


Introduction
Now it is no doubt that climate is critical for the main characteristics of massive biomes and numerous features of their inhabitants. Therefore, it is necessary to have a clear sense of the power of climatic environmental factors to understand clearly the adaptive reactions to them. Some of these reactions of birds to the climate factors can be found out with the help of ranking species or populations in the axes of spatial variables. (Dunn et al., 2000; Martin et al., 2006; McNamara et al., 2008; Bears et al., 2009; Lu, 2011; Hille, Cooper, 2015; Boyle et al., 2016 etc.) The changes in taxonomical characteristics of body size, fertility, mortality and other characteristics are considered as a reaction to the spatial environmental gradient. In the majority of such studies it is directly or indirectly indicated that definite climatic factors change by studied gradients, but it is also reminded about the adaptation of any species or population to all complex of ambient conditions. In addition to climatic factors this complex includes biotic impacts, including predation risk, competition, population density etc. (e.g. Donázar, 1990; Martin, 1995; Jetz et al., 2008). Conotic variables, such as vegetation type and/or food resources are discussed (Donázar, 1990; McNamara et al., 2008; Boyle et al., 2016). This list can be supplemented with some difficult to account for and because of that non-discussed variables, such as geochemical regime (trophicity) of landscape as well as anthropogenic load able to distort natural bioclimatic trends, including vast areas (Ravkin, Ravkin, 2005). The deformation of explaining variables is also possible when including into the analysis taxons, which areas occupy several continents and/or are represented by circumpolar areas without any internal differentiation (for example, Balasubramaniam, Rotenberry, 2016).
It is almost impossible to isolate the impact of some variables from the complex of factors grouped in this way, and therefore the approach is considered as heuristic (Hille, Cooper, 2005). This approach provides the reliable conclusions only for massive taxons (family and higher), vast territories (tropics/ mid-latitude) or according to clearly differed characteristics, for example, orographic ones (Jetz et al., 2008; Bears et al., 2009; Lu, 2011; Boyle et al., 2016 and others). In fact, this approach makes one operate with the groups of explaining variables, part of them is not quantified, but some variables have not been determined yet. It is this uncertainty of the list of factors and their power that significantly complicate the research: any territory (administrative district, natural zone, species area, its part etc.) is always the complex of environmental impacts, which are difficult to divide, and adaptations produced by them. We tried to maximally concretize and simplify the analysis: we forcedly narrowed the number of explaining factors to the minimum and ranked the space used by birds.
Certainly, climatic indicators change in latitude, longitude and altitude (Alisov, Poltaraus, 1974). However, there were no apparent attempts to adequately translate the three variables into a climatic scale in order to study the adaptive reactions of birds: in available literature we cannot find the investigations of widely spread taxons and species of plant-feeding birds in the axes of only climatic characteristics. It is most probable that this fact reflects the preventive reaction of investigators to the quite expected effects of collinearity. The realization of another approach - ranking populations (below this term is used as a synonym of “regional population”) in the axes of some known variables – make it possible to carry out a visual analysis of a mutual arrangement of populations on factorial gradients and to assess the impact of quite concrete environmental variables on them.
Below the attempt to assess the influence of the main climatic factors on the distribution of wide spread species of birds using this approach is presented. As an example the real populations of wood-grouse were taken: capercaillie Tetrao urogallus and rock capercaillie T. parvirostris. Their arrangement in geographical axes refers to trivial facts. But this arrangement is more likely to be transformed when changing geographical axes on climatic ones. However, it is not easy to assume the character of allocation of the same populations (for example, in the axes of warmth and humidity). For example, where is it warmer: in the northern plains of Eurasia of on the ridge-top parts of its southern mountains? The response is not obvious, as many others. Do po’ulations' centers (points) form compact spot, stretch linearly or on some curves in ecological axes? Do they settle linearly or in clusters? Will this clustering correspond to taxonomic classification? Most of these questions appear due to the multifaceted impact of a terrain on animals. It concerns especially mountain regions where it needs in particular to specify the height limits of vegetation, which can determine the distribution of birds. These limits are not the same in different ranges (Alisov, Poltaraus, 1974). For example, even in adjacent mountain groups (the Alps/ the Dinars) wood-grouse occupies different altitudinal levels (Čas, 2010).
The purpose of the study: 1) to submit the general arrangement of real populations of wood-grouse in the axis of air temperature and precipitation, 2) on the basis of its analysis to assess the mutual arrangement of the populations in these axis, trying to identify the differences and similarities of intergroup climatic adaptations not leaving without attention, whenever possible, the genesis of these differences.
At first sight the achievement of these purposes cannot cause particular differences: climatic results have been collected for the great number of points, but in the case of their absence there are interpolation algorithms. Unfortunately, the spatial location of meteorological stations is quite uneven: they are basically concentrated in the territory of human activity. However, these territories as a rule are not inhabited by wood-grouse, or are only in the periphery of its population areas. In the most of them the meteorological stations are rare. Moreover, in the conditions of contradictory tendencies the interpolations are not reliable. For example, in the mountains where local inversion events can distort the general course of temperature changes (the higher, the colder) by the adverse trend (Khromov, Petrosyants, 2006). In addition, the passage of air masses even over low mountain systems is able to impact totally opposite on the slopes of different expositions: adjacent windward and windless slopes of the same height of even a short mountain ridge usually receive various precipitations, e.g., low-hill terrains of Scotland (Moss, 2015). Within large mountain systems (e.g. the Alps), where the own air masses can be formed (Dobrynin, 1948), it is possible for small mosaic with the strongest variation of precipitations in the local areas of slopes and valleys to be formed (Beniston, 2006). The terrain seems to determine better predictability of temperature regime of the region than precipitation one, since the latter is formed with the greater number of variables.
The strong differences in the terrain between the population arenas of wood-grouse (mountains/valleys) as well as insufficient number of the points with climatic data for most arenas make risky estimates interpolation using some common algorithms. The development of climatic models for every definite population (or their groups) seems more constructive. Let them be even the simplest, but individual models. For the above reasons our work consisted of a successive solution of a number of tasks.
- For the center of every analyzed population its geographical parameters, such as latitude, longitude and altitude were determined.
- For a wide space holding each population (their group) a set of points was chosen, for each point five characteristics: latitude, longitude, absolute altitude, average annual temperature (further, “temperature” ºC) and average annual precipitation (further, “precipitation” mm/year) were estimated. Simultaneous use of the last two indicators largely removes the issues of leveling the seasonal nuances by average annual estimates.
- According to the data of each set of points taken only from the internal contours of population arenas, the individual models (formulae) reflecting the regional dependencies of temperature and precipitation (separately) on latitude, altitude and longitude were calculated.
- Modeling results were verified using climatic sources not engaged to the model design.
- Verified models were used to calculate the values of temperature and precipitation for each population (its center).
- By these calculated values each population was positioned in the axes of temperature and precipitation, the plane between the axes is considered as a thermohumid space.
- The location of points (populations) and/or their groups in this space was characterized by means of a simple visual analysis, more rarely with the help of common correlation and regression procedures.
It should be reminded that both species of wood-grouse are resident phytophages. In the winter their trophic needs are supplied most exclusively by tree vegetation, therefore the northern and southern boundaries of their areas almost coincide with the forests distribution (Kirikov, 1952; Potapov, Sale, 2013). The usage of the space above the upper edge of forests by these dendrophilous birds is poorly studied and, apparently, underestimated (see, e.g. Ostrovsky, 1973; Mènoni, 1991; Neyfeld, Bobretsov, 2002). Nevertheless, in this study all the areas above this line were not included in the analysis as well as the space below the forest boundary: steppe zones of Bashkiria, Altai, Tuva etc.
Materials
Geographical variables
Wood-grouse was or is a traditional object of hunting, and at present in literature one can find estimates of the body weight of these birds belonging to a lot of populations including extinct ones (i.e. indisputable proof of their presence in definite points). Since the populations with such estimates are located throughout the whole area (Fig.1), it is these places that were the objects of the analysis given below. Taxonomic affiliation of the birds is given according to Potapov, Sale (2013).
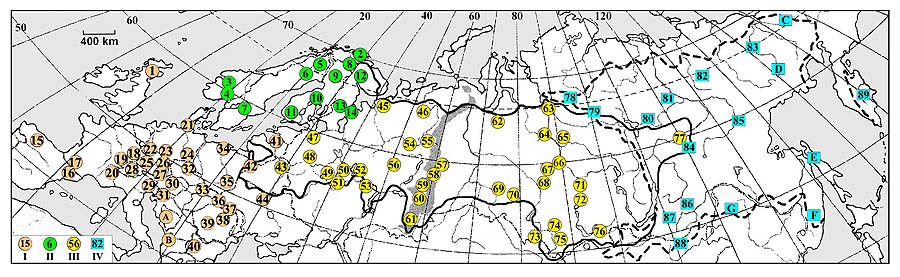
Fig. 1. Locations of present and some extinct (after ~1930 years) populations of capercaillie for which the indexes of air temperature and precipitation were evaluated. А – boundaries of the geographical range of Tetrao urogallus (not indicated in western Europe due to high fragmentation), Б – the ones of T.parvirostris. The population groups: I – from southern and central parts of Western Europe (T. u. major, T. u. cantabricus, T. u. aquitanicus, T. u. rudolfi), II – fennoscandian (T. u. urogallus, T. u. lonnbergi, T. u. karelikus), III – all other of T.urogallus, IV – all populations of T.parvirostris. Figures in circles and squares denote the serial numbers of populations: 1 – East Scotland (Aberdeenshire, Aberdeen, Angus, Clackmannanshire, North Lanarkshire, East Lothian, West Lothian, Midlothian, Mray, Prert-and-Kinross, Fife, Falkenre); Norway: 2 – Finnmark, 3 – Buskerud, Ostfold, Akershus, Oppland; 4 – Buskerud, Ostfold, East-Agder; Sweden: 5 – Lapland, 6 – Norbotten, Westerbotten; 7 – Gävleborg, Stockholm, Dalarna, Uppsala, Vermland, Örebro, Vestmannland, Södermanland, Ostergotlands, Vestra Gotaland, Kalmar; Finland: 8 – Reserve Värriö (north-east Lapland), 9 – Lapland and north-east. Oulu, 10 – south-west of Oulu, Vaasa, Kuopio, 11 – increasingly southern provinces; Russia: 12 – Lapland Reserve (Monchegorsk and Kovdor districts of Murmansk region); 13 – Republic of Karelia (whole); 14 – the Ileksa river basin (Onega district, Arkhangelsk region); Spain: 15 – Cantabrian Mountains (Asturias, León, northern Palencia, northern Zamoro, northern Burgos, Cantabria, northwest of Galicia); Spain and Andorra: 16 – Navarre, Aragon, Catalonia; Andorra; France: 17 – Pyrenees (Atlantic Pyrenees, High Pyrenees, Upper Garonne, Ariège, Aude, East Pyrenees); 18 – Vosges (Belfort, Upper Rhine, Lower Rhine, Vosges, Meurthe and Moselle, Moselle); 19 – Jura Mountains (Du, Jura, En); 20 – Alps (Savoy, Haute-Savoy); Germany: 21 – Northern Schleswig (Schleswig-Holstein); 22 – Sauerland (North Rhine-Westphalia); 23 – Hessen; 24 – Saxony; 25 – Baden-Württemberg (Freiburg, south of Karlsruhe); 26 – Odenwald, Spessart, Rien: Hesse, Rhine-Pfalz, Lower Franconia; 27 – Bavaria (Bavarian Alps, Bavarian Forest); 28 – Switzerland (only the alpine part: Vau, Berne, Valais, Ticino, Graubunden); 29 – Italy (eastern Alps: Lombardy, Trentino-Alto-Adige, Veneto, Friuli-Venezia-Giulia); 30 – Austria (Carinthia, Tyrol, Salzburg, Styria); 31 – Slovenia (whole country); 32 – Czech Republic (only mountain areas: Zlinsky, Moravian, South Moravian, Hradec Králové, Karlovy Vary, Liberec, Ustecky, Moravian-Silesian, Olomouc, Pardubice, Pilsen, South Bohemian districts); 33 – Slovakia (only mountain areas: Preshkovsky, Zhilinsky, Bansko Bistritsky, Trencinsky, Trnavsky, Nitran districts); Poland: 34 – Pomerania (Pomerania and West Pomerania voivodeships); 35 – Lubelskie, Сwiętokrzyskie voivodeships; 36 – Ukraine (Carpathians: Ivano-Frankivsk region); 37 – Ukraine / Romania: Bukovina (Chernivtsi region of Ukraine, Suceava judet of Romania); Romania: 38 – Northern and Eastern Carpathians (Alba, Vrancea, Covasna, Mures, Baczu, Ramatz, Hargita, Maramures, Suceava, Bistrita-Nasoud); 39 – Southern Carpathians (Arges, Brasov, Buzau, Valcea, Gorzh, Dymbovitsa, Karash Severin, Prahova, Sibiu, Hunedoara); 40 – Bulagaria: Western Rhodopes (Pazardzhis region); 41 – The Baltic States: Estonia (whole), Latvia (Vidzeme, Zemgale, Kurzeme, Latgale), Lithuania (Alytus, Vilnius, Kaunas, Klaipeda, Marijampolė, Panevėžys, Tauragi, Telšiai, Utena, Siauliai districts); Belarus: 42 – Reserve Belovezhskaya Pushcha (Kamenetsky, Pruzhany district of Brest region., Svisloch district of Grodno region); 43 – Minsk, Vitebsk regions; 44 – Ukraine: Polessye (Volyn, Rivne, Zhytomyr, Kiev, Chernigov, Sumy regions); Russia: 45 –White Sea Peninsula (Primorsky, Pinezhsky, Mezensky districts of Arkhangelsk region); 46 – North of Komi Republic and headwaters of the Vym river (Timan Mountains: Ust-Tsilemsky, Usinsk, Pechora districts); 47 – Leningrad region; 48 – Tver region; 49 – Moscow region; 50 – Ivanovo region; 51 – Oka Reserve (Spassky, Klepikovsky, Kasimovsky districts of Ryazan region); 52 – Nizhny Novgorod region (all left bank of the Volga river); 53 – Republic of Mordovia (Temnikovsky, Ichalkovsky, Bolsheignatovsky, Ardatov, Atiashevsky, Chamzinsky, Dubyonsky, Bolshebereznikovsky districts); 54 – south of Komi Republic (Syktyvdinsky, Kortkerossky districts); 55 – Pechora-Ilychsky reserve (Troitsko-Pechorsky district, Komi Republic); 56 – Kirov region (Slobodskoy district); 57 – north of Sverdlovsk region. (Nizhneturinsky, Verkhotursky, north of Alapaevsky, Turin, Tavdin and increasingly northern districts); 58 – south of Sverdlovsk region (all other southern districts); 59 – South Ural Reserve (Beloretsky district of Bashkiria, Katav-Ivanovskiy district of Chelyabinsk region); 60 – Kuragchinsky, Arkhangelsk Regions of Bashkiria; 61 – Bashkir reserve and south of Bashkiria (Burzyan, Abzelilovsky, Beloretsky, Kugarchinsky, Zilair, Zianchurinsky districts of Bashkiria); 62 – Ob-Taz watershed (Shuryshkarskiy, Priuralsky, Nadymsky, Purovsky districts of Yamal-Nenets Autonomous Okrug); 63 – lower reaches of the river Kureika (Turukhansk, Ilimpiysky, Dudinka districts of Krasnoyarsk Territory); 64 – Turukhansk district of Krasnoyarsk Territory; 65 – Turukhansky, Evenkiysky districts of Krasnoyarsk Territory; 66 – basin of the river Sym (Yenisei district of Krasnoyarsk Territory); 67 – basin of the river Lisitsa (Verkhneketskiy district of Tomsk region.); 68 – Baraba and Kulunda (Parbigsky, Bakcharsky districts, Tomsk region); 69 – Kyshtovsky district of Novosibirsk region; 70 – Ust-Tarsky, Wengerovo, Kuibyshev, Ubinsky, Kargatsky, Chulymsky and all more northern and eastern districts of Novosibirsk region; 71 – south of Yenisei plain (Ermakovsky, Karatuzsky, Minusinsk, Kuraginsky, Krasnoturansky, Idrinsky, Partizansky, Sayansky, Irbeisky, Novoselovsky, Balakhtinsky, Berezovsky, Mansky districts of Krasnoyarsk Territory); 72 – outskirts of Krasnoyarsk town; 73 – Kazakhstan: Glubokovsky, Leninogorsk, Zyryanovsky, Katon-Karagai, Ulan, Kokpekty, Kurishim districts of East Kazakhstan region; Russia: 74 – Biysk district of Altai Territory; 75 – Altai Reserve (Turchaksky, Ulagansky districts, Republic Altai); 76 – Piy-Khem, Kaa-Khem, Kyzyl, Todzhinsky kojuuns of Republic Tyva; 77 – middle flow of the Vilyui River (Suntarsky district of Yakutia); 78 – Plateau Putorana (Taimyrsky, Evenkiysky districts of Krasnoyarsk Territory); 79 – the Kochechum river basin (Evenkiysky district of Krasnoyarsk Territory); 80 – middle flow of the Vilyui River (Mirninsky / Suntarsky districts of Yakutia); 81 – the Kenkeme river basin (Gorniy district of Yakutia); 82 – Verkhoyansk district of Yakutia; 83 – middle flow of the Kolyma River (Nizhnekolymsky district of Yakutia); 84 – the Tokko river basin (Olekminsky district of Yakutia); 85 – lower flow of the Maya River (Ust-Maysky district of Yakutia); 86 – Zabaikalsky Territory (whole); 87 – Bauntovsky, Eravninsky districts of Buryatia; 88 – Mongolia: Cantiy (aimak Hentiy); 89 – Russia: Karaginsky, Ust-Kamchatsky, Milkovsky, Elizovsky, Ust-Bolsheretsky districts of Kamchatka region. Added points (climatic models were not developed, see text). A – Bosnia Herzegovina: Chukhovitsy (Herzegovina-Neretva canton); B – Albania: Arrt (Dibra region); Russia: C – Bilibino district of Chukotka; D – Srednekansky district of Magadan region; E – Tymovskiy district of Sakhalin region; F – Kirov district of Primorsky Territory; G – China: Jagdachi (Inner Mongolia)
The coordinates of population were found in literature or determined by spatial reference points denoted in it, by means of which the spatial contour of a population area and its geographical center was estimated. Latitude and longitude of the center were read off from the images of the program Google Earth.
In a lot of cases in literature not the places occupied by populations were indicated, but regions where they are located. In such a case the whole territory occupied by wood-grouse was included into the population area. For example, they were the whole Karelia, but only the North of Tuva inhabited by wood-grouse. In some cases the close arenas were characterized in different sources, and we considered them as a single territory. For example, all the references to Livonia, Kurland, Estonia, “Baltic Coast” etc. were united into the region comprising three modern Baltic states ( further, it is called Baltics). When referencing to different though overlapping spaces, e.g., the regions of southern Norway (see Fig.1, №3 and 4) or Krasnoyarsk region (see Fig.1, №64 and 65) we considered them as different populations. In one case the boundaries of the whole region - Swedish Lapland - were undefined. Further it was considered as a part of the fief Norrbotten lying to the North of the Polar circle. When referencing to the locations of populations in a river basin (the Sym river, Tokko river etc.) the whole or almost whole basin was included into the area. In more details (“middle Kolyma”, “lower riches of the May river, etc.) the relevant third of the basin was included into the area. Possible shifts of populations’ centers (maximum up to 2.0-2.5%, usually much less) due to these uncertainties within the whole area of T. Urogallus and even T. Parvirostris are insignificant.
Altitudes occupied by each population were characterized by a single indicator: some average level for each population. It was evaluated by literary sources, or if it was impossible, by the sets of altitudinal estimates read off from the images of the program Google Earth (see details below). The number of estimates (points) for each space holding a population changed depending on its size and terrain: on the plains - 50-380 points, in the mountains – 140-700 points. After excluding the points lying outside the forest boundaries 12.6 thousand estimates (points) were included into calculations.
Climatic parameters
Model design
One (and the only) information source for model design significantly reduces the influence of climate oscillations on certain estimates. However, it does not eliminate the problem. Therefore, unified and the only data set was used as the main information resource (Climatic…, 2017). On its pages there are: 1) geographical names of certain points, 2) their altitudes, 3) average annual air temperatures and 4) annual total precipitation. These two last values were received by the modeling of weather data (method is not given) “from thousands meteorological stations” from 1982 to 2012 (Website data source). There are no geographical coordinates of the points on the website, therefore, every point taken from it was found according to its name in the images of the programs Google Earth, Yandex-maps or OpenStreeMap (2017), there the values of latitude and longitude were read off.
The values of both temperature and precipitation (certainly both parameters) were taken for each point included into the analysis. The number of points used to build a particular model changed depending on complexity and the area of a region: from 48 till 376. Their total sum was about 5.6 thousand. Totally 69 models of temperature and 67 models of precipitation were developed for the assessment of climatic parameters by the geographical location of 89 populations of wood-grouse.
To provide more comprehensive idea about the whole generic area of wood-grouse, some peripheral points (with letter symbols, see Fig.1) were added to the analysis. For these points models were not developed: each point was presented by the long-term data from the only meteorological station (Climatic…, 2017). Altitudes of each of these stations may not be adequate to the average altitudes of wood-grouse’s regional habitats, but undoubtedly lie within their limits.
Boundaries of thermohumid space in North Eurasia
Although the distribution of wood-grouse is mainly determined by forest environment (see above), we tried to define more exactly the boundaries of his available thermohumid space, i.e. the boundaries of this space within the land area of North Eurasia. Since there are a lot of steppes and deserts (hot and arctic) in the arid sector of the space (precipitation
Model Verification
Different sources can give various values of temperature and precipitation for the same points. The possible reasons are that their compilers used non-matching chronological segments, data from different meteorological stations (distinct heights) of the same settlement etc. Further these distortions are considered as information noise. To measure it we matched the data of the same points (n=27) taken from two different sources (Thermograph…, 2017; Clima…, 2017).
The same two websites were used to verify the model adequacy. To them we added the data from seven meteorological stations located on the Russian plain received from our colleagues (see Acknowledgements). For the temperature models validation a total of 100 geographical points were used, for precipitation models validation – 97 points. Each of them was characterized by the values of temperature, precipitation, latitude, longitude and altitude. All these estimates were used in model development and considered as expected or control.
From 69 temperature models and 67 precipitation ones only 49 most problematic models were tested: mountains and/or large area regions. Models received for small flat regions (Leningrad region, Tver region, Moscow region etc.) were not tested.
Methods
Altitudes in population centers
These parameters were assessed by two methods. The first one was used only for middle- and high-mountain regions. From literature the data on height-distribution of wood-grouse were chosen in certain mountain systems: extreme or preferred levels were usually provided. The altitude of a whole population was assessed as an average of two values of this or that interval. The notices about vertical migration of birds, gender preferences for high-rise tiers etc., unfortunately, were not taken into account. When there were several publications concerning the same population with different data on occupied heights, interval of overlapping values was used. If there were no data on altitudinal distribution of wood-grouse, the data on the height interval of forest vegetation were used. It was stated according to general geographic (Dobrynin, 1948; Mil’kov, 1964; Gvozdetsky, 1976; Gvozdetsry,Mikhaylov, 1978) or regional sources (Chernov, 1964; Vasil’yev et al., 1980; Romanov, 2004; Gorichev, 2008 etc.), electronic reference books and personal messages of colleagues.
The second method was used when working with plains and low-hill terrains. In the program Google Earth geographic grid lines were applied to the image of a whole regional area of wood-grouse, and in several cross points the altitude were read off: the points were included into samplings at regular intervals. The average of these estimates characterized the altitude for the whole population. The same method was used when working with mountainous regions if only the height interval of forest vegetation was received. In fact, by this method the average altitude occupied by wood-grouse in the forest belt of the mountain system was assessed.
When reading off altitudes the terrain representations were used in scale 1:400 000 - 1:700 000 for plains and 1:100 000 - 1:500 000 for mountainous regions.
Climatic models
The idea behind the development of these models consisted in obtaining the regional dependencies (formulae) of temperature and precipitation on the three spatial parameters (latitude, longitude and altitude) only within population arenas of wood-grouse or for groups of some of them. From these formulae the values of temperature and precipitation for the center of each population shown in Fig.1 were calculated.
The work with the materials was reduced to the selection of some set of spatially mutually distant points sufficient to reveal regional patterns of changing temperature and precipitation depending on the arrangement of points included into the analysis. Since the spatial arrangement of points seemed to be extremely uneven (see Introduction), the special attention was paid to searching points with extreme values (a few out of hundreds looked through). These points in addition to the set of others gave the necessary ‘supports” for the approximating curves of our models.
All the regional models were built with the help of the multiple linear regression analysis (package Statistica 6.0), at that the series of climatic parameters were used as dependent variables and series of spatial estimates – as independent (explaining) variables:
Y = In + b · La + c · Lo + d · Al,
Where Y is value of temperature or perspiration, In – absolute term, La – latitude, Lo – longitude, Al – altitude; b,c,d – calculated coefficients for latitude, longitude and altitude. The attempts at non-linear modeling gave unsatisfactory results, and we did not return to them.
In the case of good approximation of dependent series by only one or two valuables (for example, only values of Al or only Lo and Al) one- or two-dimensional models, respectively, were applied, but in most cases they were based on all three valuables. If combined effect of two or three valuables was both reliable and strong, but that of one of them seemed to be unreliable, all two or three valuables were included into the model, provided that exclusion of unreliable effects did not lead to the increase in bond strength (control by R2adj ). When receiving alternative variants the models with the strongest bonds were preferable.
The temperature and precipitation models were developed separately for each of these parameters, but using the same regional data except in rare cases, when the initial analysis did not give reliable bonds or their strength seemed to be weak. In such a case we either increased or reduced the scale of the investigation. Thus, we could not find any reliable bonds for the Cantabrian Mountains (see Fig.1, №15) either for temperature or for precipitation. However, these bonds were revealed for the territories within this region: for the provinces of Asturia, Leon (together with the neighboring districts of Palensia, Samor and Burgos), Cantabria and Lugo. The models were built for each of four provinces, and the values of temperature and precipitation were calculated. The whole region (№15) was characterized by the average of four sub-regional estimates. However, more often we had to increase the area of the region. For the points № 8-11 (see Fig.1) the data throughout Finland as a whole were used, for №14 and 45 – throughout Arkhangelsk region, for point № 46,54 and 55 – throughout Komi region, for points № 42 and 43 – throughout Belorussia, for № 57 and 58 – throughout Sverdlovsk region, for № 63-66, 71, 72, 78 and 79 – throughout Krasnoyarsk region, for № 74 and 80-85 - throughout Yakutia.
For some regions a “mixed” approach was used, for example, Bashkortostan (№ 59-61) was divided into three parts: western, central and eastern. For the point № 59 the data were taken only from the north of central Bashkortostan ( with addition of adjacent areas of Chelyabinsk region), for the point № 60 – only from its western part, for the point № 61 – both from the East and center of the republic. The temperature model for Mordovia was built only by the data from its own territory, but for the precipitation model the additional data were applied from Ryazan region.
In the process of model verification it was found out whether control estimates go beyond the extreme values of variation series used to build these models. For example, the series used for modeling precipitation in Bavaria (n=82) at the height 1043 m gives the variation 634-1337 mm. But one of three control points of the region (Zugspitze, altitude 2960) goes beyond the height series and shows precipitation of 2004 mm, which exceeds the maximal value of precipitation series by 961 mm (deviation by 92%). When it is impossible to explain such deviations, the model was regarded as incorrect.
Results
Climatic models quality
All the temperature models were adequate to initial data (p<0.001) and strongly depend on locations (for 90% models r>0.9, for the rest r>0.7). Most precipitation models (83%) also demonstrated strong dependence (r>0.7), and middle strength bonds (r=0.40-0.69) were obtained for 14% of models and only two (3%) showed weak dependence: Italy (№ 29, r=0.25) and Austria (№ 30, r=0.22). Almost all the models were characterized by the high level of adequacy (p<0.001) except for four: Italy (p=0.001), Austria (p=0.005), Slovenia (№ 31: p=0.011) and Buryatia (№ 87: p=0012).
In the temperature model for northern Schleswig (№ 21) we managed to significantly increase the bond strength, when combined it with Pomerania (№34). But the reliable dependence of precipitation on the point locations in northern Schleswig was not revealed. For it the average precipitation (n=12) falling in the height interval 0-32 m was calculated, at that, the average region height was 16 m. For Trans-Baikal Territory (№ 86) the reliable bond by precipitation was not received either, and this parameter was also calculated as the average (n=12) for the height interval 900-1000 m, at that, the average height of the forest belt was 948 m.
When studying the information noise, it was shown that the values of temperature and precipitation obtained from different sources do not coincide, as it was expected. The greatest discrepancy was in temperature values: maximum up to 60%. But only for two points from 27 (8% of paired comparisons) the discrepancy values exceeded 20 %. As for precipitation, maximal discrepancies of any paired comparisons reached only 20%. Hence, the main noise level was within ± 20% of the absolute values from information sources. Therefore, in assessing the model correctness we neglected such discrepancies.
The temperature model verification showed that only four control estimates went beyond the limits of empiric variation series (models for the French Alps, Saxony, Westphalia and Bulgaria). One of the control estimates (for Bulgaria) was received for the altitude not getting into the interval of height series used to build given model and, hence, cannot testify to its incorrectness. The deviations of the rest three control estimates from the terminal values of the variation series were ± 0.2-0.5°C, which does not exceed 5% of terminal values used for modeling. It means that all real deviations fit into the framework of the information noise, and all the temperature models can be considered as correct.
Outside the variation series of our precipitation models there are 13 control estimates (for 12 models). Three of them characterize the points lying outside the limits of altitudes used to develop models (for southern Norway, Ivanov region and particularly for Bavaria, see above). The rest ten control estimates go beyond the terminal values of variation series by 6-177 mm, i.e., 1-17 %. Such deviations do not exceed the noise level and, hence, all the models can be considered as correct.
Two other approaches to functional verification of models is according to the deviation of the verified estimates from the curve approximating control values (with and without information noise) were also realized by us. They also showed that our models were correct. In this article their results are not presented (too cumbersome).
Temperature and precipitation within the generic area
Within the entire generic area of wood-grouse both studied variables – temperature and precipitation – showed the reliable dependencies on latitude and longitude, but precipitation also on altitude (Table 1). Besides, our samplings (populations in Fig.1: n=89) were characterized by the reliable relations between all geographical valuables: latitude and longitude (r=0.39 p<0.001), latitude and altitude (r=-0.68 p<0.001) as well as longitude and altitude (r=-0.21 p=0.05). Perhaps, it is such weak, latent relationships that cause significant problems in realization of a traditional analysis of adaptive reaction of birds on the axes of spatial variables (see Introduction), which we refused. The visual analysis of mutual arrangement of populations in the axes of temperature and precipitation is indifferent for these relations.
Table1. Dependence of temperature and precipitation on geographic variables within the generic area of wood-grouse
| Temperature | Precipitation | |||
| r | p | r | p | |
| Latitude | – 0.62 | < 0.001 | – 0.59 | < 0.001 |
| Longitude | – 0.86 | < 0.001 | – 0.67 | < 0.001 |
| Altitude | 0.17 | 0.113 | 0.55 | < 0.001 |
The entire, studied thermohumid space (Fig.2) was divided into four parts, and dividing lines were drawn through the centers of the intervals formed by the terminal values of regional climatic estimates. For all wood-grouse populations included into the analysis these terminal values were: temperature from -13.4 to +8.5°C, precipitation from 196 to 1426 mm. In the upper right quarter there are groups living in warm, humid conditions, in the lower right quarter – those living in warm and dry conditions, in the lower left one – those living in cold, dry conditions. The space in the upper left quarter (cold and humid conditions) appears to be nearly non-existent on land of North Eurasia. Although in the upper right quarter this space goes beyond the limits indicated by us and spreads (as temperature rises) along the precipitation axis form 1200-1300 ( some points in Norway and on Sakhalin island) to 2000-3100 mm/year (Norway, Switzerland, Bavaria, Austria, Italy). The multistep character of the whole work (see Introduction) and low quality of the description of population areas in some sources allow considering the final results of modeling only as an approximation to reality. Therefore, the final estimates are not supplied with variation indexes (see Fig.2), which could evoke the illusion of precision, what is not in reality.

Fig.2. Location of habitats of some capercaillie populations in axes of air temperature (X) and precipitation (Y) (calculated values). The population groups: I – in southern and central parts of West Europe (T. u. major, T. u. cantabricus, T. u. aquitanicus, T. u. rudolfi), II – fennoscandian (T. u. urogallus, T. u. lonnbergi, T. u. karelikus), III – all the rest of Tetrao urogallus, IV – thermohumid space seems to be absent on land of North Eurasia. Dotted line is the boundary of group I. The signs next to circles and squares are the same as in figure 1
The overlapping area of the thermohumid space of common and rock wood-grouse is essential, what the terminal species values indicate: from -8.5 to +3.0°C on temperature axis. And such a scope occupies 52-53% of the whole generic area on the temperature axis. The overlapping on the precipitation axis is from 260 to 839 mm (47%). In the zone of overlapping the distribution of separate populations is of interest. In the quarter of cold and dry climate there is point № 8 (see Fig.2) occupied preferable by rock wood-grouse. This point lies in the North-West of Lapland, only about 250 km from the sea coast. The temperature estimate for the population of T. Parvirostris from Primorsky Territory (point F, see Fig.2) is located not far from the estimates for the group of T. Urogallus from Leningrad region (№ 47), and even for those from northern Norway (№ 4). Kamchatkan population (№89) dwells in approximately the same conditions of precipitation as the group of T. Urogallus from Ukrainian (№ 36) and Romanian Carpathians (№ 38), and even from Germany (№ 26). The precipitation seems to be close for such distant regions as Yakutia (№ 80, 81,85), southern Siberia (№ 86,87) and Mongolia (№ 88) from one side, and Swedish Lapland (№ 5), from the other side.
Despite the scatter of estimates, interspecies differences are visible: T. Urogallus mainly inhabits warmer both dry and humid regions, while T. Parvirostris - colder and mainly dry ones. Only two populations of T. parvirostris - from Kolyma (№ 83) and Chukotka (point C) - go beyond the lowest (dry) limit of the area of T. Urogallus (№ 75, see Fig.2). The space with the precipitation > 600 mm inhabited by this species appears to be limited. In our analysis only three points: Kamchatka (839 mm, № 89), Primorye (702 mm, point F) and Sakhalin (662 mm, point E) were included. It is possible that the area of rock wood-grouse mainly occupies a dry part of the area of T. Urogallus , almost completely fitting within the limits of the latter and occupying its smaller part.
The obtained estimates allow partly delimiting even the areas of subspecies. Within the thermohumid space of T.urogallus some West European subspecies: T. u. major (23 points: № 1, 18-35, 37, 42, 44 see Fig.2), T. u. cantabricus (№ 15), T. u. aquitanicus (№ 16, 17) and T. u. rudolfi (№ 36, 38, 39, 40) form territorially integrated, whole group. As quite expected, they occupy the warmest and wet part of the whole area of genus Tetrao, and in Fig.2 they are separated from the rest subspecies with dotted line. The warmest part of the area is occupied by four German populations (№ 21, 23, 25, 26, see Fig. 2), the coldest – by the groupings from Romania (39), Austria (30), Slovakia (32) and Czechia (33). In the wet part - with maximal precipitation for the entire generic area - there are populations from Yura ((№ 19), the French Alps (20), Switzerland (28) and Slovenia (31). The thermohumid area of West European group appears to hardly overlap with the similar areas of other subspecies (see Fig.2). Conversely, the integrated thermohumid space of three fennoscandian subspecies (T. u. urogallus, T. u. lonnbergi, T. u. karelikus, 13 points: № 2–14, see Fig. 2) significantly overlap with the space not only of East European, the Urals’ and West Siberian subspecies (№ 43, 45–62, 67–70), but it approaches the central Siberian conditions ( the vicinity of Krasnoyarsk, № 72). We fail to delineate the own thermohumid space only of fennoscandian group (without other subspecies) with continuous or weakly windy line.
In the area of rock wood-grouse the Kamchatka population (T. p. camtschaticus, № 89, Fig. 2) holds a special position. This population reaches the area of distribution of West European subspecies of T. urogallus on precipitation scale and lies in warmer zone than, for example, all the fennoscandian populations except one of Norway (№ 3). T. p. stegmanni (points 86–88, G) seems to be located in the dry periphery of the warm part of the species area. The location of T. p. Parvirostris was not defined, since for the inhabited by this population Khabarovsk and Primorye Territories the regional models were not developed.
For the whole generic area a direct, but not linear relation between temperature (x) and precipitation (y) was observed: rs = 0.77 p < 0.001, i.e., the warmer the habitat, the more precipitation falls there. The general distribution of wood-grouse in the thermohumid space can be described with the dependence of precipitation on temperature: у = 605.78 + (32.94х) + (0.70х2). The refinement of the general picture shows that the non-linearity of this relationship appears due to two contrary trends. For the thermohumid area of West European group of subspecies a linear inverse relation is characteristic: the warmer, the less precipitation (Fig.3), at that the elimination of plain populations from the calculation leads to the loss of its reliability: r = – 0.07 p = 0.78. The spaces of the rest subspecies show a direct linear relation between warmth and precipitation: the warmer, the more precipitation. These relations are not strong, but reliable. For the fennoscandian group: r = 0.61 p = 0.03; for all other subspecies of T. urogallus dwelling to the East of Brest, Saint-Petersburg and the Onega: r = 0.48 p = 0.004; to the whole area of T. parvirostris: r = 0.70 p = 0.02. It is quite expected that the whole territory occupied not by West European subspecies of common wood-grouse , but all the subspecies of rock wood-grouse also showed a direct relation between warmth and precipitation: r = 0.64 p < 0.001. Hence, the pattern of spatial changes of the two most significant climatic parameters (their integrated gradient) in the area of West European subspecies dramatically differs from the similar pattern in the territory of all the rest subspecies of the genus Tetrao: in the first case precipitation increases while temperature reduces, in the second one the former reduces.

Fig.3. Relation between air temperature (axis X) and precipitation (Y) (calculated values) within the area of T.urogallus in southern and central parts of West Europe (R2 = 0.267 r = –0.516 p = 0.004; y = 1393.3 – 78.3 x). I is mountains (altitudes 700–1825 m), II – plains and lowlands (16-485 m). The numbers next to circles and squares are the same as in figures 1 and 2
It can be noted that while temperature increases the area of Tetrao seems to extend on the axis of precipitation (see Fig.2). To test this assumption we compared the values of ranges in precipitation variation, where the birds’ populations are located, with the correspondent temperature estimates dividing them into the 1°C intervals, i.e. from –13.5 оС to –12.5 оС, from –12.4 оС tо –11.5 оС… and from +7.5 оС to +8.5 оС. The results confirm the assumption: temperature determines the precipitation variation in the habitats of the whole genus at 45% (r = 0.67 p = 0.003), in the habitats of T. urogallus – at 55% (r = 0.75 p = 0.003); for T. parvirostris any reliable relation was not revealed, perhaps, due to small sampling. At that, for all the populations included into the analysis the maximal values of the precipitation interval as a whole increase faster (y = 843.6 + 42.0x; r = 0.81 p < 0.001, where x – temperature, y – precipitation), than the minimal ones (y = 482.6 + 16.7x; r = 0.88 p < 0.001). Hence, in colder conditions of the environment the birds occupy the places with the narrower range of precipitation: low temperature seems to enhance the dependence of wood-grouse on precipitation, and in so doing limits the usage of the space.
Discussion
It is evident that within the population boundaries animals are able to choose the most optimal patches, which characteristics can differ from the average value for the whole population area. Therefore, using the average population values of temperature and precipitation in our work may have shifted somewhat the calculated estimates relative to reality. It is undoubtedly, however, these shifts can be significant only for the comparison of neighboring or close groupings, but almost insignificant at the level of large regions and even more subspecies and species areas.
The analysis of geographical and climatic variables showed the reliable, though not the strongest dependences between them (r = 0.55–0.86, see Table 1), it also revealed the relations between the geographical variables themselves (r = 0.21–0.68, Results). Even these scanty facts testify about the problems in the interpretation of the results when only geographical variables are used to study the adaptation of birds (see Introduction). In this case one can expect the impact of a number of significant and minor factors, which are completely or partly accumulated by the following variables: latitude, longitude and altitude. Our approach leaves no doubts: it is in the axes of temperature and precipitation that the mutual arrangement of wood-grouse populations is described.
The general picture of variation of these values within the boundaries of the whole generic area seems quite adequate. The transfer of air masses from the West over Eurasia causes the reduction of precipitation and winter temperatures with the distance from the Atlantic Ocean (Alisov, Potarus, 1974). It makes quite logical the location of most West European populations in the warmest and wet region of the area, most East Siberian – in cold and dry region, the rest groupings occupy intermediate position (see Fig.2).
The fact that wood-grouse does not occupy the regions with low temperature and a large amount of precipitation (upper left quarter in Fig.2) is caused by the trivial absence of such type of thermohumid space: cold air has less water capacity that warm one. The same factor dictates the significant and sharp contraction of a potentially available space also in the left lower quarter (cold and dry). This gap is most likely to mark the territories of plain tundra and mountain vegetation above the forest line: in cold conditions the rising humidity inhibits forest vegetation (Puzachenko, 1985) and blocks the wood-grouse distribution, since this genus is presented by dendrophilous species (see Introduction). It appears that the narrow thermohumid space of rock wood-grouse on precipitation axis is determined by quite small size of this space really existing in cold zone of North Eurasia. In this sector the development of treeless communities additionally limits the thermohumid space potentially usable for birds. Only two populations from the opposite parts of geographical generic area: from the plateau Putorana and from the mountain regions of northern Tuva (points 78 and 76, see Fig.3) are located on its boundaries. On their slopes indeed there is an increased amount of precipitation (Gvozdetsky, Mikhailov, 1978). This is likely to be one of the most eastern picks of precipitation due to both orography and transfer of air masses from the West. The significant amount of precipitation in Kamchatka, Sakhalin and Primorye (points 89, E, F) is caused by the influence of the Pacific. Thus, the relatively narrow the thermohumid space of rock wood-grouse comparing with common one is the consequence of objective reasons: in the area of the first species the environmental conditions are more uniform. And T. parvirostris seems to utilize the entire, potentially usable space. There is only one limitation concerning the thermohumid conditions: the presence of forest environment.
The geographical areas of common and rock wood-grouse overlap (see Fig.1), and their thermohumid spaces also overlap as indicated by us (see Results). However, in the axes of temperature and precipitation this overlapping seems to be very large. So, the location of terminal points of the area of T. parvirostris in these axes (№ 89, E and F, see Fig. 2) indicates to its potential ability to utilize the vast territories occupied today only by common wood-grouse, including Middle and West Siberia (№ 67, 71, 74), the Urals, the piedmont of the western Urals (54–56, 58, 60), and even a lot of regions of Fennoscandia (2, 9, 10, 12–14). It is quite possible that the modern adaptive potential of T. parvirostris to thermo-humid environmental conditions quite allows it to occupy the significant part of a vast space of the modern distribution of “Russian” and fennoscandian subspecies T. urogallus (groups I and III in Fig.1). On the contrary, the potential spread of T. urogallus in the thermohumid space of T. parvirostris seems limited: maximum the south-eastward of Yakutia (№ 85), northward, north-eastward and even south-eastward seems to be excluded.
Hence, the zone of interspecies overlapping of thermo-humid space shows the evident disparity of two species of the ability to potentially settle: they are great in rock wood-grouse and small in common one. These differences allow the assumption that the eastern boundary of T. urogallus area is located not far from the limiting line, where its species adaptations to the studied variables seem exhausted. For further northing, easting and south-easting the additional means, which are likely to be outside the species possibilities, are necessary. It is not excluded that the issue of eastern boundary of T. urogallus will be resolved in more detailed study of adaptation to thermohumid environmental conditions. On the contrary, modern knowledge of T. parvirostris biology (Isaev, 2014) does not show the evident abiotic or cenotic limitations for species expansion westward, i.e. to more favorable conditions (softer climate, more diverse vegetation). Factors blocking this expansion seem very unclear at the level of modern knowledge. It is possible that one of them is interspecies relationships: in the cohabitation places the common wood-grouse squeezes the rock one (Kirpichev, 1974).
The mutual arrangement of some populations in thermohumid space of T. urogallus seems somewhat unexpected. Thus, the maximal amount of precipitation should be expected in the terminal western regions, i.e. for Scotland (№ 1, see Fig. 1), Norway (№ 3, 4) and the Cantabrian Mountains (№ 15). However, the calculated values showed the moderate estimates for these points and their proximity to the line dividing the entire data set into dry and wet regions (see Fig.2). Among the considered population arenas the Jurassic Mountains (№ 19, Fig. 2), the French Alps (№ 20), Switzerland (№ 30), Slovenia (№ 31), Bavaria (№ 18) and the Pyrenees (№ 16 and 17) receive the maximal amount of precipitation.
The plain areas of France do not prevent the penetration of humid air in the interior of the continent. Well expressed orographic genesis of the great amount of precipitation in this region is seen on Vosges (№ 18), which is the low but the first barrier on the way of prevailing western winds. Vast and high mountain groups – the Pyrenees and the Alps on the way of humid sea air masses receive the most precipitation. The same masses moving over the low mountains of Scotland (maximum< 1400 m) seem to lose most of moisture on western slopes (see Moss, 2015). This causes smaller precipitation in eastern Scotland as compared to the Alps and the Pyrenees. However, the population which is not included in our analysis of populations of western Scotland and, possible, south-western Norway still exist in the conditions of greater precipitation than those of in the Alps and the Pyrenees.
Despite expectation assuming the increase of temperature on the planet from poles to equator, the warmest places inhabited by wood-grouse (> +7.9°C) seem to be the regions of Germany: (№ 21–26, see Fig. 3).These not the southernmost points (see Fig.2) are located at the low hypsometric levels (16- 483 m). It may be the main reason of high temperature estimates obtained for them. European populations from southern latitudes (№ 15–17, 20, 29, 36–40) are located much higher (1200-1825 m) and it is quite logical that they receive less warmth: from +3.2 to +7.7°C (see Fig. 3). That is the factor “altitude” completely neutralizes the effect of the factor “latitude”.
The variety of conditions in the habitat of fennoscandian populations (№ 1–11, see Fig. 2) is striking. They are located geographically densely (see Fig. 1) and cover almost the whole thermal interval occupied by the populations of Russian and West Siberian plains (№ 49–66, see Fig. 2). On the axis of precipitation fennoscandian groupings occupy the space holding almost all the populations of T. urogallus , except for some West European ones. Such a variation of our estimates for Fennoscandia is an obvious consequence of the greatest diversity of its climatic conditions.
Several points located both in the warmest and in the relatively dry region of the generic area should be highlighted: Saxony (№ 24, see Fig. 3), Pomerania (№ 34), Lublin (№ 35), Belovezhskaya Pushcha (№ 42) and Ukrainian Polesye (№ 44). This territory was most likely to be considered as a habitat of wood-grouse males with maximal weigh in the whole area (Zedlitz, 1924; Couturier, Couturier, 1980) and was opposed to the populations with small individuals mostly from the Austrian (№ 30) and Bavarian (№ 27) Alps (Zedlitz, 1933). Our results allow us to assume that one of the reasons the noted weigh differences can be not only and not so much interregional discrepancy between the estimated temperature (just by 1.5-2.0°C), but almost twice the difference of precipitation (580-605 mm vs. 1140-1180 mm, see Fig.3). Apparently, the combination of the two factors can directly or indirectly influence the growth and development of wood-grouse. Its optimal habitat conditions are probably in the region of relatively high temperature and small precipitation. However, that can be true only for the group of West European subspecies or only for T. u. major.
The negative effect of the increased amount of precipitation on wood-grouse is usually considered from the perspective of chick survival and population renewal (Teplov, 1947; Gavrin, 1956; Lindén, 1981; Ménoni, 1991; Watson, Moss, 2008). Our results showed that in its habitats the precipitation interval diminishes when temperature decreases. This fact allows us to assume the general negative effect of humid climate on the individuals of genus Tetrao, including adult specimens: they are also negatively influenced by a large amount of precipitation, mostly rains. For example, through increasing heat input to dry feathering and/or to diminish time for foraging because of the need to hide from heavy rains and for some time after.
The interesting finding of our study was the fundamental difference between the area of West European subspecies group (T. u. major, T. u. cantabricus, T. u. aquitanicus, T. u. rudolfi) and, perhaps, all the populations of genus Tetrao in the patterns of spatial change of temperature and precipitation. The main reasons for this can be the relatively small area of West European area and strong variation of the terrain within it. The first factor allows latitude patterns of climate variability as well as all the consequences of western air masses transfer in Eurasia to develop only to a lesser degree. Both these patterns seem not to be noticeably detected due to a trivial lack of place, the arena of their realization in West Europe. In this situation the role of terrain increases: the higher the mountains, the less warmth and more moisture. Apparently, it is the high heterogeneity of the terrain in West Europe that is the main reason for the discussed differences, since for area of fennoscandian group inhabiting mainly plains and low-hill terrains another pattern peculiar to vast spaces of the central and western parts of generic areal is noted. Within their limits the air masses circulation along longitude moves aside the effect of even large mountain systems and demonstrates direct relation between temperature and precipitation: the East and the farther from the Atlantic with the Gulf-Stream, the less warmth and the less moisture.
These patterns are uninteresting for the systems with rigid fixation of objects in space. But birds are mobile, and migrations, including distant ones, often purposeless can take them out into pessimal environmental conditions, i.e. cold ones. It is important that living organisms usually endure cold in dry conditions easily. In such context the spatial differences in thermohumid variations revealed by us are of particular importance. To demonstrate it let us separate two types of climatic gradients. One of them is characteristic for West, central and South Europe as a whole: the shift from high temperatures to low ones is accompanied by the increase of precipitation, which additionally (to low temperature) complicates the conditions of existence. Since the exaggeration of environmental conditions requires the increasing energy expenditure for life (Dol’nik, 1995), let us call such a gradient “rigid”. The opposite type – “soft” gradient – is characteristic for the most places of North Eurasia: downward shift in the temperature scale is accompanied by increasing dryness, which has to determine less noticeable deterioration in living conditions; there are no significant climatic barriers in its colossal spaces, changes are smooth and gradual.
It should be recalled that hierarchy of spatial structure of populations requires the universality of group adaptations: for the entire population the selection makes stable only those which are adequate through its areal. Others, less important adaptations are viable at local levels (Severtsov, 2013). Consequently, in the conditions of rigid environmental gradient even limited spatial expansions must be accompanied by significant adaptive restructuring. The soft gradient is likely to be more tolerant to immigrants and requires noticeable adaptive shifts of them only in the colonization of very distant spaces.
In the conditions of extremely rigid gradient local adaptations must be clearly expressed, the spaces utilized by their carriers – small, and populations themselves have to be relatively isolated groupings of specialists-stenotops. Interspecies crossing and inbreeding are quite real, since distant migrations are successful only for specimens with the broadest rate of reaction. The opposite extreme (the softest environmental gradient) proposes functionally close local and more general climatic adaptations. The intraspecific spatial structure should be smooth with fuzzy boundaries of the adjacent territorial groupings and gradual spatial variations of genotypes. Interspecies crossing is possible only in the conditions of low abundance, but distant migrations are quite usual and can be successful for the wide range of individuals.
Such a view on climatic gradients allows us to understand, in particular, a strong variation in fennoscandian populations through the thermohumid space: migrations even from the regions of Siberia to European part of Russia and farther to the North-West, as well as in opposite direction do not require significant adaptive restructuring. Soft gradient can be used to explain a striking genetic unity of Russian wood-grouse in colossal space from Arkhangelsk and Tver to Krasnoyarsk (Duriez et al., 2007) as well as step-by-step changes of haplotypes: differences between West European (including Swedish) and Russian wood-grouses are greater than between Finnish and Russian ones (Liukkonen-Anttila et al., 2004). The differences in migration activity of wood-grouse increased in north-western Russia (Borchtchevski, Moss, 2014) or in Sweden (K. Sjöberg, cit. by: Åhlen et al., 2013) and quite moderate in the centre of West Europe (Storch, 1993) become clearer. Without any idea of different climatic gradients all these facts are only the list of unrelated phenomena. The proposed view allows us to notice a definite system in a jumble of such facts: various, possibly unusual in some way adaptive complexes (apparently, territorial) of behavioral, physiological, genetic and possibly other reactions of warm-blooded animals to divergent trends of habitat.
Unfortunately, the detailed discussion of thermo-humid gradients which must include the clarification of the boundaries of subspecies groups isolated by us and their lists is beyond the framework of the work. It should be noted that the phenomenon itself of multidirectional relations between temperature and perspiration which seems to remain beyond the view field of specialists, undoubtedly requires the verification and understanding.
Conclusions
The general arrangement of the centers of really existing and recently extinct populations of wood-grouse in the axes of temperature and precipitation looks quite natural: in the warmest and wet region of generic area the groupings of central and southern regions of West Europe are located, in cold and dry – most West Siberian ones, the rest populations occupy the intermediate position. Such an arrangement confirms the correctness of the most climatic models developed to describe general thermo- humid space of genus Tetrao.
Wood-grouse is almost completely absent in the area of negative average annual temperatures with the significant precipitation (>800-900 mm/year). This appears to be connected with the extreme limitation of such a type of thermohumid space on the land of North Europe.
When moving from maximal to minimal temperatures, the thermohumid space occupied by the genus shrinks on the precipitation axis, demonstrating the trend to wider usage of habitats with the greater precipitation and the growth of temperature.
The overlapping of species zones of the thermohumid space appears to be much wider than the overlapping of species geographical areas. The limits of the thermohumid space occupied by common wood-grouse approximately correspond to modern boundaries of its geographical distribution. It allows considering warmth and precipitation as limiters of its potential expansion in an easterly direction. However, the same factors seem not to prevent the counter expansion of rock wood-grouse in a westerly and north-westerly direction up to Fennoscandia.
Different populations of wood-grouse dwell in the conditions of mismatched spatial co-variation of warmth and moisture. In the area where the main part of generic population lives the amount of precipitation reduces when moving from warm regions to cold ones. However, according to the same gradient in the territory of the southern and central sectors of West Europe the amount of precipitation does not reduce, but increases. These differences not noticed earlier may be able to form the complexes of mismatched group adaptations. The revealed differences possibly will allow finding the explanation to known but inexplicable similarity and the differences of the spatial groupings of wood-grouse by means of genetics, morphology, ecology, spatial mobility and, perhaps, some other signs.
Ranging certain populations in the thermohumid space showed that, despite expectation, in the warmest conditions of generic area not southernmost grouping (Spanish, Balkan, Bulgarian), but populations of north-western Germany are located. Also contrary to expectation, the climatic models showed maximal precipitation not in the habitats of Atlantic groupings (Cantabria, western Scotland, Norway), but mainly in biotops of birds from north-western foot-hills of the Alps (Vogezy, Yura), from the central and southern regions of this mountain country (Switzerland, Austria, Bavaria, Slovenia, Italy), as well as from the Spanish Pyrenees. Although the populations from western Scotland not included in the analysis and possibly, from north-western Norway seem to exist in the conditions of maximal amount of precipitation. Fenno-scandian subspecies utilize the vast part of the thermohumid space penetrating into the dry zone of low temperatures occupied by “Russian” populations including Siberian ones.

 © 2011 - 2026
© 2011 - 2026