



Введение
В серии предыдущих статей по фаунистическому и флористическому районированию Северной Евразии подведены итоги работ сначала отдельно по всем классам наземных позвоночных, затем по рыбам и круглоротым и, наконец, по всем этим группам вместе (обобщение см. в статье Равкина, Богомоловой, Цыбулина, 2015а), а также по беспозвоночным животным и флоре древесных растений (Равкин и др., 2014, 2015б). Цель такого анализа и, соответственно, этой статьи, помимо сопоставления вариантов частного районирования, выполненного на одинаковой основе и единым набором подходов и вычислительных средств, – переход к комплексному биотическому районированию. Обычно фаунистическое районирование проводили по отдельным классам животных (Северцов, 1877; Семенов-Тян-Шанский, 1936; Кузнецов, 1950; Мекаев, 1987), а по флоре – чаще в целом и лишь иногда по отдельным группам растений (Курнаев, 1973; Равкин и др., 2015б). Единое фаунистическое районирование проводили на примере отдельных представителей тех или иных классов экспертно-умозрительно (Семенов-Тян-Шанский, 1936), то есть без применения формализованных методов сопоставления и классификации, хотя это вполне корректный подход теоретического обобщения. Лишь в последнее время для решения таких задач применены методы непараметрической статистики (Малышев, 2000; Бобров, Алещенко, 2001; Блинова, Равкин, 2008; Holt et al., 2013), которые способствуют сравнимости полученных результатов. К сожалению, в идеале проведение единого флоро-фаунистического районирования пока невозможно. Для решения этой задачи следовало бы выполнить все варианты районирования после решения всех споров в систематике растений и животных и полного завершения сбора данных по флоре и фауне. Но и после этого провести анализ биоты в целом будет весьма затруднительно, т. к. число видов даже сосудистых растений, а соответственно, и сходство за счет флоры может быть существенно выше, чем беспозвоночных животных, количество видов и разнообразие которых значительно больше, чем позвоночных. Поэтому при объединении повидовых списков закономерности неоднородности флоры подавят специфику изменчивости фауны в случае их несовпадения. Поэтому статью, предлагаемую вниманию читателей, можно рассматривать как опыт использования иного способа объединенного флоро-фаунистического районирования.
Материалы
Материалы, методы и подходы к этим исследованиям изложены ранее в публикациях авторов данной статьи, перечисленных во введении. Алгоритмы классификации и оценки связи с факторами среды и их неразделимыми сочетаниями (природными режимами) тоже опубликованы (Трофимов, 1976, 1978; Равкин и др., 1978; Трофимов, Равкин, 1980; Челинцев, 2000). Территория Северной Евразии (в границах СССР 1991 г.) разделена на 597 участков так, как это сделано в базе данных «Биодат» (http:www.biodat.ru). Сведения о встречаемости животных и древесных растений, а также об использованной карте-основе заимствованы из этого портала с уточнениями по источникам, упомянутым в статьях, посвященных вариантам частного районирования (см.: Равкин и др., 2014, 2015б). Общая подборка данных включает показатели встречаемости 2685 видов животных и растений (земноводных – 40 видов, пресмыкающихся – 170, птиц – 734, млекопитающих – 341, рыб и круглоротых – 335, жуков – 529, древесных растений – 536 видов). Выполненное исследование, с нашей точки зрения, достаточно актуально. Подобные работы могут быть использованы, как считают Хольт и др. (Holt et al., 2013), при планировании природоохранных мероприятий.
В данной работе принято следующее допущение. Все таксоны использованной при районировании классификации выделены в том случае, если в каждый из них входит не менее восьми участков. Кластеры с меньшим их числом включены в представительные таксоны того же уровня, с которыми они граничат и максимально сходны.
Методы
Один из возможных выходов из ситуации, описанной во введении, представляет собой следующий способ анализа. Сначала необходимо провести частное районирование по отдельным классам наземных позвоночных, а затем по всей группе этих животных, фауна которых описана достаточно полно. Сведения о распространении беспозвоночных явно недостаточны, поэтому по ним целесообразнее проанализировать какую-нибудь эталонную подборку хорошо изученных видов. Такой группой можно считать хотя бы часть видов жуков. Мы использовали для этого сведения по короедам, немонихидам, ложнослоникам и трубковертам (Равкин и др., 2014; Ravkin et al., 2015). В качестве такой же эталонной флористической группы взяты древесные растения. Эти три подборки условно приняты в качестве равнозначных. Для этого рассчитанные по ним матрицы коэффициентов сходства усреднены. Такой подход показал свою эффективность при совместном анализе почв, морт-, фито- и зоомассы, значения по которым существенно различаются (Равкин и др., 2011). При анализе нашей выборки по флоре и фауне необходимость нормирования соответствующих матриц сходства отпадает, т. к. суммы коэффициентов и, соответственно, «веса» по каждой из них очень близки.
Результаты
При автоматической классификации выявлено восемь подобластей, поэтому объединение их в регионы, как это делали раньше, оказалось нецелесообразным. Половина подобластей разделена на провинции, одна из которых делится еще и на округа (рис. 1). Полная классификация приведена ниже.
Флоро-фаунистическое районирование Северной Евразии
Подобласти
- Полярно-пустынно-тундровая островная (полярно-пустынных участков 7, тундровых 4);
- Тундровая материковая кольско-чукотская (тундровых 54, редколесный 1).
Провинции
2.1 – Западная (тундровых 16);
2.2 – Срединная (тундровых 30, редколесных 1);
2.3 – Восточная (тундровых 8).
Подобласти
3. Тундрово-таежная северо-восточная притихоокеанская корякско-камчатская (тундровых 17, редколесных 4, среднетаежных 2);
4. Предтундрово-редколесная большеземельско-колымская (редколесных 69, северотаежных 3).
Провинции
4.1 – Западная (редколесных 14, северотаежных 2);
4.2 – Северо-восточная (редколесных 42, северотаежных 1);
4.3 – Юго-восточная (редколесных 13).
5. Таежно-степная балтийско-амурская подобласть (таежных 167, подтаежных и широколиственнолесных 23 и 14, степных и лесостепных 55 и 20, редколесных 3).
5.1 – Северная таежно-степная провинция (северо- и среднетаежных 29 и 62, южно- и подтаежных 19 и 16, горно-таежных 25, лесостепных и степных 6 и 7, редколесных и широколиственнолесных 3 и 1).
Округа
5.1.1 – Северо-западный таежный (северо- и среднетаежных 28 и 39, южно- и подтаежных 9 и 1, лесостепных 2);
5.1.2 – Восточный таежный (редколесных 3, таежных 46);
5.1.3 – Юго-западный южнотаежно-подтаежный (южно- и подтаежных 7 и 16, широколиственнолесных и лесостепных 1 и 3);
5.1.4 – Срединный таежно-степной (степных 7, лесостепных 1, таежных 6).
Провинции
5.2 – Юго-западная широколиственностепная (степных и лесостепных 31 и 14, широколиственнолесных 13, южно- и подтаежных по 2);
5.3 – Срединная степная (степных 11);
5.4 – Юго-восточная таежно-степная (таежных 30, подтаежных и степных 5 и 6).
Подобласти
6. Крымско-кавказская горная (горно-лесных 13, степных 12, полупустынных 4);
7. Таежно-лесная юго-восточная притихоокеанская уссурийско-сахалинско-курильская (средне- и южнотаежных 2 и 8, подтаежных и широколиственных по 2);
8. Пустынно-степная казахстанско-среднеазиатская (пустынных 51, полупустынных 33, степных 25).
Провинции
8.1 – Северо-западная степная (степных 14);
8.2 – Северо-восточная пустынно-полупустынная (полупустынных 9, пустынных 5);
8.3 – Срединная полупустынно-пустынная (пустынных 35, полупустынных 18);
8.4 – Южная полупустынно-пустынно-горностепная (горностепных 13, пустынных 11, полупустынных 6).
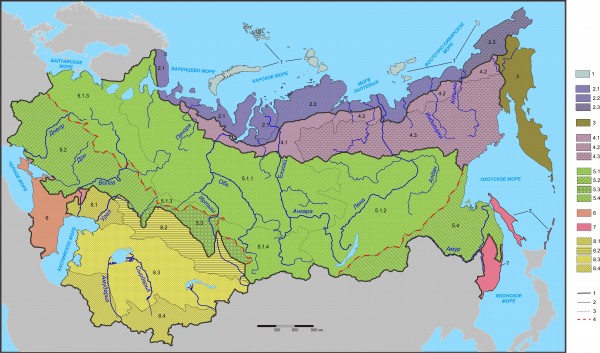
Рис. 1. Флоро-фаунистическое районирование Северной Евразии.
Характеристики таксонов см. выше в тексте классификации; границы: 1 – подобластей, 2 – провинций, 3 – округов, 4 – максимальной диагональности
Fig. 1. Floro-faunistic zoning of Northern Eurasia.
Characteristics of the taxa listed above in the text on classification; borders of: 1 – subregions; 2 – provinces; 3 – districts; 4 – maximum diagonality
Анализ зонально-подзональной принадлежности участков предварительного разделения, включенных в таксоны классификации, показывает значительное сходство их с зональным и подзональным делением территории, хотя стопроцентного совпадения состава зон и подобластей по участкам нигде нет. Так, в Полярно-пустынно-тундровой островной подобласти 64 % участков приходится на полярно-пустынные, а остальное – на тундровые. В Тундровой материковой подобласти номинальных (тундровых) участков 98 %, в Тундрово-таежной притихоокеанской их 74 %, а в Предтундрово-редколесной – 96 % редколесных участков. В Таежно-степной подобласти 72 % участков лесной зоны, а степных (если лесостепь считать подзоной степной зоны) – 27 %. Крымско-кавказская подобласть представлена преимущественно горно-лесными и степными участками (45 и 41 %), а юго-восточная притихоокеанская – на 100 % лесными участками. Почти половина Пустынно-степной казахстанско-среднеазиатской подобласти приходится на территорию пустынных участков (47 %), на 30 % это полупустыни, на 23 % степи. Сопоставляя подобный анализ по таксонам частного районирования, можно говорить о большей плавности флоро-фаунистических изменений и большей степени совпадения их с зональными.
Есть еще одна особенность неоднородности общего состава флоры и фауны. При частном районировании отдельно по группам животных и растений четко и в разной степени проявляется диагональность, то есть смещение границ выделяемых таксонов с северо-запада к юго-востоку, причем под разным углом к границам природных зон (с различной дифферентностью). При комплексном районировании северо-западная диагональная граница в пределах Европы и Западной Сибири смещена к юго-западу и приближается к зональному разделению. В то же время ранее не значимое диагональное смещение в Восточной Сибири и на Дальнем Востоке (в связи с более слабым отепляющим влиянием Тихого океана по сравнению с Атлантическим) при комплексном районировании четко прослеживается вплоть до Забайкалья, хотя в северной части Приохотья из-за горных хребтов южная граница Предтундрово-редколесной подобласти смещена к югу. Провинциальность, определяемая континентальностью и горными массивами, прослежена и при первом формализованном разбиении, хотя и в виде непредставительных включений. Так, в островной подобласти самая западная и самая восточная части (Земля Франца Иосифа и остров Врангеля) не приняты в качестве самостоятельных подобластей только из-за малой их представительности (соответственно два и один участок). То же самое отмечено и в Тундровой подобласти (Кольский полуостров), но в ней, в отличие от первой подобласти, восточное отклонение уже достаточно представительно и дает основание для выделения отдельной подобласти – Северо-восточной притихоокеанской. В таежно-степной части территории таких подобластей две: Крымско-кавказская на западе и Юго-восточная притихоокеанская на востоке.
Таким образом, можно констатировать наличие четырех широтных флоро-фаунистических полос, примерно соответствующих полярным пустыням, тундрам, предтундровым редколесьям и широтной полосе от северной тайги до северных (луговых) степей. Кроме того, имеются как минимум два клинальных включения – более южных флор и фаун: средиземноморских в Крым и на Кавказ, а на востоке, в юго-восточной притихоокеанской подобласти, японско-китайских флор и фаун в Уссурийский край, на Сахалин и Курилы. Причины этих проникновений – в отепляющем влиянии Средиземноморья на западе и муссонов на юго-востоке. Казахстанско-среднеазиатскую подобласть тоже можно рассматривать как клинальное включение полупустынно-пустынных условий с юга и соответствующей специфичности флор и фаун. Отепляющее влияние Атлантики приводит к диагональному смещению границ с северо-запада на юго-восток, а Тихого океана – с северо-востока на юго-запад. При этом отклонение от этого тренда четко видно в Северном Приохотье из-за влияния горных хребтов Восточной Сибири и Дальнего Востока.
По отношению к карте Г. Д. Рихтера (1964) диагональ юго-восточного смещения сначала отсекает на западе часть южнотаежно-подтаежной полосы, затем восточнее Тулы опускается до границы широколиственно-лесостепной полосы. Далее за Омском диагональная граница пересекает эту полосу и идет почти вдоль границы со степной зоной, слегка заходя в ее северную оконечность. Таким образом, диагональ пересекает с северо-запада на юго-восток три подзональных полосы. Аналогичное смещение с северо-востока на юго-запад от Тихого океана сначала идет по южной границе полосы темнохвойно-лиственничных лесов и редколесий, затем рассекает ее и примыкает к западным таежным елово-березовым редколесьям с тундрами и лугами Забайкалья. Таким образом, и это диагональное смещение тремя ступенями пересекает две природно-географические полосы.
Пространственно-типологическая структура и экологическая организация флоро-фаунистической неоднородности
На структурном графе флоро-фаунистической изменчивости в Северной Евразии четко прослеживаются два основных тренда (рис. 2).
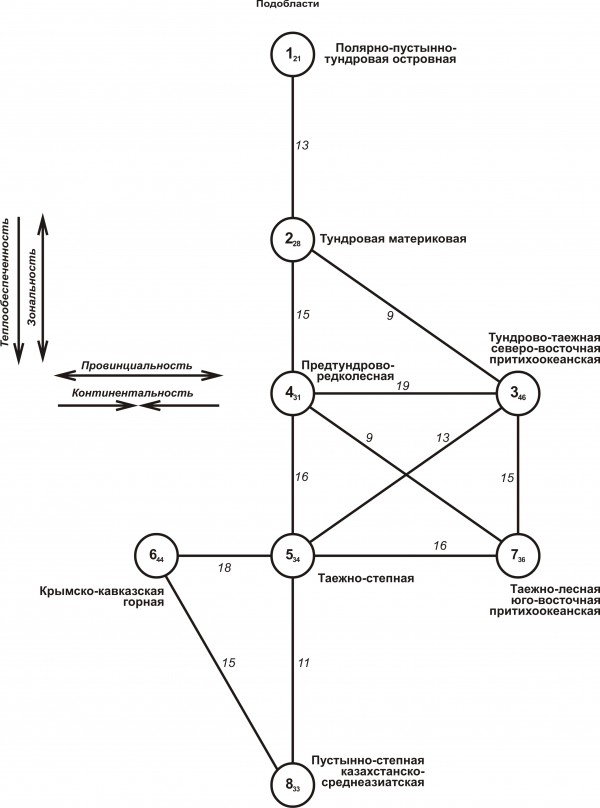
Рис. 2. Пространственно-типологическая структура флоро-фаунистической неоднородности Северной Евразии на уровне подобласти. Внутри значков приведены номера таксонов классификации (см. в тексте), в виде подстрочного индекса показана величина внутригруппового сходства. Между таксонами обозначена величина межгруппового сходства. Стрелки у перечня основных структурообразующих факторов среды указывают направление увеличения их влияния и флоро-фаунистические тренды
Fig. 2. Spatial-typological structure of the Floro-faunistic heterogeneity of northern Eurasia at the level of subregion. The numbers of the taxa see in the text, a subscript shows the value of intra-group similarity. Between the taxa the value of intergroup similarity is indicated . The arrows near the list of basic structure-forming environmental factors indicate the direction of increasing their influence and floro-faunistic trends
Первый из них иллюстрирует широтный характер изменений (от полярных до жарких пустынь через тундры, предтундровые редколесья, тайгу, леса, лесостепь и степи). Второй тренд имеет долготное анклавное простирание, представленное Крымско-кавказской подобластью на юго-западе, а также северо- и юго-восточными притихоокеанскими подобластями на востоке. Первое определяет отепляющая роль Средиземноморья и барьерная защита горными массивами от северного переноса холодных воздушных масс. Второе связано с отепляющим воздействием Тихого океана.
Оценка связи пространственной изменчивости факторов среды с неоднородностью флоры и фауны Северной Евразии
Estimates of correlation between of the spatial variability of environmental factors and floro-faunistic heterogeneity in Northern Eurasia
| Фактор, режим | Учтенная дисперсия, % |
| Теплообеспеченность (зональность+ провинциальность + поясность) | 62 |
| Зональность | 50 |
| Провинциальность | 20 |
| Островной эффект | 4 |
| Высотная поясность | 0.6 |
| Все факторы | 72 |
| Режимы: | |
| классификационные | 63 |
| структурные | 57 |
| Все режимы | 70 |
| Все факторы и режимы | 83 |
| Районирование*: | |
| климатическое | 42 |
| биогеографическое (биомное) | 39 |
| флористическое | 33 |
| физико-географическое | 27 |
| лесорастительное | 22 |
Примечание. * – варианты районирования заимствованы из литературы (Флора СССР, 1937; Курнаев, 1973; Udvardy, 1975; Воронов, Кучерук, 1977; Атлас СССР, 1983).
Итак, судя по классификации, можно говорить о влиянии на изменчивость флоры и фауны пяти скоррелированных факторов среды. Все они носят сборный характер, но в итоге могут быть сведены к различиям в теплообеспеченности (таблица). С нею можно связать 62 % дисперсии матрицы коэффициентов флоро-фаунистического сходства. Отдельно зональность и провинциальность учитывают соответственно 50 и 20 % дисперсии. Островной эффект и наличие гор определяют существенно меньшую часть неоднородности из-за относительно небольшой площади островов и горных массивов. Всеми перечисленными факторами можно объяснить 72 % дисперсии сходства, т. е. приращение к влиянию теплообеспеченности составляет по отдельным факторам среды всего 10 % дисперсии. Классификационные и структурные режимы, как неразделимые сочетания факторов, определяют 70 % дисперсии и добавляют к объяснению теплообеспеченностью еще 13 %.
Обсуждение
Биомное (по растительности и животному населению) экспертно-умозрительное деление Палеарктики (Udvardy, 1975; Воронов, Кучерук, 1976) на общей части территории заметно отличается от выполненного нами строго формализованного биотического (флоро-фаунистического) районирования. Если не принимать во внимание частности в местах проведения границ, различия сводятся к следующему. Указанные выше авторы, в отличие от нас, не выделяют полярные пустыни, включая их полностью в тундровый биом. То же следует сказать о предтундровых редколесьях, отнесенных частично в тундровый или таежный биомы. Территория выделенных ими отдельно летнезеленых широколиственных лесов и степной биом входят в состав Таежно-степной подобласти биотического деления. Остальные биомы с несколько иными границами и названиями примерно совпадают с выделенными нами биотическими провинциями. Таким образом, можно считать, что принципиальные отличия прослеживаются в основном в арктической и субарктической части Северной Евразии и в меньшей степени в юго-западной части Таежно-степной провинции флоро-фаунистического деления. Степной биом при флоро-фаунистическом районировании разделен примерно пополам. Его северная часть (луговые степи) входит в Таежно-степную подобласть, а южная (сухие или настоящие степи) вместе с полупустынями – в Пустынно-степную.
Биотическое районирование этой части Палеарктики в 1.6 раза полнее аппроксимирует матрицу флоро-фаунистической неоднородности, чем биомное разделение. Последнее ближе к зональным представлениям, но, скорее всего, потому, что оно в значительной степени задано авторами, а не выявлено ими в процессе анализа. Результаты фаунистического районирования по позвоночным животным на уровне регионов и флоро-фаунистических подобластей в целом весьма сходны, если не принимать во внимание локальные несовпадения в местах проведения границ. Значимые отличия сводятся к следующему. Предтундрово-редколесная флоро-фаунистическая подобласть разделена по фауне позвоночных между Субарктическим тундровым и Таежно-степным регионами, а Тундрово-таежная притихоокеанская подобласть объединена с последним из них, который включает также часть южнее расположенных полупустынь и северных пустынь, а не только лесостепь и северные степи. Среднеазиатская часть южных пустынь занимает меньшую площадь, образуя по фауне отдельные регион и подобласть. Сихотэ-Алиньский и Сахалино-Курильский фаунистические округа при флоро-фаунистическом делении образуют самостоятельную подобласть так же, как крымско-кавказские округа.
Таким образом, флоро-фаунистическое деление отличается от фаунистического в основном рангом и представительностью ряда таксонов, а также принадлежностью предтундрово-редколесных территорий к двум соседним фаунистическим подобластям и регионам. То есть они не образуют единую подобласть, как при флоро-фаунистическом районировании. Это дает основание утверждать, что дифференциация указанной территории по флоро-фаунистической неоднородности выше, чем по позвоночным, и четче совпадает с зональным делением.
По фауне короедов сходство с результатами флоро-фаунистического районирования очень велико. Отличия в общем сводятся к объединению по короедам в отдельную подобласть территории Тундрово-таежной притихоокеанской флоро-фаунистической подобласти с приохотской частью редколесий, а также отделению фауны Курильских островов от сахалинской и уссурийской из-за большей близости их к таковой на Камчатке (Курильско-Камчатско-Охотская подобласть). При разделении территории Северной Евразии по остальным видам жуков как части эталонной группы беспозвоночных эти территории, кроме Уссурийского края и южной части Сахалина, входят в Балтийско-Тихоокеанскую лесную подобласть. Кроме того, по беспозвоночным Крымско-Кавказская подобласть занимает бóльшую, чем совместно по флоре и фауне, площадь за счет включения в ее состав южного Причерноморья, Крыма и Предкавказья. Итак, результаты районирования по выбранной группе жуков ближе к флоро-фаунистическому делению, чем к выполненному по позвоночным.
Результаты районирования по видам древесных растений на уровне региона и подобласти отличаются от флоро-фаунистического разделения очень сильно. Так, Полярно-пустынно-тундровый регион объединяет две подобласти, в первую из которых входит Земля Франца Иосифа, где, по данным портала Биодат, древесных растений нет. Вторая включает все остальные острова Ледовитого океана и большую часть тундровых и редколесных материковых участков. Третий регион образуют остальные редколесья, а также таежные и отчасти лесостепные и степные участки. К этому же региону отнесены все притихоокеанские территории, включая Сахалин и Курилы, которые по флоро-фаунистическому делению выделены как самостоятельные подобласти и не включают северо-восточные тундры и приохотскую материковую часть. На западе, напротив, лесные, степные и полупустынные участки входят в лесопустынно-степную подобласть Юго-Западного региона. Территория Кавказа, пустынно-горно-степные участки Средней Азии, так же как Приморье с Курилами, составляют отдельные подобласти. Главным отличием разделения территории по флоре древесных растений от комплексного флоро-фаунистического можно считать отделение в самостоятельную подобласть юго-западной части, вместе с территорией настоящих степей, полупустынь и северных пустынь.
В итоге можно говорить о вариабельности (подвижности) границ на стыке крупных регионов, подобластей и провинций в зависимости от состава анализируемой группы растений и животных. Объединение всех проанализированных выборок приводит к более четкому по зональности выделению биотических таксонов, но не снимает необходимости дальнейших работ в этом направлении. При этом возможны два варианта. Первый, наиболее желательный, сводится к следующему: дальнейшее дополнение групп уже не будет значимо менять результаты районирования. Второй сведется к продолжению различий в прохождении границ. В этом случае аналогичные работы следует продолжать, пока существенная вариабельность границ не прекратится.
В целом предлагаемое нами районирование в большей степени объясняет неоднородность усредненной матрицы флоро-фаунистического сходства, чем результаты климатического, биогеографического, флористического и особенно физико-географического и лесорастительного районирования соответственно в 1.5–1.9 и 2.3–2.9 раза (в среднем вдвое). Общий множественный коэффициент корреляции всей системы наших объяснений равен примерно 0.91.
Заключение
Результаты проведенного кластерного анализа флоро-фаунистической неоднородности Северной Евразии при сделанных допущениях в общем сходны с ранее полученными представлениями об изменчивости флоры и фауны этой территории. Однако использование методов непараметрической статистики позволило подтвердить некоторые утверждения по флористическому и фаунистическому разделению указанной территории, отвергнуть другие из них или показать невысокую информативность ряда традиционных представлений, например, о высокой значимости Урала и Енисея как рубежей в неоднородности флоры и фауны. Кроме того, состав подобластей по числу участков, как правило, весьма сходен с зональным (от 45 до 100 % в каждом из выделенных таксонов), хотя никогда не совпадает со списком их в целом по зоне. Высокой сборностью в этом плане отличаются Таежно-степная, Крымско-Кавказская и Казахстанско-Среднеазиатская подобласти, поскольку 23–47 % участков в них относятся к соседним зонам. Относительно новым можно считать и выявление диагональных смещений под разным углом по отношению к границам природных зон в результате отепляющего влияния Северо-Атлантического течения и, в меньшей степени, Тихого океана. Главная новизна, как нам представляется, заключается в количественных оценках связи между флоро-фаунистической неоднородностью и изменчивостью основных факторов среды и их сочетаний (природно-географических режимов), как по отдельности, так и по суммарной оценке информативности представлений, рассчитанной через учитываемую ими дисперсию коэффициентов сходства комплекса флор и фаун. При этом не только показано преобладание влияния зональности по сравнению с провинциальностью (континентальностью), но и оценено их интегральное влияние через изменения теплообеспеченности территорий. Поэтому большее совпадение выявлено не с физико-географическим, а с климатическим районированием (соответственно 27 и 42 % дисперсии).
Библиография
Атлас СССР . М.: ГУГК, 1983. 260 с.
Блинова Т. К., Равкин Ю. С. Орнитофаунистическое районирование Северной Евразии // Сиб. экол. журн. 2008. Т. 15. № 1. С. 101–121.
Бобров В. В., Алещенко Г. М. Схема герпетогеографического районирования России и сопредельных стран // Вопросы герпетологии. Пущино; Москва, 2001. С. 31–34.
Воронов А. Г., Кучерук В. В. Биотическое разнообразие Палеарктики: проблемы изучения и охраны // Биосферные заповедники: Труды I советско-американского симпозиума, СССР, 5–17 мая 1976 г. Л.: Гидрометеоиздат, 1977. С. 7–20.
Кузнецов Б. А. Очерк зоогеографического районирования СССР . М.: Изд-во Моск. об-ва испытателей природы, 1950. 176 с.
Курнаев С. Ф. Лесорастительное районирование СССР . М.: Наука, 1973. 203 с.
Малышев Л. И. Моделирование флористического районирования кластерным анализом элементарных выделов Северной Азии и Европы // Сравнительная флористика на рубеже III тысячелетия: достижения, проблемы, перспективы: Материалы V рабочего совещания по сравнительной флористике. СПб., 2000. С. 20–36.
Мекаев Ю. А. Зоогеографические комплексы Евразии . Л.: Наука. Ленингр. отд-ние, 1987. 125 с.
Равкин Ю. С., Богомолова И. Н., Цыбулин С. М. Фаунистическое районирование Северной Евразии // Изв. РАН. Сер. геогр. 2015а. № 3. С. 29–40.
Равкин Ю. С., Богомолова И. Н., Цыбулин С. М. Сравнительное районирование Северной Евразии по древесным растениям и наземным позвоночным // Сибирский лесной журнал. 2015б. № 5. С. 42–53.
Равкин Ю. С., Богомолова И. Н., Цыбулин С. М., Легалов А. А. Районирование Северной Евразии по фауне короедов (Scolytidae) // Евразиатский энтомологический журнал. 2014. Т. 13. Вып. 3. С. 271–279.
Равкин Ю. С., Куперштох В. Л., Трофимов В. А. Пространственная организация населения птиц // Птицы лесной зоны Приобья / Ю. С. Равкин. Новосибирск: Наука. Сиб. отд-ние, 1978. С. 253–269.
Равкин Ю. С., Ливанов С. Г. Факторная зоогеография . Новосибирск: Наука. Сиб. отд-ние, 2008. 205 с.
Равкин Ю. С., Седельников В. П., Сергеев М. Г., Титлянова А. А., Хмелев В. А., Богомолова И. Н., Цыбулин С. М. Пространственно-типологическая дифференциация экосистем Западно-Сибирской равнины. Сообщение V. Экосистемы суши // Сиб. экол. журн. 2011. Т. 18. № 6. С. 773–788.
Рихтер Г. Д. Физико-географическое районирование СССР // Физико-географический атлас мира. М.: Изд-во АН СССР и Главного управления геодезии и картографии СССР, 1964. С. 248–249.
Северцов Н. А. О зоологических (преимущественно орнитологических) областях внетропических частей нашего материка // Изв. Русск. геогр. об-ва. СПб., 1877. Т. 13. Вып. 3. С. 125–155.
Семенов-Тян-Шанский А. Пределы и зоогеографические подразделения Палеарктической области для наземных сухопутных животных на основании географического распределения жесткокрылых насекомых . М.; Л.: Изд-во АН СССР, 1936. 16 c.
Трофимов В. А. Модели и методы качественного и факторного анализа матрицы связи // Проблемы анализа дискретной информации. Ч. II. Новосибирск, 1976. С. 24–36.
Трофимов В. А. Качественный факторный анализ матриц связей в пространстве разбиений со структурой // Модели агрегирования социально-экономической информации. Новосибирск: Наука, 1978. С. 91–106.
Трофимов В. А., Равкин Ю. С. Экспресс-метод оценки связи пространственной неоднородности животного населения и факторов среды // Количественные методы в экологии. Л., 1980. С. 113–115.
Флора СССР . М.; Л., 1934. Т. 1. 302 с.
Челинцев Н. Г. Математические основы учета животных . М.: ГУ Центроохотконтроль, 2000. 431 с.
Ravkin Yu. S., Bogomolova I. N., Tsybulin S. M., Legalov A. A. Zoning of Northern Eurasia based on the fauna of pine flower, fungus and leaf-rolling weevils (Coleoptera: Nemonychidae, Anthribidae, Rhynchitidae, Attelabidae) // Евразийский энтомологический журнал. 2015. № 14 (4). С. 367–373.
Holt Ben G., Lessard Jean-Phitippe, Borregaard Michael K., Fritz Susanne A., Araújo Miguel B., Dimitrov Dimitar, Fabre Pierre-Henri, Graham Catherine H., Graves Gary R., Jensson Knud A., Nogués-Bravo David, Wang Zhiheng, Whittaker Robert J., Fjeldsá Jon, Rahbek Carsten. An Update of Wallace’s Zoogeographic Regions of the World // Science. 2013. Vol. 339. № 4. P. 74–79.
Udvardy M. D. F. Classification of the Biogeographic Provinces of the World Occasional № 18 Int. Union for Conservation of Nature. Morges. Swittzerland, 1975. P. 1–48.
Благодарности
Исследования, послужившие основой для настоящей статьи, выполнены по программе ФНИ государственных академий наук на 2013–2020 гг., проект № VI.51.1.8, и частично в рамках «Программы повышения конкурентоспособности Томского государственного университета».

 © 2011 - 2026
© 2011 - 2026