черношапочным сурком (Marmota camtschatica), белогрудым (Erinaceus roumanicus) и европейским (E. europaeus) ежами, у которых в период зимней спячки температура тела может иметь значение 0 оС и ниже (Ануфриев, 2008; Ануфриев, Охлопков, 2015; Рутовская и др., 2019б).
Заселяя наиболее северный участок ареала вида, длиннохвостый суслик прекрасно адаптировался к условиям зимовки в условиях отрицательных окружающих температур. Ранее для этого вида уже отмечалась зона оптимальных для зимней спячки температур среды (Ануфриев, 2008). В этом температурном интервале суточные потери массы тела были минимальными, уровень метаболизма по потреблению кислорода также был значительно ниже, чем вне данного интервала температур. Оптимальная для спячки температура тела у длиннохвостого суслика имеет околонулевые значения, как и у белогрудого ежа (Рутовская и др., 2019а). У золотистого суслика периоды спячки наиболее продолжительны при температурах тела около 2 оС (Twente, Twente, 1965, 1967), а гипотермия отмечена до температуры тела 22 оС, у белогрудого ежа до 22 оС, у длиннохвостого суслика до 17 оС. У всех трех видов графическая зависимость аппроксимировалась логарифмической кривой. Снижение температуры тела у длиннохвостых сусликов до минус 1 оС, как и увеличение до 2–4 оС, вызывало уменьшение продолжительности периодов гипотермии, соответственно рост энергозатрат в спячке. Зависимость температуры тела длиннохвостого суслика от температуры окружающей среды выражена отчетливо. Ее график, скорее, ближе к экспоненте, хотя на отдельных участках почти линеен. Ранее нами рассмотрены сходство и различия зимней спячки длиннохвостого суслика с арктическим сусликом и черношапочным сурком в Якутии (Ануфриев, Охлопков, 2015). Большинство процессов, происходящих в спячке, у сравниваемых видов было сходным.
Совсем недавно опубликованы сообщения о зимней спячке белогрудого (E. roumanicus) и европейского (E. europaeus) ежа, в том числе и с температурами тела ниже 0 оС (Рутовская и др., 2019а, б). У белогрудого ежа в состоянии гипотермии также выявлено наличие оптимальной для зимней спячки температурной зоны, которая включает температурный интервал с положительными и отрицательными температурами среды. Оптимальная для спячки температура тела также находилась в области околонулевых температур, а минимальная температура в полости тела в гипотермии составляла минус 1.3 оС, у длиннохвостого суслика минус 2 оС (Ануфриев, 2008). У этих филогенетически достаточно удаленных видов, белогрудого ежа и длиннохвостого суслика, прослеживается ряд сходных черт в общей организации зимней спячки и частей, ее составляющих. В начальный период спячки интервалы гипотермии относительно непродолжительны, с ноября по март достигают максимальной длительности, у ежей до 17, у сусликов до 18–20 сут. Перед окончанием спячки, в конце марта – начале апреля, пробуждения учащаются. Для этих близких по массе тела видов характерен относительно быстрый рост температуры при спонтанных пробуждениях. Примерно за пять часов животные с околонулевыми положительными либо отрицательными температурами тела превращаются в гомойотермных с температурой в полости тела около 35 оС. Сходны и механизмы влияния температуры среды на формирование ритмов зимней спячки. Оба вида способны к спячке в условиях отрицательных температур среды, с температурой тела ниже 0 оС (Рутовская и др., 2019а, б).
Имеются видовые особенности зимовки и спячки сравниваемых видов. Белогрудые ежи залегают в спячку в сроки, гораздо более ранние, чем длиннохвостые суслики. Уже с первой декады сентября у ежей отмечены кратковременные, постепенно нарастающие периоды гипотермии, соответственно сезон зимней спячки у этих животных более продолжительный. В общем бюджете времени спячки доля гомойотермии у ежей выше, от 7 до 20 %, чем у сусликов. Средняя продолжительность периодов гипотермии у белогрудых ежей была существенно ниже, чем у длиннохвостых сусликов. Снижение температуры тела после спонтанного пробуждения на 30–40 % времени продолжительнее, чем у длиннохвостых сусликов.
Заключение
Длиннохвостый суслик на севере ареала демонстрирует высокий уровень адаптации к условиям среды в период спячки. Температура среды непосредственно влияет и является регулятором при формировании ритмов спячки. Температурный оптимум для спячки включает положительные и отрицательные температуры, соответствующие зимнему ходу температур месторасположения зимовочных гнезд в почве, коренных обитаний животных. Температура тела у длиннохвостых сусликов в спячке зависит от температуры среды, а оптимальная для спячки температура тела находится в области околонулевых значений. Ряд механизмов зимней спячки, выявленных для длиннохвостых сусликов, характерен не только для видов сем. Sciuridae, но отмечен и у представителей сем. Erinaceus.
Библиография
Ануфриев А. И. Механизмы зимней спячки мелких млекопитающих Якутии . Новосибирск: Изд-во СО РАН, 2008. 157 с.
Ануфриев А. И. Экологические механизмы температурных адаптаций млекопитающих и зимующих птиц Якутии . Новосибирск: Изд-во СО РАН, 2013. 220 с.
Ануфриев А. И., Охлопков И. М. Зимняя спячка трех видов SCIURIDAE с температурой тела ниже нуля // Экология. 2015. № 1. С. 1–9.
Калабухов Н. И. Летняя спячка сусликов (C. fulvus и C. pygmaeus) // Труды лаборатории экспериментальной биологии Московского зоопарка. 1929. Т. 5. С. 163–176.
Калабухов Н. И. Спячка млекопитающих . М.: Наука, 1985. 264 с.
Ларионов П. Д. Экологические наблюдения над якутским длиннохвостым сусликом // Зоологический журнал. 1943. T. 22. Вып. 2. С. 234–246.
Ларионов П. Д. Ареал, стации и численность якутского длиннохвостого суслика // Ученые записки Якутского государственного университета. 1958. Вып. 3. С. 35–49.
Рутовская М. В., Диатроптов М. Е., Кузнецова Е. В., Ануфриев А. И., Феоктистова Н. Ю., Суров А. В. Динамика температуры тела белогрудого ежа (Erinaceus roumanicus Barrett-Hamilton, 1900) во время зимней спячки // Зоологический журнал. 2019а. № 5. С. 556–566.
Рутовская М. В., Диатроптов М. Е., Кузнецова Е. В., Ануфриев А. И., Феоктистова Н. Ю., Суров А. В. Феномен снижения температуры тела до отрицательных значений у ежей рода Erinaceus во время зимней спячки // Журнал эволюционной биохимии и физиологии. 2019б. № 4. (в печ.).
Слоним А. Д. Экологическая физиология животных . М.: Высш. шк., 1971. 448 с.
Buck С. L., Barnes B. М. Effects of ambient temperature on metabolic rate, respiratory quotient, and torpor in an arctic hibernator // Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000. Vol. 279. № 1. P. 255–262.
Drescher J. W. Environmental influences on initiation and maintenance of hibernation in the Arctic ground squirrel Citellus undulatus // Ecology. 1967. Vol. 48. № 6. Р. 962–966.
Duboeus R. Physiological compared (Etude sur le mechanism de la thermogenèse of dusommeil chez mammifères). Paris, 1898. 286 p.
French A. R. Allometries of the duration of torpid and euthermic intervals during mammalian hibernation: a test of the theory of metabolic control of the timing of changes in body temperature // J. Comp. Physiol. (B). 1985. Vol. 156. P. 13–19.
Galster W. A., Morrison P. Cyclic changes in carbohydrates concentrates during hibernation in the Arctic ground squirrel // Amer. J. Physiol. 1970. Vol. 218. № 4. P. 1228–1232.
Heller H., Poulson T. L. Circanian rhythms. II. Endogenous and exogenous factors controlling reproduction and hibernation in chipmunks (Eutamias) and ground squirrel (Spermophilus) // Biochem. and Physiol. 1970. Vol. 33. P. 357–383.
Hut R. A., Barnes B. M., Daan S. Body temperature patterns before, during, and after semi-natural hibernation in the European ground squirrel // J. Comp. Physiol. (B). 2002. Vol. 172 (1). P. 47–58.
Kayser Ch. The physiology of natural hibernation. NY: Pergamon Press, 1961. 325 p.
Kristofersson R., Soivio A. Hibernation of the hedgehog (Erinaceus europaeus L.) The periodicity of hibernation of undisturbed animals during the winter in a constant ambient temperature // Ann. Sci. Fenn. 1964 (A). IV. Biol. Vol. 122. P. 1–22.
Oklejewicz M., Daan S. Strijkstra A. M. Temporal organization of hibernation in wild-type and tau mutant Syrian hamsters // J. Comp. Physiol. (B). 2001. Vol. 171. P. 431–439.
Ortmann S., Heldmaier G. Regulation of body temperature and energy requirements of hibernating Alpine marmots (Marmota marmota) // J. Physiol. Regulatory Integrative Comp. Physiol. 2000. Vol. 278. No 3. P. 698–704.
Pengelley E. T., Fisher K. C. Onset and cessation of hibernation under constant temperature and light in the golden-mantled ground squirrel, Citellus lateralis // Nature. 1957. Vol. 180. P. 1371–1372.
Pengelley E. T., Fisher K. C. The effect of temperature and rhythmical arousal ground squirrels (Citellus lateralis) // Can. J. Zool. 1961. Vol. 39. No 1. P. 105–120.
Pengelley E. T., Fisher K. C. The effect of temperature and photoperiod on the yearly hibernating behavior of captive golden-mantled ground squirrel (Citellus lateralis) // Canadian. J. Zool. 1963. Vol. 41. P. 1103–1120.
Strumwasser F. Factors in the pattern, timing and predictability of hibernation in the squirrel, Citellus beecheryi // Am. J. Physiol. 1959. Vol. 196. P. 8–14.
Twente J. W., Twente J. A. Regulation of hibernation periods by temperature // Proc. Natl. Acad. Sci. USA. 1965. Vol. 54. P. 1058–1061.
Twente J. W., Twente J. A. Seasonal variation in the hibernating behavior of Citellus lateralis // Mammalian hibernating 111. Edinburg-London: Oliver and Boyd, 1967. P. 47–63.
Willis L. S., Goldman S. S., Foster R. F. Tissue K-concentration in relation to the role of kidney in hibernation and in the cause of periodic arousal // Comp. Biochem. and Physiol. 1971. Vol. (A) 39. № 3. P. 437–445.
Благодарности
Работа выполнена в рамках темы Государственного задания ИБПК ЯНЦ СОРАН: «Структура и динамика популяций и сообществ животных холодного региона Северо-Востока России в современных условиях глобального изменения климата и антропогенной трансформации северных экосистем: факторы, механизмы, адаптации, сохранение» АААА-А17-117020110058-4.
Авторы статьи очень признательны руководству Института мерзлотоведения СО РАН за возможность проведения исследований в мерзлотной лаборатории Института.

 © 2011 - 2026
© 2011 - 2026