



Введение
Сегодня не подлежит сомнению определяющее влияние климата на важнейшие характеристики крупных биомов и многие экологические черты их обитателей. Поэтому четкое представление о силе проявления климатических факторов среды в ареалах животных необходимо для ясного понимания адаптивных реакций на их воздействия. Ряд таких реакций со стороны птиц на влияние климата удается выяснить с помощью ранжирования видов или популяций в осях пространственных переменных: широты, долготы и высоты местности (Dunn et al., 2000; Martin et al., 2006; McNamara et al., 2008; Bears et al., 2009; Lu, 2011; Hille, Cooper, 2015; Boyle et al., 2016 и др.). Изменения таксономических показателей размеров тела, плодовитости, смертности и других параметров рассматриваются как реакция на пространственные градиенты среды. В большинстве таких работ прямо или косвенно указывается на изменения конкретных климатических показателей по изучаемым градиентам, но также напоминается об адаптации любого вида или популяции ко всему комплексу внешних условий. И этот комплекс, кроме климатических факторов, включает биотические воздействия, среди которых обычно рассматривают риск хищничества, конкуренцию, популяционную плотность и т. д. (например, Donázar, 1990; Martin, 1995; Jetz et al., 2008). Обсуждаются также ценотические переменные: тип растительности и/или кормовые ресурсы (Donázar, 1990; McNamara et al., 2008; Boyle et al., 2016). К этому списку можно добавить трудно учитываемые и потому не обсуждаемые переменные: геохимический режим (трофность) ландшафта, а также антропогенные нагрузки, способные искажать естественные биоклиматические тренды, в том числе и на обширных пространствах (Равкин, Равкин, 2005). Деформация объясняющих переменных возможна, по-видимому, и при включении в анализ таксонов, ареалы которых занимают несколько континентов и/или представлены циркумполярными ареалами без какой-либо внутренней дифференциации таких ареалов (например, Balasubramaniam, Rotenberry, 2016).
Вычленить воздействие отдельных переменных из сгруппированного таким образом комплекса факторов почти невозможно, и не случайно весь подход рассматривается как эвристический (Hille, Cooper, 2015), дающий надежные заключения лишь для крупных таксонов (семейство и выше), обширных по площади территорий (тропики/умеренные широты) или по четко различаемым, например орографическим, признакам (Jetz et al., 2008; Bears et al., 2009; Lu, 2011; Boyle et al., 2016 и др.). Фактически этот подход вынуждает оперировать группами объясняющих переменных, часть из которых имеет неустановленные количественные значения, а некоторые переменные остаются даже неидентифицированными. Именно эта неопределенность – и перечня факторов, и силы их проявления – существенно затрудняет исследования: любая территория (административная область, природная зона, видовой ареал, его часть и т. д.) – это всегда комплекс трудно расчленяемых воздействий среды и индуцируемых им адаптаций. Мы попытались предельно конкретизировать и упростить анализ: принудительно сузить до минимума число объясняющих переменных и по ним ранжировать используемое птицами пространство.
Безусловно, климатические показатели меняются по широте, долготе и высоте местности (Алисов, Полтараус, 1974). Однако попыток адекватной трансформации этих трех переменных в климатические шкалы с целью изучения адаптивных реакций птиц, по всей видимости, не предпринималось: исследований широко распространенных родов или видов птиц-фитофагов в осях только климатических характеристик (любых) в доступной нам литературе найти не удалось. Наиболее вероятно, что отсутствие таких работ отражает превентивную реакцию исследователей на вполне ожидаемые эффекты коллинеарности. Реализация другого подхода – ранжирование популяций (ниже этот термин используется как синоним понятия «региональное население») в осях немногих известных переменных – позволяет проводить как минимум визуальный анализ взаимного расположения популяций по факториальным градиентам и оценивать степень воздействия на них со стороны вполне конкретных переменных среды.
Ниже представлена попытка оценить с помощью этого подхода воздействие основных климатических факторов на размещение широко распространенных видов птиц. В качестве примера взяты реальные популяции глухаря – обыкновенного Tetrao urogallus и каменного T. parvirostris. Их расположение в географических осях относится к тривиальным фактам. Но это расположение, скорее всего, трансформируется при замене географических осей на климатические. Однако предсказать характер размещения тех же популяций (например, в осях тепла и влаги) совсем не просто. Например, где теплее: на северных равнинах Евразии или на привершинных частях ее южных гор? Ответ не очевиден, как и многие другие. В пространстве экологических осей популяционные центры (точки) образуют компактное пятно, растянутся линейно или по неким кривым? Каким? Лягут они равномерно или кластерами? Будет ли такая кластеризация соответствовать таксономическим классификациям? Большинство подобных вопросов возникает из-за многопланового воздействия на животных со стороны рельефа. Особенно это касается горных регионов, что требует в том числе и уточнений высотных пределов растительности, от которых может зависеть распространение самих птиц. И эти пределы не всегда совпадают в разных горных системах (Алисов, Полтараус, 1974). Например, даже в соседних массивах (Альпы/Динары) глухарь занимает разные высотные ярусы (Čas, 2010).
Цели данной работы: 1) представить общую картину размещения реальных популяций глухаря в осях температуры воздуха и атмосферных осадков, 2) на основании ее визуального анализа оценить взаимное расположение популяций в этих осях, стараясь уяснить различия или сходства межгрупповых климатических адаптаций, не оставляя, по мере возможностей, без внимания генезис таких различий.
С первого взгляда достижение этих целей не должно вызывать особых затруднений: климатические оценки накоплены для огромного количества точек, а на случай их недостатка существуют интерполяционные алгоритмы. К сожалению, пространственное расположение метеостанций крайне неравномерно: в основном они сконцентрированы на территориях активной деятельности человека, которые, как правило, не заселены глухарем или занимают лишь периферию его популяционных арен. И для многих из них метеостанции – редкость. Кроме того, в условиях противоречивых трендов интерполяции ненадежны. Например, в горах, где локальные инверсионные явления могут искажать общий ход изменения температуры (чем выше, тем холоднее) противоположным трендом (Хромов, Петросянц, 2006). К тому же прохождение воздушных масс даже через невысокие горные системы способно диаметрально противоположным образом отражаться на склонах различных экспозиций: соседние наветренный и подветренный склоны одинаковой высоты даже небольшого по протяженности горного хребта обычно получают разное количество осадков, например низкогорья Шотландии (Moss, 2015). Внутри крупных горных систем (например, Альпы), над которыми могут формироваться собственные воздушные массы (Добрынин, 1948), возможно образование мелкой мозаики с сильнейшей вариацией осадков по локальным участкам склонов и долин (Beniston, 2006). По-видимому, рельеф определяет более высокую предсказуемость температурного режима региона, чем режима осадков, т. к. последний формируется бóльшим количеством переменных.
Сильные различия по характеру рельефа между популяционными аренами глухаря (горы/равнины) и дефицит точек с климатическими данными для многих арен делают рискованной интерполяцию оценок по неким общим алгоритмам. Разработка климатических моделей для каждой конкретной популяции (или их групп) представлялась более конструктивной. Пусть даже самых простых моделей, но индивидуальных. По изложенным выше причинам наша работа состояла из последовательного решения ряда задач.
- Для центра каждой принятой к анализу популяции выяснялись географические параметры: широта, долгота и высота территории.
- Для широкого пространства, вмещающего каждую популяцию (их группу), выбирались множества точек и для каждой такой точки устанавливалось пять характеристик: широта, долгота, абсолютная высота, среднегодовая температура воздуха (далее просто «температура», °C) и среднегодовая сумма атмосферных осадков (ниже «осадки», мм/год). Одновременное использование двух последних показателей во многом снимает вопросы нивелирования сезонных нюансов среднегодовыми оценками.
- По данным каждого множества точек, взятых только из внутренних контуров популяционных арен, рассчитывались индивидуальные модели (формулы), отражавшие региональные зависимости температуры и осадков (раздельно) от широты, высоты и долготы.
- Результаты моделирования проверялись с помощью климатических источников, не привлекавшихся для построения моделей.
- Проверенные модели использованы для расчета значений температуры и осадков для каждой популяции (ее центра).
- По этим расчетным значениям каждая популяция позиционировалась в осях температуры и осадков, плоскость между которыми ниже рассматривается как термогумидное пространство.
- Расположение точек (популяций) и/или их групп в этом пространстве охарактеризовано по результатам простого визуального анализа, реже с помощью обычных корреляционных или регрессионных процедур.
Напомним, что оба вида глухаря – неперелетные фитофаги. Зимой их трофические потребности удовлетворяются почти исключительно за счет древесной растительности, поэтому северные и южные границы их ареалов примерно совпадают с распространением лесов (Кириков, 1952; Potapov, Sale, 2013). Использование этими дендрофильными птицами пространств выше верхней границы леса изучено слабо и, по-видимому, недооценивается (см., например, Островский, 1973; Ménoni, 1991; Нейфельд, Бобрецов, 2002). Тем не менее в данной работе все участки выше этой линии к анализу не привлекались, равно как и пространства ниже границы леса: степные территории Башкирии, Алтая, Тувы и т. д.
Материалы
Географические переменные
Глухарь был или является традиционным объектом охоты, и в настоящее время для многих популяций, включая вымершие, в литературе можно найти оценки веса тела этих птиц (т. е. бесспорные подтверждения их присутствия в конкретных точках). Поскольку популяции с такими оценками размещены по всему ареалу (рис. 1), именно их места расположения являлись объектом представленного ниже анализа. Таксономическая принадлежность птиц указана по Potapov, Sale (2013).
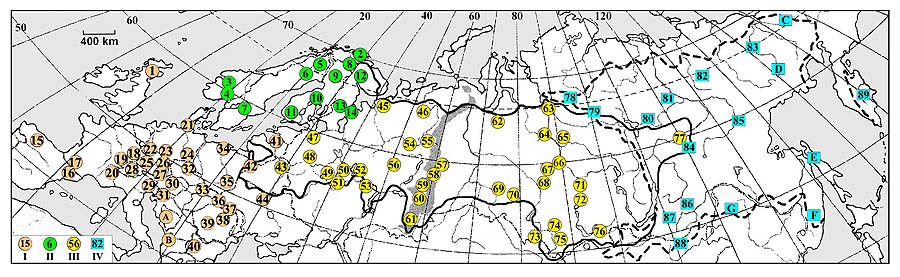
Рис. 1. Места расположения современных и некоторых вымерших (после ~1930-х гг.) популяций глухаря, для которых оценивались характеристики температуры воздуха и количества осадков. А – границы ареала Tetrao urogallus (в западной Европе не показаны из-за сильной фрагментации ареала), Б – ареала T. parvirostris. Группы популяций: I – западноевропейских (T. u. major, T. u. cantabricus, T. u. aquitanicus, T. u. rudolfi), II – фенноскандийских (T. u. urogallus, T. u. lonnbergi, T. u. karelikus), III – всех остальных популяций T. urogallusи, IV – всех популяций T. parvirostris. Цифры в кружках и квадратах – порядковые номера популяций: 1 – Вост. Шотландия (Абердиншир, Абердин, Ангус, Клакманнаншир, Сев. Ланаркшир, Вост. Лотиан, Зап. Лотиан, Мидлотиан, Мрей, Пэрт-энд-Кинросс, Файф, Фолкенр); Норвегия: 2 – Финнмарк, 3 – Бускеруд, Эстфолл, Акерсхус, Оппланд; 4 – Бускеруд, Эстфолл, Эуст-Агдер; Швеция: 5 – Лапландия, 6 – Норботтен, Вестерботтен; 7 – Евлеборг, Стокгольм, Даларна, Уппсала, Вермланд, Эребру, Вестманланд, Сёдерманланд, Эстергётланд, Вестра-Гёталанд, Кальмар; Финляндия: 8 – заповедник Вярриё (сев.-вост. Лапландия), 9 – Лапландия и сев.-вост. Оулу, 10 – юго-запад Оулу, Вааза, Куопио, 11 – все более южные провинции; Россия: 12 – Лапландский заповедник (Мончегорский и Ковдорский р-ны Мурманской обл.); 13 – Республика Карелия (вся); 14 – бассейн р. Илекса (Онежский р-н Архангельской обл.); Испания: 15 – Кантабрийские горы (Астурия, Леон, сев. Паленсия, сев. Заморо, сев. Бургос, Кантабрия, сев.-зап. Галисии); Испания и Андорра: 16 – Наварра, Арагон, Каталония; Андорра; Франция: 17 – Пиренейские горы (Атлантические Пиренеи, Высокие Пиренеи, Верхняя Гаронна, Арьеж, Од, Восточные Пиренеи); 18 – Вогезы (Бельфор, Верхний Рейн, Нижний Рейн, Вогезы, Мёрт и Мозель, Мозель); 19 – Юрские горы (Ду, Юра, Эн); 20 – Альпы (Савойя, Верхняя Савойя); Германия: 21 – Северный Шлезвиг (Шлезвиг-Гольштейн); 22 – Зауэрланд (Северный Рейн-Вестфалия); 23 – Гессен; 24 – Саксония; 25 – Фрайбург, юг Карлсруэ (Баден-Вюртемберг); 26 – Оденвальд, Шпессарт, Рён (Гессен, Рейн-Пфальц, Ниж. Франкония); 27 – Баварские Альпы, Баварский Лес (Бавария); 28 – Швейцария (только альпийская часть: Во, Берн, Вале, Тичино, Граубюнден); 29 – Италия (вост. Альпы: Ломбардия, Трентино-Альто-Адидже, Венето, Фриулия-Венеция-Джулия); 30 – Австрия (Каринтия, Тироль, Зальцбург, Штирия); 31 – Словения (вся страна); 32 – Чехия (только горные территории: Злинский, Моравский, Южноморавский, Краловеградецкий, Карловарский, Либерецкий, Устецкий, Моравскосилезский, Оломуцкий, Пардубицкий, Пльзеньский, Южночешский края); 33 – Словакия (только горные территории: Прешковский, Жилинский, Банскобистрицкий, Тренчинский, Трнавский, Нитранский края); Польша: 34 – Померания (Поморское и Западно-Поморское воеводства); 35 – Люблинское, Свентокшистское воеводства; 36 – Украина (Карпаты: Ивано-Франковская обл.); 37 – Украина / Румыния: Буковина (Черновицкая обл. Украины, жудеца Сучава Румынии); Румыния: 38 – Сев. и Вост. Карпаты (Алба, Вранча, Ковасна, Муреш, Бакзу, Рямц, Харгита, Марамуреш, Сучава, Бистрица-Нэсэуд); 39 – Южные Карпаты (Арджеш, Брашов, Бузэу, Вылча, Горж, Дымбовица, Караш-Северин, Прахова, Сибиу, Хунедоара); 40 – Болгария: Зап. Родопы (Пазарджишская обл.); 41 – Прибалтика: Эстония (вся), Латвия (Видземе, Земгале, Курземе, Латгалия), Литва (Алитусский, Вильнюсский, Каунасский, Клайпедский, Мариямпольский, Паневежский, Таурагский, Тельшяйский, Утенский, Шауляйский р-оны); Белоруссия: 42 – заповедник «Беловежская Пуща» (Каменецкий, Пружанский р-ны Брестской обл., Свислочский р-н Гродненской обл.); 43 – Минская, Витебская обл.; 44 – Украина: Полесье (Волынская, Ровенская, Житомирская, Киевская, Черниговская, Сумская обл.); Россия: 45 – Беломорский п-ов (Приморский, Пинежский, Мезенский р-ны Архангельской обл.); 46 – Север Республики Коми и верховье р. Вымь (Тиманский кряж: Усть-Цилемский, Усинский, Печорский р-ны); 47 – Ленинградская обл.; 48 – Тверская обл.; 49 – Московская обл.; 50 – Ивановская обл.; 51 – Окский заповедник (Спасский, Клепиковский, Касимовский р-ны Рязанской обл.); 52 – Нижегородская обл. (все левобережье р. Волги); 53 – Республика Мордовия (Темниковский, Ичалковский, Большеигнатовский, Ардатовский, Атяшевский, Чамзинский, Дубёнский, Большеберезниковский р-ны); 54 – юг Республики Коми (Сыктывдинский, Корткеросский р-ны); 55 – Печоро-Илычский заповедник (Троицко-Печорский р-н Республики Коми); 56 – Кировская обл. (Слободской р-н); 57 – север Свердловской обл. (Нижнетуринский, Верхотурский, север Алапаевского, Туринский, Тавдинский и все более северные р-ны); 58 – юг Свердловской обл. (все остальные южные р-ны); 59 – Южно-Уральский заповедник (Белорецкий р-н Башкирии, Катав-Ивановский р-н Челябинской обл.); 60 – Курагчинский, Архангельский р-ны Республики Башкирия; 61 – Башкирский заповедник и юг Башкирии (Бурзянский, Абзелиловский, Белорецкий, Кугарчинский, Зилаирский, Зианчуринский р-ны Республики Башкирия); 62 – Обско-Тазовское междуречье (Шурышкарский, Приуральский, Надымский, Пуровский р-ны Ямало-Ненецкого АО); 63 – низовья р. Курейка (Туруханский, Илимпийский р-ны, адм. р-н г. Дудинка, Красноярского края); 64 – Туруханский р-н Красноярского края; 65 – Туруханский, Эвенкийский р-ны Красноярский края; 66 – бассейн р. Сым (Енисейский р-н, Красноярский края); 67 – бассейн р. Лисица (Верхнекетский р-н Томской обл.); 68 – Бараба и Кулунда (Парбигский, Бакчарский р-ны Томской обл.); 69 – Кыштовский р-н Новосибирской обл.; 70 – Усть-Тарский, Венгеровыский, Куйбышевский, Убинский, Каргатский, Чулымский и все более северные и восточные р-ны Новосибирской обл.; 71 – юг Енисейской равнины (Ермаковский, Каратузский, Минусинский, Курагинский, Краснотуранский, Идринский, Партизанский, Саянский, Ирбейский, Новоселовский, Балахтинский, Березовский, Манский р-ны Красноярского края); 72 – окрестности г. Красноярск; 73 – Казахстан: Глубоковский, Лениногорский, Зыряновский, Катон-Карагайский, Уланский, Кокпектинский, Куршимский р-ны Восточно-Казахстанской обл.; Россия: 74 – Бийский р-н Алтайского края; 75 – Алтайский заповедник (Турчакский, Улаганский р-ны Республики Алтай); 76 – Пий-Хемский, Каа-Хемский, Кызылский, Тоджинский кожууны Республики Тува; 77 – среднее течение р. Вилюй (Сунтарский р-н Якутии); 78 – плато Путорана (Таймырский, Эвенкийский р-ны Красноярского края); 79 – бассейн р. Кочечум (Эвенкийский р-н Красноярского края); 80 – среднее течение р. Вилюй (Мирнинский/Сунтарский р-ны Якутии); 81 – бассейн р. Кенкэмэ (Горный р-н Якутии); 82 – Верхоянский р-н Якутии; 83 – среднее течение р. Колыма (Нижнеколымской р-н Якутии); 84 – бассейн р. Токко (Олёкминский р-н Якутии); 85 – низовья р. Мая (Усть-Майский р-н Якутии); 86 – Забайкальский край (весь); 87 – Баунтовский, Еравнинский р-ны Бурятии; 88 – Монголия: Кэнтий (аймак Хэнтий); 89 – Россия: Карагинский, Усть-Камчатский, Мильковский, Елизовский, Усть-Большерецкий р-ны Камчатской обл. Добавленные точки (для них климатические модели не строились, пояснения см. текст). A – Босния и Герцеговина: Чуховицы (Герцеговино-Неретвенский кантон); B – Албания: Аррт (обл. Дибра); Россия: C – Билибинский р-н Чукотки; D – Среднеканский р-н Магаданской обл.; E – Тымовский р-н Сахалинской обл.; F – Кировский р-н Приморского края; G – Китай: Джагдачи (Внутренняя Монголия)
Fig. 1. Locations of present and some extinct (after ~1930 years) populations of capercaillie for which the indexes of air temperature and precipitation were evaluated. А – boundaries of the geographical range of Tetrao urogallus (not indicated in western Europe due to high fragmentation), Б – the ones of T.parvirostris. The population groups: I – from southern and central parts of Western Europe (T. u. major, T. u. cantabricus, T. u. aquitanicus, T. u. rudolfi), II – fennoscandian (T. u. urogallus, T. u. lonnbergi, T. u. karelikus), III – all other of T.urogallus, IV – all populations of T.parvirostris. Figures in circles and squares denote the serial numbers of populations: 1 – East Scotland (Aberdeenshire, Aberdeen, Angus, Clackmannanshire, North Lanarkshire, East Lothian, West Lothian, Midlothian, Mray, Prert-and-Kinross, Fife, Falkenre); Norway: 2 – Finnmark, 3 – Buskerud, Ostfold, Akershus, Oppland; 4 – Buskerud, Ostfold, East-Agder; Sweden: 5 – Lapland, 6 – Norbotten, Westerbotten; 7 – Gävleborg, Stockholm, Dalarna, Uppsala, Vermland, Örebro, Vestmannland, Södermanland, Ostergotlands, Vestra Gotaland, Kalmar; Finland: 8 – Reserve Värriö (north-east Lapland), 9 – Lapland and north-east. Oulu, 10 – south-west of Oulu, Vaasa, Kuopio, 11 – increasingly southern provinces; Russia: 12 – Lapland Reserve (Monchegorsk and Kovdor districts of Murmansk region); 13 – Republic of Karelia (whole); 14 – the Ileksa river basin (Onega district, Arkhangelsk region); Spain: 15 – Cantabrian Mountains (Asturias, León, northern Palencia, northern Zamoro, northern Burgos, Cantabria, northwest of Galicia); Spain and Andorra: 16 – Navarre, Aragon, Catalonia; Andorra; France: 17 – Pyrenees (Atlantic Pyrenees, High Pyrenees, Upper Garonne, Ariège, Aude, East Pyrenees); 18 – Vosges (Belfort, Upper Rhine, Lower Rhine, Vosges, Meurthe and Moselle, Moselle); 19 – Jura Mountains (Du, Jura, En); 20 – Alps (Savoy, Haute-Savoy); Germany: 21 – Northern Schleswig (Schleswig-Holstein); 22 – Sauerland (North Rhine-Westphalia); 23 – Hessen; 24 – Saxony; 25 – Baden-Württemberg (Freiburg, south of Karlsruhe); 26 – Odenwald, Spessart, Rien: Hesse, Rhine-Pfalz, Lower Franconia; 27 – Bavaria (Bavarian Alps, Bavarian Forest); 28 – Switzerland (only the alpine part: Vau, Berne, Valais, Ticino, Graubunden); 29 – Italy (eastern Alps: Lombardy, Trentino-Alto-Adige, Veneto, Friuli-Venezia-Giulia); 30 – Austria (Carinthia, Tyrol, Salzburg, Styria); 31 – Slovenia (whole country); 32 – Czech Republic (only mountain areas: Zlinsky, Moravian, South Moravian, Hradec Králové, Karlovy Vary, Liberec, Ustecky, Moravian-Silesian, Olomouc, Pardubice, Pilsen, South Bohemian districts); 33 – Slovakia (only mountain areas: Preshkovsky, Zhilinsky, Bansko Bistritsky, Trencinsky, Trnavsky, Nitran districts); Poland: 34 – Pomerania (Pomerania and West Pomerania voivodeships); 35 – Lubelskie, Сwiętokrzyskie voivodeships; 36 – Ukraine (Carpathians: Ivano-Frankivsk region); 37 – Ukraine / Romania: Bukovina (Chernivtsi region of Ukraine, Suceava judet of Romania); Romania: 38 – Northern and Eastern Carpathians (Alba, Vrancea, Covasna, Mures, Baczu, Ramatz, Hargita, Maramures, Suceava, Bistrita-Nasoud); 39 – Southern Carpathians (Arges, Brasov, Buzau, Valcea, Gorzh, Dymbovitsa, Karash Severin, Prahova, Sibiu, Hunedoara); 40 – Bulagaria: Western Rhodopes (Pazardzhis region); 41 – The Baltic States: Estonia (whole), Latvia (Vidzeme, Zemgale, Kurzeme, Latgale), Lithuania (Alytus, Vilnius, Kaunas, Klaipeda, Marijampolė, Panevėžys, Tauragi, Telšiai, Utena, Siauliai districts); Belarus: 42 – Reserve Belovezhskaya Pushcha (Kamenetsky, Pruzhany district of Brest region., Svisloch district of Grodno region); 43 – Minsk, Vitebsk regions; 44 – Ukraine: Polessye (Volyn, Rivne, Zhytomyr, Kiev, Chernigov, Sumy regions); Russia: 45 –White Sea Peninsula (Primorsky, Pinezhsky, Mezensky districts of Arkhangelsk region); 46 – North of Komi Republic and headwaters of the Vym river (Timan Mountains: Ust-Tsilemsky, Usinsk, Pechora districts); 47 – Leningrad region; 48 – Tver region; 49 – Moscow region; 50 – Ivanovo region; 51 – Oka Reserve (Spassky, Klepikovsky, Kasimovsky districts of Ryazan region); 52 – Nizhny Novgorod region (all left bank of the Volga river); 53 – Republic of Mordovia (Temnikovsky, Ichalkovsky, Bolsheignatovsky, Ardatov, Atiashevsky, Chamzinsky, Dubyonsky, Bolshebereznikovsky districts); 54 – south of Komi Republic (Syktyvdinsky, Kortkerossky districts); 55 – Pechora-Ilychsky reserve (Troitsko-Pechorsky district, Komi Republic); 56 – Kirov region (Slobodskoy district); 57 – north of Sverdlovsk region. (Nizhneturinsky, Verkhotursky, north of Alapaevsky, Turin, Tavdin and increasingly northern districts); 58 – south of Sverdlovsk region (all other southern districts); 59 – South Ural Reserve (Beloretsky district of Bashkiria, Katav-Ivanovskiy district of Chelyabinsk region); 60 – Kuragchinsky, Arkhangelsk Regions of Bashkiria; 61 – Bashkir reserve and south of Bashkiria (Burzyan, Abzelilovsky, Beloretsky, Kugarchinsky, Zilair, Zianchurinsky districts of Bashkiria); 62 – Ob-Taz watershed (Shuryshkarskiy, Priuralsky, Nadymsky, Purovsky districts of Yamal-Nenets Autonomous Okrug); 63 – lower reaches of the river Kureika (Turukhansk, Ilimpiysky, Dudinka districts of Krasnoyarsk Territory); 64 – Turukhansk district of Krasnoyarsk Territory; 65 – Turukhansky, Evenkiysky districts of Krasnoyarsk Territory; 66 – basin of the river Sym (Yenisei district of Krasnoyarsk Territory); 67 – basin of the river Lisitsa (Verkhneketskiy district of Tomsk region.); 68 – Baraba and Kulunda (Parbigsky, Bakcharsky districts, Tomsk region); 69 – Kyshtovsky district of Novosibirsk region; 70 – Ust-Tarsky, Wengerovo, Kuibyshev, Ubinsky, Kargatsky, Chulymsky and all more northern and eastern districts of Novosibirsk region; 71 – south of Yenisei plain (Ermakovsky, Karatuzsky, Minusinsk, Kuraginsky, Krasnoturansky, Idrinsky, Partizansky, Sayansky, Irbeisky, Novoselovsky, Balakhtinsky, Berezovsky, Mansky districts of Krasnoyarsk Territory); 72 – outskirts of Krasnoyarsk town; 73 – Kazakhstan: Glubokovsky, Leninogorsk, Zyryanovsky, Katon-Karagai, Ulan, Kokpekty, Kurishim districts of East Kazakhstan region; Russia: 74 – Biysk district of Altai Territory; 75 – Altai Reserve (Turchaksky, Ulagansky districts, Republic Altai); 76 – Piy-Khem, Kaa-Khem, Kyzyl, Todzhinsky kojuuns of Republic Tyva; 77 – middle flow of the Vilyui River (Suntarsky district of Yakutia); 78 – Plateau Putorana (Taimyrsky, Evenkiysky districts of Krasnoyarsk Territory); 79 – the Kochechum river basin (Evenkiysky district of Krasnoyarsk Territory); 80 – middle flow of the Vilyui River (Mirninsky / Suntarsky districts of Yakutia); 81 – the Kenkeme river basin (Gorniy district of Yakutia); 82 – Verkhoyansk district of Yakutia; 83 – middle flow of the Kolyma River (Nizhnekolymsky district of Yakutia); 84 – the Tokko river basin (Olekminsky district of Yakutia); 85 – lower flow of the Maya River (Ust-Maysky district of Yakutia); 86 – Zabaikalsky Territory (whole); 87 – Bauntovsky, Eravninsky districts of Buryatia; 88 – Mongolia: Cantiy (aimak Hentiy); 89 – Russia: Karaginsky, Ust-Kamchatsky, Milkovsky, Elizovsky, Ust-Bolsheretsky districts of Kamchatka region. Added points (climatic models were not developed, see text). A – Bosnia Herzegovina: Chukhovitsy (Herzegovina-Neretva canton); B – Albania: Arrt (Dibra region); Russia: C – Bilibino district of Chukotka; D – Srednekansky district of Magadan region; E – Tymovskiy district of Sakhalin region; F – Kirov district of Primorsky Territory; G – China: Jagdachi (Inner Mongolia)
Координаты популяций (широта, долгота) взяты из литературы или определены по обозначенным в ней ориентирам, с помощью которых выяснялся пространственный контур популяционного ареала и его географический центр. Долгота и широта центра считывались с изображений программы Google Earth.
Во многих случаях в литературе указаны не места, занимаемые популяциями, а регионы, в которых они располагались. В таком случае в популяционный ареал включалась вся занятая глухарем часть региона. Например, вся Карелия, но только населенный глухарем север Тувы. В отдельных случаях разные источники характеризовали близкие популяционные арены, и мы рассматривали их как единые территории. Например, все указания на Ливонию, Курляндию, Лифляндию, Эстонию, «Балтийское побережье» и т. п. объединены в регион, охватывающий все три современные прибалтийские государства (ниже именуется Прибалтикой). При указаниях на различные, хотя и перекрывающиеся пространства, например, регионы южной Норвегии (см. рис. 1, № 3 и 4) или Красноярского края (см. рис. 1, № 64 и 65), мы рассматривали их как разные популяции. В одном случае неопределенными оказались и границы всего региона – Шведской Лапландии. Ниже она рассматривается как вся лежащая к северу от полярного круга часть лена Норрботтен. При указании на расположения популяций в бассейнах рек (р. Сым, р. Токко и т. д.) к ареалу причислялся весь или почти весь бассейн. При большей конкретизации («средняя Колыма», «низовья р. Маи» и т. п.) в ареал включалась соответствующая треть бассейна. Вероятные смещения популяционных центров (максимум до 2.0–2.5о, обычно гораздо меньше) из-за этих неопределенностей в рамках всего ареала T. urogallus и даже T. parvirostris представляются несущественными.
Высоты местности, занимаемые каждой популяцией, охарактеризованы единственными показателями: для каждой популяции – неким средним уровнем. Он оценивался по литературным источникам или, если такой информации найти не удавалось, по множествам высотных оценок, считанных с изображений программы Google Earth (подробнее см. ниже). Количество оценок (точек) для каждого вмещающего популяцию пространства менялось в зависимости от его размеров и характера рельефа: на равнинах 50–380 точек, в горах – 140–700. После удаления точек, лежавших за границами леса, к расчетам принято почти 12.6 тыс. оценок (точек).
Климатические показатели
Разработка моделей
Единый (и единственный) информационный источник для разработки моделей, хотя и не снимает проблем осцилляций климата, однако существенно ограничивает ее влияние на конкретные оценки. Поэтому в качестве основного информационного ресурса использован единый и единственный массив данных: материалы климатического сайта (Климатические…, 2017). На его страницах представлены: 1) географические названия конкретных точек, 2) их абсолютные высоты, 3) среднегодовые температуры воздуха и 4) годовые суммы атмосферных осадков. Эти две последние величины получены по результатам моделирования (методика не указана) данных о погоде «с тысяч метеостанций» за 1982–2012 гг. (Источник данных сайта). Географические координаты точек на данном сайте отсутствуют, поэтому каждая взятая с него точка отыскивалась по ее названию на изображениях программ Google Earth, Яндекс-Карты или OpenStreeMap (2017), откуда и считывались значения широты и долготы.
Значения и температуры, и осадков (непременно оба показателя) брались для каждой принятой к анализу точки. Количество точек, использованных для построения конкретных моделей, менялось в зависимости от сложности и площади региона: от 48 до 376 шт. Их суммарное количество – почти 5.6 тыс. шт. Всего для оценки климатических параметров по географическому положению 89 популяций глухаря (см. рис. 1) разработано 69 моделей температуры и 67 моделей осадков.
Для более полного представления обо всем родовом ареале глухаря в анализ добавлено несколько периферийных точек (с буквенными символами, см. рис. 1). Модели для них не разрабатывались: каждая точка представлена многолетними данными с единственной метеостанции (Климатические…, 2017). Абсолютные высоты каждой из этих станций, возможно, не адекватны средним высотам региональных мест обитания глухаря, но, безусловно, лежат в их пределах.
Границы термогумидного пространства Северной Евразии
Хотя распространение глухаря в основном определяется лесной средой (см. выше), мы попытались уточнить общие пределы доступного ему термогумидного пространства, т. е. уяснить границы этого пространства в пределах всей суши Северной Евразии. Господство степей и пустынь (жарких или арктических) в аридном секторе этого пространства (количество осадков < 150–180 мм/год) поднимало данный вопрос только в отношении гумидного сектора. Для каждого одного градусного интервала температуры с сайтов (Климатические…, 2017; Clima…, 2017) отбирались точки с предельно высоким количеством осадков. Выборка (n > 8 тыс. точек, к анализу принята 51) проведена только по районам с большой вероятностью сочетания низких температур и повышенного количества осадков: высокогорья Европы (Пиренеи, Альпы, Карпаты) и побережья холодных морей (от Англии и Норвегии до Чукотки и далее на юг до Приморья). Для отображения максимального количества осадков при данных температурах воздуха в пределах суши Северной Евразии использовано лишь 18 точек.
Проверка моделей
Разные источники могут давать для одних и тех же точек различные значения температуры и осадков, что возможно из-за привлечения их составителями несовпадающих хронологических отрезков, материалов с разных метеостанций (несовпадающие высоты) одного и того же населенного пункта и т. д. Ниже эти расхождения рассматриваются как информационный шум. Для его измерения мы сопоставили данные для одних и тех же точек (n = 27), взятых из двух разных источников (Термограф…, 2017; Clima…, 2017).
Эти же два сайта использованы для проверки адекватности моделей. К ним добавлены полученные от коллег (см. Благодарности) данные с семи метеостанций Русской равнины. Всего для проверки температурных моделей использовано 100 географических точек, для моделей осадков – 97. Каждая из них характеризовалась значениями температуры, осадков, широты, долготы и высоты. Все эти оценки не привлекались к разработке моделей и рассматривались как ожидаемые, или контрольные.
Из 69 температурных и 67 моделей осадков тестировалось лишь 49 наиболее проблемных: горные и/или крупные по площади регионы. Модели, полученные для небольших равнинных регионов (Ленинградская, Тверская, Московская обл.…), не проверялись.
Методы
Высоты местности в популяционных центрах
Эти показатели оценивались двумя методами. Первый – только для средне- и высокогорных районов. Из публикаций выбирались показатели высотного размещения глухаря в конкретных горных системах: обычно указываются предельные или предпочитаемые уровни. Высота расположения всей популяции оценивалась как среднее арифметическое из двух значений того или иного интервала. Сообщения о вертикальных миграциях птиц, гендерные предпочтения высотных ярусов и т. п. во внимание, к сожалению, не принимались. При наличии нескольких публикаций для одной и той же популяции с несовпадавшими указаниями на занимаемые высоты использован интервал перекрывавшихся значений. Если указаний на высотное размещение глухаря в горной местности найти не удавалось, использовались данные о высотном интервале лесной растительности. Он устанавливался по общегеографическим (Добрынин, 1948; Мильков, Гвоздецкий, 1976; Гвоздецкий, Михайлов, 1978) или региональным источникам (Чернов, 1964; Васильев и др., 1980; Романов, 2004; Горичев, 2008 и др.), по электронным справочникам и личным сообщениям коллег.
Второй метод использован при работе с равнинами и низкогорьями. В программе Google Earth на изображение всего регионального ареала глухаря наносились линии географической сетки и с нескольких точек их пересечения считывались значения высот: точки включались в выборки через равномерные интервалы. Средняя арифметическая величина из этих оценок характеризовала абсолютную высоту местности для всей конкретной популяции. Этот же метод использован в работе с горными районами, если для них был получен только высотный интервал лесной растительности. Фактически таким способом оценивался средний уровень абсолютных высот, занимаемых глухарем в лесном поясе конкретной горной системы.
При считывании высот использованы изображения местности в масштабе 1:400 000–1:700 000 для равнин и 1:100 000–1:500 000 для горных районов.
Климатические модели
Смысл создания этих моделей определялся необходимостью получить региональные зависимости (формулы) температуры и осадков от трех пространственных показателей (широты, долготы и высоты) только внутри популяционных арен глухаря или для групп некоторых из них. По этим формулам рассчитаны значения температуры и осадков для центра каждой популяции, показанной на рис. 1.
Работа с материалом сводилась к отбору некоего множества пространственно взаимоудаленных точек, достаточного для выявления региональных закономерностей изменения температуры и осадков в зависимости от расположения оценок, принимаемых к анализу. Поскольку пространственное расположение точек с оценками температуры и осадков оказалось крайне неравномерным (см. Введение), особое внимание уделялось поиску точек с предельными значениями (единицы из сотен просматривавшихся нами). Такие точки в дополнение к множеству других давали необходимые «опоры» для аппроксимирующих кривых наших моделей.
Все региональные модели построены с помощью множественного линейного регрессионного анализа (пакет Statistica 6.0), для чего в качестве зависимых переменных использованы ряды климатических показателей, в качестве независимых (объясняющих) переменных – ряды пространственных оценок:
Y = In + b · La + c · Lo + d · Al,
где Y – значение температуры или осадков, In – свободный член, La – широта, Lo – долгота, Al – высота местности; b, c, d – расчетные коэффициенты для широты, долготы и высоты. Попытки нелинейного моделирования дали неудовлетворительные результаты, и мы к ним больше не возвращались.
При хорошей аппроксимации зависимых рядов только одной или двумя переменными (например, только значениями Al или только Lo и Al) к работе принимались соответственно одно- или двухмерные модели, но в большинстве случаев они базировались на всех трех переменных. Если совместное влияние двух или трех переменных было и достоверным, и сильным, но воздействие одной из них оказывалось недостоверным, в модель включались все две или три переменные, при условии, что исключение недостоверных влияний не приводило к увеличению силы связей (контроль по multiple R2adj). При получении альтернативных вариантов предпочтения отдавались моделям с наиболее сильными связями.
Модели температуры и осадков разработаны для каждого из этих параметров раздельно, но по одним и тем же региональным данным за исключением редких случаев, когда первоначальный анализ не давал достоверных связей или их сила оказывалась слабой. В таком случае мы либо увеличивали, либо уменьшали масштаб исследования. Так, достоверных связей для Кантабрийских гор (см. рис. 1, № 15) найти не удалось ни для температуры, ни для осадков. Однако они выявлены для территорий внутри этого региона: для провинций Астурия, Леон (вместе с соседними участками Паленсии, Самора и Бургоса), Кантабрия и Луго. Модели построены для каждой из четырех провинций и для каждой рассчитаны значения температуры и осадков; весь регион (№ 15) в целом охарактеризован средней величиной из четырех субрегиональных оценок. Однако чаще приходилось увеличивать площадь региона. Для точек № 8–11 (см. рис. 1) использованы данные по всей Финляндии в целом, для № 14 и 45 – по всей Архангельской обл., для точек № 46, 54 и 55 – по всей территории Коми, для точек № 42 и 43 – по всей Белоруссии, для № 57 и 58 – по всей Свердловской обл., для № 63–66, 71, 72, 78 и 79 – по всему Красноярскому краю, для № 74 и 80–85 – по всей Якутии.
Для некоторых регионов использован «смешанный» подход, например, Башкирия (№ 59–61) разделена на три части: запад, центр и восток. Для точки № 59 взяты данные только с севера центральной Башкирии (с добавлением соседних участков Челябинской обл.), для точки № 60 – только с ее западной части, для точки № 61 – и с востока, и из центра республики. Температурная модель для Мордовии построена только по данным с ее собственной территории, но для модели осадков дополнительно привлечены материалы из соседней Рязанской области.
В процессе проверки моделей выяснялось, выходят ли контрольные оценки за крайние значения вариационных рядов, использованных для построения этих моделей. Например, ряд, взятый для моделирования количества осадков в Баварии (n = 82) на высотах в 318–1043 м, дает вариацию в пределах 634–1337 мм. Но одна из трех контрольных точек этого региона (Цугшпитце, абс. высота 2960 м), выходя за пределы высотного ряда, показывает количество осадков в 2004 мм, что превышает максимальное значение ряда осадков на 961 мм (отклонение на 92 %). При невозможности объяснить такие отклонения модель рассматривалась как некорректная.
Результаты
Качество климатических моделей
Все модели температуры показали адекватность исходным данным (p < 0.001) и сильную зависимость от мест расположения (для 90 % моделей r > 0.9, для остальных r > 0.7). Большинство (83 %) моделей осадков также демонстрировали высокую зависимость (r > 0.7), связи средней силы (r = 0.40–0.69) получены для 14 % моделей и лишь две (3 %) показали слабые зависимости: Италия (№ 29, r = 0.25) и Австрия (№ 30, r = 0.22). Почти для всех моделей характерен высокий уровень адекватности (p < 0.001) за исключением четырех: Италии (p = 0.001), Австрии (p = 0.005), Словении (№ 31: p = 0.011) и Бурятии (№ 87: p = 0.012). Как и предполагалось (см. Введение), на региональном уровне широта, долгота и высота местности жестче связаны с температурой, чем с режимом осадков.
Для Северного Шлезвига (№ 21) силу связи в температурной модели удалось существенно повысить при его объединении с Померанией (№ 34). Но достоверной зависимости осадков от расположения точек в Северном Шлезвиге не выявлено. Для него рассчитано среднее (n = 12) из количества осадков, выпадающих в интервале высот 0–32 м, при средней высоте региона в 16 м. Для Забайкальского края (№ 86) также не получено достоверной связи по осадкам, и этот показатель тоже рассчитан как среднее (n = 12) для интервала высот в 900–1000 м, при средней высоте лесного пояса в 948 м.
Изучение информационного шума показало ожидаемое несовпадение значений температуры и осадков в разных источниках. Наибольшие расхождения дали оценки температуры: максимум – до 60 % от сравниваемых величин. Но только для двух точек из 27 (8 % парных сравнений) величины расхождений превышали 20 %. По осадкам максимальные расхождения любых парных сравнений достигали лишь 20 %. Следовательно, основной уровень шума расположен в пределах ± 20 % от абсолютных значений, содержащихся в информационных источниках, и при оценке корректности моделей мы пренебрегали такими расхождениями.
Проверка температурных моделей показала выход только четырех контрольных оценок за пределы эмпирических вариационных рядов (модели по французским Альпам, Саксонии, Вестфалии и Болгарии). Одна из контрольных оценок (по Болгарии) получена для абсолютной высоты, которая не попадает в интервал высотного ряда, использованного для построения данной модели, и, следовательно, не может быть аргументом ее некорректности. Отклонения трех остальных контрольных оценок от крайних значений вариационного рада составляют ±0.2–0.5 оС, что не превышает 5 % крайних величин, использованных для моделирования. Следовательно, все реальные отклонения укладываются в рамки информационного шума, и все модели температуры можно рассматривать как корректные.
За пределы вариационных рядов наших моделей осадков выходит 13 контрольных оценок (к 12 моделям). Три из них характеризуют точки, лежащие за пределами абсолютных высот, использованных для разработки моделей (по южной Норвегии, Ивановской обл. и особенно по Баварии, см. выше). Десять остальных контрольных оценок выходят за пределы крайних значений вариационных рядов на 6–177 мм, т. е. на 1–17 %. Такие отклонения не превышают уровня шума, и, следовательно, все модели могут также рассматриваться как корректные.
Два других подхода к функциональной проверке моделей – по отклонениям проверяемых оценок от кривой, аппроксимирующей контрольные значения (с и без учета информационного шума), – также реализованы нами и также показали порректность моделей. В данной работе их результаты не представлены (слишком громоздкие).
Температура и осдки в пределах родового ареала
В пределах всего родового ареала глухаря обе изучаемые переменные – температура и осадки – показали достоверные зависимости от широты и долготы, а осадки еще и от высоты местности (табл. 1). Кроме того, для наших выборок (популяции на рис. 1: n = 89) характерны достоверные связи всех географических переменных: широты с долготой (r = 0.39 p < 0.001), широты с высотой (r = –0.68 p < 0.001) и долготы с высотой (r = –0.21 p = 0.05). По-видимому, именно такие несильные, латентные связи создают существенные проблемы при реализации традиционного анализа адаптивных реакций птиц по осям пространственных переменных (см. Введение), от которого мы отказались. Визуальный анализ взаимного расположения популяций в осях температуры и осадков индифферентен к этим связям.
Таблица 1. Зависимость температуры и осадков от географических переменных в родовом ареале глухаря
Table 1. Dependence of temperature and precipitation on geographic variables within the genus range of capercaillie
| Температура / Temperature | Осадки / Precipitation | |||
| r | p | r | p | |
| Широта / Latitude | – 0.62 | < 0.001 | – 0.59 | < 0.001 |
| Долгота / Longitude | – 0.86 | < 0.001 | – 0.67 | < 0.001 |
| Высота / Altitude | 0.17 | 0.113 | 0.55 | < 0.001 |
Все обсуждаемое термогумидное пространство (рис. 2) разбито на четыре части, разделительные линии проведены через центры интервалов, образованных крайними значениями региональных климатических оценок. Для всех принятых к анализу популяций глухаря эти крайние значения составляют: температуры – от –13.4 до +8.5 оС, осадков – от 196 до 1426 мм. В верхнюю правую четверть попадают группировки, живущие в теплых и влажных условиях, в нижнюю правую – в теплых и сухих, в нижнюю левую – в холодных и сухих; пространство в верхней левой четверти (холодные и влажные условия) на суше Северной Евразии, по-видимому, практически отсутствует. Хотя в верхнем правом секторе это пространство выходит далеко за указанные нами пределы, простираясь (по мере нарастания температур) вдоль шкалы осадков от 1200–1300 (отдельные точки в Норвегии и на о. Сахалин) до 2000–3100 мм/год (Норвегия, Швейцария, Бавария, Австрия, Италия). Многоступенчатый характер всей работы (см. Введение) и невысокое качество описаний популяционных ареалов в некоторых первоисточниках позволяет рассматривать конечные результаты моделирования лишь как приближение к реальности. Поэтому конечные оценки не сопровождаются индексами вариации (см. рис. 2), которые могли вызвать иллюзию высокой точности, чего нет в реальности.

Рис. 2. Расположение мест обитания некоторых популяций глухаря в осях температуры воздуха и количества атмосферных осадков (расчетные значения). Группы популяций: I – западноевропейских (T. u. major, T. u. cantabricus, T. u. aquitanicus, T. u. rudolfi), II – фенноскандийских (T. u. urogallus, T. u. lonnbergi, T. u. karelikus), III – всех остальных популяций T. urogallus. IV – термогумидное пространство, которое, по-видимому, отсутствует на суше Северной Евразии. Пунктир – граница группы I (проведена вручную). Номера и символы возле точек те же, что на рис. 1
Fig. 2. Location of habitats of some capercaillie populations in axes of air temperature (X) and precipitation (Y) (calculated values). The population groups: I – in southern and central parts of Western Europe (T. u. major, T. u. cantabricus, T. u. aquitanicus, T. u. rudolfi), II – fennoscandian (T. u. urogallus, T. u. lonnbergi, T. u. karelikus), III – all the rest of Tetrao urogallus, IV – thermohumid space seems to be absent on land of Northern Eurasia. Dotted line is the boundary of group I. The signs next to circles and squares are the same as in figure 1
Область перекрывания термогумидного пространства обыкновенного и каменного глухарей представляется существенной, о чем говорят крайние видовые значения: по оси температуры от –8.5 до +3.0 оС. И такой размах занимает 52–53 % всего родового ареала по оси температуры. Перекрывание по оси осадков – от 260 до 839 мм (47 %). В зоне перекрывания интересно размещение отдельных популяций. В область холодного и сухого климата, занятую преимущественно каменным глухарем, попадает, например, точка № 8 (см. рис. 2) – северо-восток финской Лапландии, – лежащая всего в ~250 км от морского побережья. Температурная оценка для популяции T. parvirostris из Приморского края (точка F, см. рис. 2) лежит недалеко от оценок, полученных для группировок T. urogallus из Ленинградской области (№ 47) и даже с юга Норвегии (№ 4). Камчатская популяция (№ 89) обитает в условиях примерно такого же количества осадков, что и группировки T. urogallus из украинских (№ 36) и румынских Карпат (№ 38) и даже из Германии (№ 26). Близким оказывается количество осадков и для таких удаленных регионов, как, с одной стороны, Якутия (№ 80, 81, 85), Южная Сибирь (№ 86, 87), Монголия (№ 88), так и, с другой стороны, …шведская Лапландия (№ 5).
Несмотря на такой разброс оценок, хорошо просматриваются и межвидовые различия: T. urogallus населяет в основном регионы более теплого климата, как сухого, так и влажного, T. parvirostris – более холодного и, главным образом, сухого. За самый низкий (сухой) предел ареала T. urogallus (№ 75, см. рис. 2) выходят всего две популяции T. parvirostris: колымская (№ 83) и чукотская (точка С). Заселенное этим видом пространство с количеством осадков > 600 мм, по-видимому, ограниченно. В наш анализ попало всего три точки: Камчатка (839 мм: № 89), Приморье (702 мм: точка F) и Сахалин (662 мм: точка Е). Похоже, что по оси осадков ареал каменного глухаря занимает в основном сухую часть ареала T. urogallus, почти полностью умещаясь в пределы последнего и занимая его меньшую часть.
Полученные оценки позволяют частично разграничить даже ареалы подвидов. В пределах термогумидного пространства T. urogallus территориально единую, целостную группу образует несколько западноевропейских подвидов: T. u. major (23 точки: № 1, 18–35, 37, 41, 42, 44, см. рис. 2), T. u. cantabricus (№ 15), T. u. aquitanicus (16, 17) и T. u. rudolfi (36, 38, 39, 40). Они вполне ожидаемо занимают самую теплую и влажную часть всего ареала рода Tetrao и на рис. 2 отделены от остальных подвидов точечным пунктиром. Самую теплую часть ареала этой группы используют четыре немецкие популяции (№ 21, 23, 25, 26, см. рис. 2), самую холодную – группировки из Румынии (39), Австрии (30), Словакии (32) и Чехии (33). В самую сырую часть – с максимальным количеством осадков для всего родового ареала в целом – попадают популяции из Юры (№ 19), французских Альп (20), Швейцарии (28) и Словении (31). Термогумидная область западноевропейской группы, по-видимому, почти не перекрывается с аналогичными областями других подвидов (см. рис. 2). Напротив, объединенное термогумидное пространство трех фенноскандийских подвидов (T. u. urogallus, T. u. lonnbergi, T. u. karelikus, 13 точек: № 2–14, см. рис. 2) сильно перекрывается с пространством не только восточно-европейских, уральских и западносибирских подвидов (№ 43, 45–62, 67–70), но и приближается к центрально-сибирским условиям (окрестности Красноярска: № 72). Оконтурить собственное термогумидное пространство только феноскандийской группы (без включения других подвидов) непрерывной и/или слабо извилистой линией не удается.
В ареале каменного глухаря явно обособленное положение занимает камчатская популяция (T. p. camtschaticus, № 89, рис. 2), которая по шкале осадков попадает в область распространения западноевропейских подвидов T. urogallus и лежит в более теплой зоне, чем, например, все фенноскандийские популяции за исключением одной из норвежских (№ 3). T. p. stegmanni (точки 86–88, G), по-видимому, располагается на сухой периферии теплой области видового ареала. Положение подвида T. p. parvirostris не выяснено, т. к. для населенных им Хабаровского и Приморского краев региональные модели не разрабатывались.
Для всего родового ареала в целом прослежена прямая, но не линейная связь температуры (x) и осадков (y): rs = 0.77 p < 0.001, т. е., чем теплее места обитания, тем больше в них выпадает осадков. И общее размещение глухаря в термогумидном пространстве удается охарактеризовать зависимостью осадков от температуры: у = 605.78 + (32.94х) + (0.70х2). Детализация общей картины показывает, что нелинейность этой связи возникает из двух противоположных трендов. Для термогумидного ареала западноевропейской группы подвидов характерна линейная обратная связь: чем теплее, тем меньше осадков (рис. 3); удаление из расчета равнинных популяций приводит к потере ее достоверности: r = – 0.07 p = 0.78. Пространства всех остальных подвидов демонстрируют прямые линейные связи между теплом и осадками: чем теплее, тем больше осадков. Связи эти не сильные, но достоверные. Для фенноскандийской группы: r = 0.61 p = 0.03; для всех других подвидов T. urogallus, обитающих восточнее Бреста, Санкт-Петербурга и Онеги: r = 0.48 p = 0.004; для всего ареала T. parvirostris: r = 0.70 p = 0.02. Вполне ожидаемо, что и вся территория в целом, занятая не западноевропейскими подвидами обыкновенного и всеми подвидами каменного глухаря, также показала прямую связь между теплом и осадками: r = 0.64 p < 0.001. Следовательно, закономерность пространственных изменений двух важнейших климатических показателей (их интегральный градиент) в ареале западноевропейских подвидов кардинальным образом отличается от аналогичной закономерности на территории всех остальных подвидов рода Tetrao: в первом случае количество осадков нарастает по мере снижения температур воздуха, во втором – уменьшается.

Рис. 3. Связь между температурой воздуха и количеством атмосферных осадков (расчетные значения) в ареале западноевропейских подвидов T. urogallus (R2 = 0.267 r = –0.516 p = 0.004; y = 1393.3 – 78.3 x). I – горы (абс. высоты 700–1825 м), II – равнины и низкогорья (16–485 м). Номера возле точек те же, что на рис. 1 и 2
Fig. 3. Connection between air temperature (axis X) and precipitation (Y) (calculated values) inside the range of T.urogallus in southern and central parts of Western Europe (R2 = 0.267 r = –0.516 p = 0.004; y = 1393.3 – 78.3 x). I – mountains (700–1825 m), II – plains and lowlands (16-485 m). The numbers next to circles and squares are the same as in figures 1 and 2
Удается также заметить, что по мере продвижения от низких к высоким значениям температуры область обитания рода Tetrao, по-видимому, расширяется по оси осадков (см. рис. 2). Для проверки этого предположения мы сопоставили значения размахов вариации осадков, в которые попадают популяции птиц, с соответствующими оценками температур (разбив их на интервалы в 1 оС, т. е. от –13.5 оС до –12.5 оС, от –12.4 оС до –11.5 оС… и от +7.5 оС до +8.5 оС). Результаты подтверждают предположение: температура определяет вариацию осадков в местах обитания всего рода в целом на 45 % (r = 0.67 p = 0.003), в местах обитания T. urogallus на 55 % (r = 0.75 p = 0.003); для T. parvirostris достоверной связи не выявлено, возможно, из-за небольшой выборки. При этом максимальные значения интервала осадков для всех принятых к анализу популяций в целом увеличиваются быстрее (y = 843.6 + 42.0x; r = 0.81 p < 0.001, где x – температура, y – осадки), чем минимальные (y = 482.6 + 16.7x; r = 0.88 p < 0.001). Следовательно, в более холодных условиях среды птицы осваивают места с более узким диапазоном осадков: низкая температура, по-видимому, повышает зависимость глухаря от атмосферных осадков и тем самым ограничивает использование пространства.
Обсуждение
Очевидно, что в пределах популяционных границ животные способны выбирать самые оптимальные участки, характеристики которых могут отличаться от средней величины для всего популяционного ареала. Поэтому использование в нашей работе именно средних популяционных значений температуры и осадков, возможно, несколько сместило расчетные оценки относительно реальности. Безусловно, однако, что эти смещения могут быть весомыми лишь для сравнений соседних или близких группировок, но почти или совсем незаметными на уровне крупных регионов и, тем более, подвидовых и видовых ареалов. Однако уточнение наших термогумидных оценок видится одной из непременных задач будущих исследований.
Анализ географических и климатических переменных показал достоверные, хотя и не самые сильные зависимости между ними (r = 0.55–0.86, см. табл. 1), а также выявил связи между самими географическими переменными (r = 0.21–0.68, Результаты). Даже эти немногочисленные факты свидетельствуют о проблемах в интерпретации результатов при использовании лишь географических переменных для изучения адаптаций птиц (см. Введение). В этом случае можно лишь предполагать воздействие целого ряда важных и второстепенных факторов, которые полностью или частично аккумулируются переменными широта, долгота и высота местности. Наш подход не оставляет сомнений: взаимное расположение популяций глухаря охарактеризовано именно в осях температуры и осадков.
Общая картина варьирования этих величин в пределах всего родового ареала видится вполне адекватной. Западный перенос воздушных масс над Евразией определяет снижение количества осадков и зимних температур воздуха по мере удаления от Атлантики (Алисов, Полтарус, 1974), что делает вполне логичным расположение большинства западноевропейских популяций в наиболее теплой и влажной области ареала, большинства восточносибирских – в холодной и сухой, остальные группировки занимают промежуточное положение (см. рис. 2).
Незанятость глухарем области низких температур и большого количества осадков (левая верхняя четверть на рис. 2) определяется тривиальным отсутствием такого типа термогумидного пространства: холодный воздух менее влагоемок относительно теплого. Этот же фактор диктует существенное и резкое сужение потенциально пригодного пространства и в левой нижней четверти (холодной и сухой). В этой четверти хорошо заметен зазор между большинством точек и границей реально существующего термогумидного пространства (см. рис. 2). По всей видимости, этот зазор маркирует территории равнинных тундр и горной растительности выше линии леса: в холодных условиях увеличение влажности подавляет лесную растительность (Пузаченко, 1985), блокируя распространение глухаря, т. к. данный род представлен дендрофильными видами (см. Введение). По всей видимости, узость термогумидного пространства каменного глухаря по оси осадков определяется весьма скромными размерами такого пространства, реально существующего в холодной зоне Северной Евразии. Развитие в этом секторе безлесных сообществ дополнительно ограничивает потенциально пригодное для птиц термогумидное пространство. На его пределы выходят всего две популяции из противоположных частей географического родового ареала: с плато Путорана и из горных районов северной Тувы (точки 78 и 76, см. рис. 2). На их склонах действительно отмечается повышенное количество осадков (Гвоздецкий, Михайлов, 1978). Вероятно, это один из наиболее восточных пиков осадков, обусловленных одновременно и орографией, и западным переносом атлантических воздушных масс. Значительное количество осадков на Камчатке, Сахалине и в Приморье (точки 89, E, F) определяется уже влиянием Тихого океана. Таким образом, относительная узость термогумидного пространства каменного глухаря по сравнению с обыкновенным – следствие объективных причин: в ареале первого вида условия среды более однообразны. И T. parvirostris осваивае, по-видимому, все потенциально пригодное для него пространство. Просматривается лишь единственное ограничение, имеющее отношение к термогумидным условиям: наличие лесной среды.
Географические ареалы обыкновенного и каменного глухарей перекрываются (см. рис. 1), что определяет и отмеченное нами перекрывание их термогумидного пространства (см. Результаты). Однако в осях температуры и осадков это перекрывание оказывается очень большим. Так, расположение крайних восточных точек ареала T. parvirostris в этих осях (№ 89, E и F, см. рис. 2) указывает на его потенциальную способность осваивать обширные территории, занятые сегодня только обыкновенным глухарем, включая Среднюю и Западную Сибирь (№ 67, 71, 74), Урал, Предуралье (54–56, 58, 60), и даже многие районы Фенноскандии (2, 9, 10, 12–14). Весьма вероятно, что современный адаптивный потенциал T. parvirostris к термогумидным условиям среды вполне позволяют ему занять значительную часть колоссальных пространств современного распространения «русских» и фенноскандийских подвидов T. urogallus (группы I и III на рис. 1). Напротив, потенциальное продвижение T. urogallus в термогуминое пространство T. parvirostris представляется ограниченным: максимум до юго-востока Якутии (№ 85), северное, северо-восточные и даже юго-восточные направления, по-видимому, исключены.
Следовательно, зона межвидового перекрывания термогумидного пространства показывает на явное неравенство двух видов по возможностям к потенциальному расселению: они огромны у каменного глухаря и очень невелики у обыкновенного. Такие различия позволяют предположить, что восточная граница видового ареала T. urogallus находится недалеко от предельной черты, возле которой его видовые адаптации к изученным нами переменным оказываются исчерпанными; для дальнейшего продвижения на север, восток и юго-восток необходимы дополнительные приспособления, выходящие, по-видимому, за рамки видовых возможностей. Не исключено, что вопрос о восточной границе ареала T. urogallus будет решен при более детальном изучении его адаптаций именно к термогумидным условиям среды. Напротив, современные знания биологии T. parvirostris (Исаев, 2014) не показывают явных абиотических или ценотических ограничений для экспансии вида в западном направлении, т. е. к более благоприятным условиям (мягче климат, разнообразнее растительность). Факторы, блокирующие такую экспансию, с уровня современных познаний просматриваются очень смутно. Возможно, что один из них – межвидовые отношения: в местах совместного обитания обыкновенный глухарь способен вытеснять каменного (Кирпичев, 1974).
Несколько неожиданным представляется взаимное расположение некоторых популяций в термогумидном пространстве T. urogallus. Так, максимального количества осадков следовало ожидать для крайних западных регионов, т. е. для Шотландии (№ 1, см. рис. 1), Норвегии (№ 3, 4) и Кантабрийских гор (№ 15). Однако расчетные значения показали умеренные оценки для этих точек и их близость к линии, разделяющей весь массив данных на сухие и влажные регионы (см. рис. 2). Среди рассмотренных популяционных арен максимальное количество осадков получают Юрские горы (№ 19, рис. 2), Французские Альпы (№ 20), Швейцария (№ 28), в меньшей мере Италия (№ 29), Австрия (№ 30), Словения (№ 31), Бавария (№ 18), а также Пиренеи (№ 16 и 17).
Равнинные районы Франции не препятствуют проникновению влажного воздуха вглубь континента. Хорошо выраженный орографический генезис большого количества осадков в этом регионе просматривается по Вогезам (№ 18), представляющим собой пусть невысокую, но первую преграду на пути господствующих западных ветров. Обширные и высокие горные массивы – Пиренеи и Альпы, встающие на пути влажных морских воздушных масс, получают и наибольшее количество осадков. Эти же массы, переваливая через невысокие горы Шотландии (максимум < 1400 м), по-видимому, теряют бóльшую часть влаги на западных склонах (см. Moss, 2015), что определяет и меньшее количество осадков в Восточной Шотландии, в том числе и по сравнению с Альпами и Пиренеями. Однако не привлеченные к нашему анализу популяции Западной Шотландии и, вероятно, юго-западной Норвегии все же существуют в условиях не меньшего количества осадков, чем альпийские и пиренейские.
Вопреки ожиданию, предполагавшему общее планетарное нарастание температуры от полюсов к экватору, самыми теплыми местами обитания глухаря (> +7.9 оС) оказались регионы Германии: (№ 21–26, см. рис 3). Эти далеко не самые южные точки ареала (см. рис. 1) лежат на низких гипсометрических уровнях (16–483 м), что, по всей видимости, и является основной причиной полученных для них высоких температурных показателей. Европейские популяции из южных широт (№ 15–17, 20, 29, 36–40) располагаются гораздо выше (1200–1825 м) и вполне логично получают меньше тепла: от +3.2 до +7.7 оС (см. рис. 3). То есть фактор «высота местности» полностью нивелирует влияние фактора «географическая широта».
Поражает разнообразие условий обитания фенноскандийских популяций (№ 1–11, см. рис. 2). Располагаясь географически компактно (см. рис. 1), они охватывают почти весь термический интервал, занятый популяциями Русской и Западно-Сибирской равнин (№ 49–66, см. рис. 2). По оси осадков фенноскандийские группировки занимают пространство, вмещающее почти все популяции T. urogallus за исключением некоторых западноевропейских. Такая вариация наших оценок для Фенноскандии – очевидное следствие повышенного разнообразия ее климатических условий.
Следует особо отметить несколько точек, лежащих одновременно и в самой теплой, и относительно сухой области родового ареала: Саксония (№ 24, см. рис. 3), Померания (№ 34), Люблин (№ 35), Беловежская Пуща (№ 42) и Украинское Полесье (№ 44). По всей видимости, примерно эта территория рассматривалась как место обитания глухарей с максимальным для всего ареала весом тела самцов (Zedlitz, 1924; Couturier, Couturier, 1980) и противопоставлялась популяциям с мелкими особями, в основном из австрийских (№ 30) и Баварских (№ 27) Альп (Zedlitz, 1933). Наш материал позволяет предположить, что одной из причин обозначенных различий веса может быть не только и не столько межрегиональное расхождение показателей температуры (всего на 1.5–2.0 оС), но почти двукратная разница по осадкам (580–605 мм против 1140–1180 мм, см. рис. 3). По-видимому, сочетание этих двух факторов способно прямо или косвенно влиять на рост и развитие глухаря. Вероятно, что его оптимальные условия обитания лежат в области относительно высоких температур с небольшим количеством осадков. Возможно, однако, что это справедливо лишь для группы западноевропейских подвидов или только для T. u. major.
Негативное влияние повышенного количества осадков на глухаря обычно рассматривается с позиций выживания птенцов и обновления популяций (Теплов, 1947; Гаврин, 1956; Lindén, 1981; Ménoni, 1991; Watson, Moss, 2008). Наш материал показал, что в местах его обитания интервал осадков сокращается по мере снижения температуры. Этот факт позволяет предполагать общее негативное воздействие сырого климата на население рода Tetrao, включая взрослых особей: и на них большое количество осадков, в основном дождей, вероятно, также оказывает отрицательное влияние. Например, через увеличение термозатрат на просушивание оперения и/или сокращение бюджетов времени на кормодобывание из-за необходимости укрываться от сильных дождей и некоторое время после них.
Интересной находкой нашей работы представляется принципиальная разница между ареалом западноевропейской подвидовой группы (T. u. major, T. u. cantabricus, T. u. aquitanicus, T. u. rudolfi) и, похоже, всех остальных популяций рода Tetrao по закономерности пространственного изменения температуры и осадков. Основными причинами этого видятся относительно небольшая площадь западноевропейского ареала и сильная вариация рельефа в его пределах. Первый фактор позволяет лишь в очень слабой мере проявиться как широтным закономерностям изменчивости климата, так и всем следствиям западного переноса воздушных масс в Евразии. Обе эти закономерности, по-видимому, просто не обнаруживаются в заметной мере из-за тривиального отсутствия места, арены их реализации в Западной Европе. В такой ситуации возрастает роль рельефа: пространственные изменения и температуры, и осадков порождаются в основном гипсометрическими причинами, которые и диктуют обратную связь температуры с осадками: чем выше в горы, тем меньше тепла и больше влаги. По-видимому, именно высокая неоднородность рельефа западной Европы является главной причиной обсуждаемых различий, т. к. для ареала фенноскандийской группы, населяющей в основном равнины и низкогорья, отмечена иная закономерность, присущая для обширных пространств центральных и восточных частей родового ареала. В их пределах циркуляция воздушных масс по долготе отодвигает на второстепенный план влияние даже крупных горных систем, демонстрируя прямую связь между температурой и осадками: чем восточнее и дальше от Атлантики с ее Гольфстримом, тем меньше тепла и меньше влаги.
Эти закономерности мало интересны для систем с жесткой фиксацией объектов в пространстве. Но птицы мобильны, и перемещения, включая дальние, часто нецеленаправленные, способны выводить их в пессимальные условия среды, например в холодные. Важно, что живые организмы обычно легче переносят холод в условиях сухости. И в таком контексте выявленные нами пространственные различия термогумидных изменений приобретают особое значение. Для его демонстрации выделим два типа климатических градиентов. Один из них характерен для западной, центральной и южной Европы в целом: смещение от высоких температур к низким сопровождается увеличением количества осадков, что дополнительно (к низким температурам) затрудняет условия существования. Поскольку ужесточение условий среды требует повышения энергозатрат на обеспечение жизни (Дольник, 1995), назовем такой градиент «жестким». Противоположный тип – «мягкий» градиент – характерен для большинства мест Северной Евразии: смещение вниз по шкале температур сопровождается нарастанием сухости, что должно определять менее ощутимое ухудшение жизненных условий; существенных климатических барьеров на ее колоссальных пространствах почти нет, изменения плавные, постепенные.
Напомним, что иерархичность пространственной структуры популяций диктует универсальность групповых адаптаций: для всей популяции в целом отбор закрепляет те из них, которые адекватны на всем ее ареале, прочие, менее значимые, адаптации жизнеспособны лишь на локальных уровнях (Северцов, 2013). Следовательно, в условиях жесткого градиента среды даже ограниченные пространственные экспансии должны сопровождаться существенными адаптивными перестройками. Мягкий градиент, вероятно, более толерантен к иммигрантам и требует от них заметных адаптивных сдвигов лишь при колонизации очень удаленных пространств.
В условиях крайне жесткого градиента среды локальные адаптации должны быть выражены четко, пространства, осваиваемые их носителями, – небольшими, а сами популяции обречены, по-видимому, представлять собой относительно изолированные группировки специалистов-стенотопов; межвидовое скрещивание и инбридинг вполне реальны, т. к. дальние миграции успешны лишь для особей с широчайшей нормой реакции. Противоположная крайность (самый мягкий градиент среды) предполагает функциональную близость локальных и более общих климатических адаптаций. Внутривидовая пространственная структура должна быть расплывчатой, с нечеткими границами соседних территориальных группировок и постепенными пространственными изменениями генотипов. Межвидовое скрещивание вероятно лишь в условиях низкой численности, а дальние миграции вполне обычны и могут быть успешными для широкого круга особей.
Такой взгляд на климатические градиенты позволяет понять, в частности, сильный разброс по термогумидному пространству фенноскандийских популяций: миграции даже из районов Сибири в европейскую часть России и далее к северо-западу, равно как и в обратном направлении, не требуют существенных адаптивных перестроек. Мягким градиентом среды удобно объяснить поразительное генетическое единство населения русского глухаря на колоссальных пространствах от Архангельска и Твери до Красноярска (Duriez et al., 2007), а также пошаговые изменения гаплотипов: различия между западноевропейскими (включая шведских) и русскими глухарями больше, чем между финскими и русскими (Liukkonen-Anttila et al., 2004). Понятнее становятся и различия в миграционной активности глухаря, повышенной в северо-западной части России (Borchtchevski, Moss, 2014) или в Швеции (K. Sjöberg, цит. по: Åhlen et al., 2013) и весьма скромной в центре Западной Европы (Storch, 1993). Без представлений о разных климатических градиентах среды все эти факты – лишь перечень несвязанных явлений. Предлагаемый нами взгляд позволяет заметить в беспорядочном нагромождении таких фактов определенную систему: разные, возможно, несколько непривычные адаптивные комплексы (по-видимому, территориальные) поведенческих, физиологических, генетических и, вероятно, иных реакций теплокровных животных на несовпадающие тренды среды обитания.
К сожалению, подробное обсуждение термогумидных градиентов, которое непременно должно включать уточнение границ выделенных нами подвидовых групп и их списки, выходит за рамки данной работы. Заметим лишь, что сам феномен разнонаправленных связей между температурой и осадками, который, по-видимому, пока не попадал в поле зрения специалистов, безусловно, нуждается в проверке и осмыслении.
Заключение
Общее расположение центров реально существующих и недавно вымерших популяций глухаря в осях температуры и осадков выглядит вполне закономерным: в наиболее теплой и влажной области родового ареала располагаются группировки центральных и южных районов Западной Европы, в холодной и сухой – большинство восточносибирских, остальные популяции занимают промежуточное положение. Такое размещение подтверждает корректность большинства климатических моделей, созданных для описания общего термогумидного пространства рода Tetrao.
Глухарь практически полностью отсутствует в области отрицательных среднегодовых температур со значительным количеством осадков (>800–900 мм/год), что, по-видимому, связано с крайней ограниченность такого типа термогумидного пространства на суше Северной Евразии.
По мере продвижения от максимальных к минимальным температурам занимаемое родом термогумидное пространство сжимается по оси осадков, показывая тренд к более широкому использованию местообитаний с бóльшим количеством осадков при нарастании температур.
Перекрывание видовых зон термогумидного пространства представляется существенно шире перекрывания видовых географических ареалов. Пределы занятого обыкновенным глухарем термогумидного пространства примерно соответствуют современным границам его географического распространения, что позволяет рассматривать тепло и осадки в качестве ограничителей его потенциальной экспансии в восточном направлении. Однако эти же факторы, по всей видимости, не препятствуют встречной экспансии каменного глухаря в западном и северо-западном направлениях вплоть до Фенноскандии.
Разные популяции глухаря обитают в условиях несовпадающей пространственной ковариации тепла и влаги. На площади обитания большей части родового населения количество осадков сокращается при движении от теплых районов к холодным. Однако по тому же градиенту на территории южного и центрального секторов Западной Европы количество осадков не снижается, а нарастает. Эти ранее не отмечавшиеся различия, вероятно, способны формировать комплексы несовпадающих групповых адаптаций. Обнаруженные различия, возможно, позволят подобрать ответы к известным, но труднообъяснимым сходствам и различиям пространственных группировок глухаря по генетике, морфологии, экологии, пространственной подвижности и, вероятно, каким-то иным признакам.
Ранжирование конкретных популяций в термогумидном пространстве показало, что вопреки ожиданию в наиболее теплых условиях видового ареала расположены не самые южные группировки (испанские, балканские, болгарские), а популяции северо-западной Германии. Также вопреки ожиданию климатические модели показали максимальное количество осадков в не местах обитания приатлантических группировок (Кантабрия, вост. Шотландия, Норвегия), а в основном в биотопах птиц из северо-западных предгорий Альп (Вогезы, Юра), из центральных и южных районов этой горной страны (Швейцария, Австрия, Бавария, Словения, Италия), а также из испанских Пиренеев. Хотя не включенные в анализ популяции из западной Шотландии и, вероятно, с юго-запада Норвегии существуют, по-видимому, в условиях максимального количества осадков. Фенноскандийские подвиды осваивают обширную часть термогумидного пространства, заходя далеко в сухую зону низких температур, занятую «русскими» популяциями, в том числе и сибирскими.
Библиография
Алисов Б. П., Полтараус Б. В. Климатология . М.: МГУ, 1974. 299 с.
Васильев Н. Г., Горин Д. А., Рашек В. Л., Рогачева Э. В., Русанова О. М., Скокова Н. Н., Сыроечковский Е. Е., Чумакова А. В., Язан Ю. П. Заповедники СССР . М.: Лесная промышленность, 1980. 240 с.
Гаврин В. Ф. Экология тетеревиных птиц Беловежской Пущи: Дис. … канд. биол. наук . Алма-Ата, 1956. 352 с.
Гвоздецкий Н. А., Михайлов Н. И. Физическая география СССР. Азиатская часть . М.: Мысль, 1978. 512 с.
Горичев Ю. П. Природные особенности Южно-Уральского государственного природного заповедника // Труды Южно-Уральского государственного природного заповедника. Вып. 1. Уфа, 2008. С. 13–56.
Добрынин Б. Ф. Физическая география Западной Европы . М.: Учпедгиз, 1948. 416 с.
Дольник В. Р. Ресурсы энергии и времени у птиц . СПб.: Наука, 1995. 360 с.
Исаев А. П. Тетеревиные птицы Якутии: распространение, численность, экология: Дис. … д-ра биол. наук . Якутск, 2014. 463 с.
Кириков С. В. Род Глухари Tetrao Linnaeus. 1758 // Птицы Советского Союза / Ред. Г. П. Дементьев, Н. А. Гладков. М., 1952. Т. 4. C. 84–107.
Кирпичев С. П. О распространении каменного и обыкновенного глухарей в Заенисейской Сибири // Материалы 6-й Всесоюзной орнитологической конференции. Ч. 2. М.: Изд-во Моск. ун-та, 1974. С. 63–64.
Климатические данные городов по всему миру . URL: http://climate-data.org (дата обращения 05.12.2017).
Мильков Ф. Н., Гвоздецкий Н. А. Физическая география СССР. Общий обзор. Европейская часть. Кавказ . М.: Мысль, 1976. 448 с.
Нейфельд Н. Д., Бобрецов А. В. О гнездовании глухаря в горнотундровом поясе Северного Урала // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Вып. 7. Екатеринбург, 2002. С. 207.
Островский А. И. Некоторые материалы к размножению карпатского глухаря // Вестник зоологии № 4. Киев: Наукова думка, 1973. С. 21–24.
Пузаченко Ю. Г. Климатическая обусловленность южной границы тундры // Сообщества Крайнего Севера и человек / Под ред. Ю. И. Чернова. М.: Наука, 1985. С. 22–56.
Равкин Е. С., Равкин Ю. С. Птицы равнин Северной Евразии. Численность, распределение и пространственная организация сообществ . Новосибирск: Наука, 2005. 303 с.
Романов А. А. (ред.) Фауна позвоночных животных плато Путорана . М., 2004. 457 с.
Северцов А. С. Эволюционная экология позвоночных животных . М.: Товарищество научных изданий КМК, 2013. 347 с.
Теплов В. П. Глухарь в Печеро-Ылычском заповеднике // Труды государственного Печеро-Ылычского заповедника. Вып. 4. Ч. 1. М., 1947. С. 3–76.
Термограф: архивные данные температуры воздуха и количества осадков . URL: http://thermograph.ru (дата обращения 05.12.2017).
Хромов С. П., Петросянц М. А. Метеорология и климатология . М.: Наука, 2006. 582 с.
Чернов Е. Г. Что растет на Кольской земле // Природа Мурманской области. Мурманск: Мурманское книжное изд-во, 1964. С. 185–224.
Åhlen P.-A., Willebrand T., Sjöberg K., Hörnel-Willebrand M. Survival of female capercaillie Tetrao urogallus in northern Sweden // Wildlife Biology. 2013. Vol. 19. P. 368–377.
Balasubramaniam P., Rotenberry J. T. Elevation and latitude interact to drive life-history variation in precocial birds: a comparative analysis using galliformes // Journal of Animal Ecology. 2016. Vol. 85. P. 1528–1539.
Bears H., Martin K., White G. C. Breeding in high-elevation habitat results in shift to slower life-history strategy within a single species // Journal of Animal Ecology. 2009. Vol. 78. P. 365–375.
Beniston M. Mountain weather and climate: a general overview and focus on climatic change in the Alps // Hydrobiologia. 2006. Vol. 562. P. 3–16.
Borchtchevski V., Moss R. Age structure of Capercaillie males (Tetrao urogallus) in NW Russia may reflect two-way movements – a hypothesis // Ornis Fennica. 2014. Vol. 91. P. 14–28.
Boyle W. A., Sandercock B. K., Martin K. Patterns and drivers of intraspecific variation in avian life history along elevational gradients: a meta-analysis // Biological Reviews. 2016. Vol. 91. P. 469–482.
Čas M. Disturbances and predation on capercaillie at leks in Alps and Dinaric mountains // Šumavski list. 2010. № 9–10. CXXXIV. P. 487–495.
Clima Temps.com. URL: http://www.climatemps.com/countries-a.php (дата обращения 05.12.2017).
Couturier M., Couturier A. Les coqs de bruyère. Le grand coq de bruyère Tetrao urogallus urogallus L. Ed. F. Dubusc. Boulogne, 1980. Vol. 1. 656 p.
Donázar J. A. Geographic variation in clutch and brood size of the eagle owl Bubo bubo in the Western Palearctic // Journal für Ornithologie. 1990. Bd. 131. S. 439–443.
Dunn P. O., Thusisu K. J., Kimber K., Winkler D. W. Geographical and ecological variation in clutch size of tree swallows // Auk. 2000. Vol. 117. P. 215–221.
Duriez O., Sachet J.-M., Ménoni E., Pidancier N., Miquel Ch., Taberlet P. Phylogeography of the capercaillie in Eurasia: what is the conservation status in the Pyrenees and Cantabrian Mounts? // Conservation Genetics. 2007. Vol. 8. P. 513–526.
Hille S. M., Cooper C. B. Elevation trends in life histories: revising the pace-of-life framework // Biological Reviews. Cambridge Philosophical Society. 2015. Vol. 90. P. 204–213.
Jetz W., Sekercioglu C. H., Böhning-Gaese K. The worldwide variation in avian clutch size across species and space // PLОS Biol. 2008. Vol. 6 (12): e303. DOI: 10.1371/journal. pbio.0060303.
Lindén H. Estimation of juvenile mortality in capercaillie, Tetrao urogallus, and black grouse, Tetrao tetrix, from indirect evidence // Paper Game Research. 1981. Vol. 39. P. 35–51.
Liukkonen-Anttila T., Rätti O., Kvist L., Helle P., Orell M. Lack of genetic structuring and subspecies differentiation in the capercaillie (Tetrao urogallus) in Finland // Annales Zoologici Fennici. 2004. Vol. 41. P. 619–633.
Lu X. Reproductive ecology of three Tibetan waterbird species, with special reference to life-history alterations along elevational gradients // Zoological Studies. 2011. Vol. 50. P. 192–292.
Martin T. E. Avian life history evolution in relation to nests sites, nest predation, and food // Ecological Monographs. 1995. Vol. 65. P. 101–127.
Martin T. E., Bassar R. D., Bassar S. K., Fontaine J. J., Lloyd P., Mathewos H. A., Niklison A. M., Chalfoun A. Life-history and ecological correlates of geographic variation in egg and clutch mass among passerine species // Evolution. 2006. Vol. 60. P. 390–398.
McNamara J. M., Barta Z., Wikelski M., Houston A. I. A theoretical investigation of the effect of latitude on avian life histories // American Naturalist. 2008. Vol. 172. P. 331–345.
Ménoni E. Ecologie et dynamique des populations du grand tétras dans les Pyrénées, avec des references speciales à la biologie de la réproduction chez les poules – quelques applications à sa conservations. Thése. Université Paul Sabatier de Toulouse, 1991.
Moss R. Global warming and grouse Tetraoninae population dynamics // Grouse News. 2015. № 50. P. 8–18.
OpenStreeMap. URL: http://www.openstreetmap.org/#map=9/47.3472/12.1014 (дата обращения 05.12.2017).
Potapov R., Sale R. Grouse of the world. London, Cape Town, Sydney, Auckland, 2013. 408 p.
Storch I. Habitat use and spacing of capercaillie in relation to forest fragmentation patterns. Thesis. University of Munich, 1993. 98 p.
Watson A., Moss R. Grouse. The natural history of British and Irish species. London, Collins ed., 2008. 529 p.
Zedlitz O. Das Gewichts als Rassenmerkmal bei Tetrao urogallus // Journal für Ornithologie. 1924. Bd. 72. S. 244–252.
Zedlitz O. Das Gewichts als Rassenmerkmal bei Tetrao urogallus (II. Teil) // Berichte des Vereins Schlesischer Ornithologen (Breslau). 1933. Bd. 18. S. 7–14.
Благодарности
Авторы признательны А. С. Северцову, не пожалевшему времени на неоднократные замечания к структурной организации исходного текста, А. В. Артемьеву, А. В. Коросову и трем анонимным рецензентам за конструктивную критику рукописи, а также М. В. Яковлевой, В. А. Алексееву, С. В. Буслаеву, А. С. Гилязову, В. П. Иванчеву, Н. Д. Нейфельду и А. В. Сивкову за климатические данные и координаты метеостанций. Отдельная благодарность В. Н. Алексееву, А. С. Гилязову, B. Leclerqc, R. Moss и Т. К. Spidsø за содействие в оценке границ высотного размещения глухаря и помощь в библиографических исследованиях.

 © 2011 - 2026
© 2011 - 2026