



Введение
Хлорорганические соединения относятся к числу широко распространенных загрязнителей экосистем. Многие из них химически стабильны, токсичны, в силу чего накапливаются и длительное время сохраняются в окружающей среде, оказывая на нее негативное воздействие. К числу таких устойчивых хлорорганических соединений с повышенной токсичностью и широким спектром действия относятся отравляющее вещество иприт – 2,2′-дихлорэтилсульфид и продукты его гидролиза (ПГИ), основным из которых является тиодигликоль – 2,2′-тиодиэтанол.
Иприт – отравляющее вещество кожно-нарывного действия, обладает многосторонним поражающим действием на все живые организмы, и прежде всего на макроорганизмы. Под действием иприта наблюдаются значительные патологические изменения в дыхательных путях, желудочно-кишечном тракте, поражения органов зрения и др. При попадании иприта в организм развиваются симптомы общей интоксикации, свидетельствующие об универсальном характере повреждающего действия иприта. Ведущими являются изменения со стороны системы крови, нервной системы, сердечно-сосудистой системы и обмена веществ. Кроме того, выявлено мутагенное, канцерогенное и тератогенное действие иприта (Александров, Емельянов, 1990; Debouzy et al., 2002; Куценко и др., 2004).
Загрязнение почв и водоемов ипритом происходило как в процессе его хранения, так и при транспортировке к местам уничтожения, при аварийных ситуациях и др. Кроме того, со времен Второй мировой войны вследствие затопления химического оружия ипритом загрязнены многие водные экосистемы, например Балтийское и Адриатическое моря, прибрежные воды Японии, США, Великобритании, Австралии (Plunkett, 2003; REDCOD, 2006; Andrulewicz, 2007; Medvedeva et al., 2009). Так, в Балтийском море в 1947–1951 гг. были затоплены десятки тысяч тонн немецкого трофейного оружия, содержащего иприт, люизит, фосген и другие вещества (Chemiche Kampftoffamunition, 1993).
В водной среде иприт подвергается гидролизу (рис. 1). Процесс гидролиза происходит медленно, т. к. лимитируется низкой растворимостью иприта.
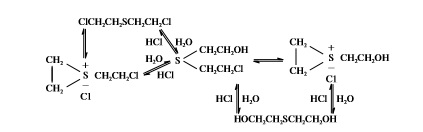
Рис. 1. Схема гидролиза иприта
Fig. 1. Scheme of the mustard gas hydrolysis
Скорость реакции гидролиза иприта замедляется в присутствии ионов хлора, поэтому в морской воде скорость гидролиза снижается в 2.5–4 раза по сравнению с пресной водой (Rosenblatt et al., 1975, 1996; Small, 1984). В среде, содержащей галогены, гидролиз иприта идет с образованием продуктов, не только препятствующих его дальнейшему растворению, но и вступающих во взаимодействие с ипритом и между собой, образуя ряд токсичных сульфониевых соединений, в несколько раз превышающих по токсичности сам иприт (Александров, Емельянов, 1990).
Попадая в окружающую среду, иприт и ПГИ оказывают негативное влияние на гидробионтов.
Введение в воду спиртово-водного раствора иприта в концентрациях от 1.73 до 26.2 мг/л приводило к гибели 50 % икры тритонов, жабы зеленой и лягушки озерной (Завьялов, 1995). Китайские ученые установили, что контаминация морской воды ипритом в концентрации 50–750 мг/л за 1 час приводит к гибели исследованных рыб и крабов, а цвет представителей зеленых и бурых водорослей изменялся через несколько минут после контакта с ипритом (Zhu, 1985).
Исследования по влиянию ПГИ на гидробионтов показали, что 50 % гибель моллюсков наступает при содержании ксенобиотиков 4.42–4.47 г/л за 96–102 часа. Аналогичный эффект достигается за 80–92 часа при воздействии ПГИ в концентрациях 1.81 и 4.4 г/л на земноводных – лягушку озерную и жабу зеленую соответственно. Наибольшая чувствительность отмечена у рака речного – 50 % гибель особей наблюдалась менее чем через 2 часа при концентрации 0.2 г ПГИ/л (Конешов, 1999).
Иприт и продукты его гидролиза вызывают морфологические и ультраструктурные изменения в клетках микроорганизмов, увеличивают проницаемость их клеточных оболочек, а также, обладая повреждающим действием на дезоксирибонуклеиновые кислоты (ДНК), оказывают негативное действие на генетический аппарат (Ichinotsubo et al., 1977; Kircher, Brendel, 1983; Зайцева, 2000; Кузикова и др., 2007; Орлова, 2007).
Загрязнение морской воды ипритом и его производными вызывает существенные изменения бактериоценозов, выражающиеся в увеличении показателей доминирования видов, снижении показателей сходства, видового разнообразия и выравненности бактериального сообщества. Снижение уровня видового разнообразия морского бактериопланктона свидетельствует о снижении устойчивости биосистемы в целом (Medvedeva et al., 2009; Медведева и др., 2012).
Продукты гидролиза иприта оказывают ингибирующее действие на рост водных микроорганизмов. При этом представители фитопланктона – цианобактерии – более чувствительны к продуктам гидролиза иприта по сравнению с бактериальными культурами (Медведева и др., 2016).
Однако информация о влиянии ПГИ на физиолого-биохимические свойства цианобактерий, являющихся первичными продуцентами водных экосистем и обладающих уникальными способностями противостоять различного рода стрессовым воздействиям как природного, так и антропогенного происхождения, в доступной литературе отсутствует.
Целью настоящей работы являлось исследование влияния продуктов гидролиза иприта на рост, фотосинтетическую активность и синтез вторичных метаболитов цианобактериями – основными возбудителями цветения воды – Aphanizomenon flos-aquae, Microcystis aeruginosa, Nodularia spumigena, Trichormus (Anabaena) variabilis.
Материалы
Исследования проводились с использованием культур цианобактерий Aphanizomenon flos-aquae CALU 1033, Microcystis aeruginosa CALU 973, Nodularia spumigena CALU 795, Trichormus (Anabaena) variabilis CALU 458, полученных из коллекции Биологического института СПбГУ (Россия).
Методы
Цианобактерии культивировали в жидкой среде BG-11 (Rippka et al., 1979), содержащей ПГИ в количестве (по хлорорганическим соединениям (ХОС)) от 0.3 до 20 мг/л. Культуры выращивали в течение 14 суток в колбах вместимостью 250 мл в стационарных условиях при температуре 22–24 °С, освещенности 1000 лк и световом режиме свет : темнота – 12 : 12. Объем питательной среды в колбе составлял 100 мл. В качестве посевного материала использовали культуры в экспоненциальной фазе роста, выращенные в тех же условиях.
Рост цианобактерий контролировали по сухому весу. Содержание хлорофилла а определяли по оптической плотности характерных полос поглощения на спектрофотометре Genesys 10UV scanning (Thermo Spectronic, США). Экстракцию хлорофилла а проводили 90 % ацетоном при 4 °С в течение 24 часов. Концентрацию хлорофилла а рассчитывали по формуле Джеффри и Хамфри (Jeffrey, Humprhray, 1975).
Модельную смесь ПГИ получали нагреванием водной смеси 0.65 М тиодигликоля (ТДГ) (ICN 103039 RT, 99 %) и 0.65 M HCl при 90 °C в течение 8 часов. Определение содержания ХОС и ТДГ проводили по описанным ранее методикам (Medvedeva et al., 2008). Модельная смесь ПГИ содержала 3.0 г/л ХОС и 66.0 г/л ТДГ.
При определении содержания микроцистина-LR в биомассе культуры Microcystis aeruginosa клетки отделяли от нативного раствора центрифугированием при 6 000 об./мин в течение 10 мин. Микроцистин-LR из клеток экстрагировали 50 % метанолом (Lawton et al., 1994) после многократного замораживания-размораживания биомассы. Для определения содержания внеклеточных микроцистинов супернатант лиофильно высушивали, осадок растворяли в 25 % растворе метанола.
Концентрацию микроцистина-LR в метанольных экстрактах определяли методом ВЭЖХ на хроматографе «Hewlett-Packard» HP1090 с диодно-матричным детектором (длина волны 238 нм, разрешение 2 нм). Условия хроматографирования: колонка Luna фирмы Phenomenex, температура – 35 °C, скорость потока – 1.5 мл/мин, детекция – 215 нм, элюент – 30 % ацетонитрил (растворитель А) и 50 % ацетонитрил (растворитель Б), подкисленные 0.1 % раствором трифторуксусной кислоты, объем пробы – 0.2 мл. Стандартные растворы микроцистина-LR были получены от Alexis Corporation (Lausen, Швейцария).
Количество водорастворимых экзополисахаридов определяли по стандартной методике (Herbert et al., 1971), содержание полисахаридов выражали в миллиграммах глюкозы на 1 г а.с.б. и миллиграммах глюкозы на 1 л.
В качестве основных токсикологических параметров использовали NOEC – наиболее высокие концентрации токсиканта, при которых не наблюдался статистически значимый ингибирующий эффект (p < 0.05), EC50 – концентрации токсиканта, вызывающие 50 % ингибирование роста цианобактерий или образования хлорофилла а. Токсикологические параметры рассчитывали методом нелинейной регрессии (Nyholm et al., 1992).
Эксперименты проводились в сериях из трех опытов с трехкратными повторностями. Статистическую обработку результатов проводили с использованием пакета компьютерных программ Statistica 6.0 (Stat Soft).
Результаты
Ингибирующее действие продуктов гидролиза иприта на рост цианобактерий проявлялось в диапазоне концентраций, превышающих 2.0–6.0 мг ХОС/л в зависимости от культуры. Подавление роста на 50 % через 14 суток культивирования наблюдалось при содержании в среде ПГИ в количестве от 5.5 мг ХОС/л (Microcystis аeruginosa) до 11.2 мг ХОС/л (Aphаnizomenon flos-aquae) (табл. 1).
Таблица 1. Параметры токсичности продуктов гидролиза иприта для цианобактерий
| Культура | Рост | Содержание хлорофилла a | ||
| NOEC | EC50 | NOEC | EC50 | |
| Aphаnizomenon flos-aquae, 1033 | 6.0 ± 0.3* | 11.2 ± 0.6 | 2.0 ± 0.1 | 6.2 ± 0.3 |
| 132 ± 7 | 246 ± 24 | 44 ± 2 | 136 ± 7 | |
| Microcystis аeruginosa, 973 | 2.0 ± 0.1 | 5.5 ± 0.3 | 1.1 ± 0.1 | 2.5 ± 0.1 |
| 44 ± 2 | 121 ± 7 | 24 ± 2 | 55± 3 | |
| Nodularia spumigena, 795 | 3.1 ± 0.2 | 6.1 ± 0.3 | 1.5 ± 0.1 | 2.5 ± 0.1 |
| 68 ± 5 | 134 ± 13 | 33 ± 2 | 55± 3 | |
| Trichormus variabilis, 458 | 5.2 ± 0.3 | 7.9 ± 0.4 | 4.5 ± 0.2 | 7.5 ± 0.4 |
| 114 ± 7 | 174 ± 9 | 99 ± 5 | 165 ± 8 | |
Примечание. * – в числителе – ХОС, мг/л, в знаменателе – ТДГ, мг/л.
В условиях стресса, вызванного действием ПГИ, у цианобактерий наряду с ингибированием роста снижалось содержание хлорофилла a (см. табл. 1).
Следует отметить, что процесс образования хлорофилла а цианобактериями Aphаnizomenon flos-aquae, Nodularia spumigena, Microcystis аeruginosa отличался большей чувствительностью к воздействию смеси ПГИ, чем рост культур. Концентрации, соответствующие NOEC и EC50 для хлорофилла а, были ниже таковых для роста в 1.8–3 и 1.8–2.4 раза соответственно (см. табл. 1). В случае же цианобактерии Trichormus variabilis не выявлено существенных различий в токсичности смеси ПГИ для роста и образования хлорофилла а.
Ингибирование роста цианобактерий в присутствии ПГИ сопровождалось образованием и выделением в среду повышенных количеств полисахаридов.
Продуктивность биомассы цианобактерий по экзополисахаридам (мг глю/г а.с.б.) и концентрация экзополисахаридов в среде (мг глю/л) в условиях ингибирования роста на 50 % возрастали на 148–180 % и 27-36 % соответственно по сравнению с контрольными значениями в зависимости от вида цианобактерий (табл. 2).
Таблица 2. Влияние продуктов гидролиза иприта на синтез экзополисахаридов цианобактериями
| Культура | Концентрация ПГИ, ХОС, мг/л
ТДГ, мг/л |
Выход биомассы, г/л | Концентрация экзополисахаридов | |
| мг глю/г а.с.б. | мг глю/л | |||
| Aphаnizomenon
flos-aquae |
Контроль | 0.25 ± 0.03 | 530.0 ± 55.1 | 132.5 ± 14.1 |
| 11.2 246 | 0.13 ± 0.01 | 1313.8 ± 132.2 | 170.8 ± 15.9 | |
| Microcystis aeruginosa | Контроль | 0.11 ± 0.01 | 20.0 ± 2.2 | 2.2 ± 0.2 |
| 5.5 121 | 0.05 ± 0.01 | 56.0 ± 5.8 | 2.8 ± 0.3 | |
| Trichormus variabilis | Контроль | 0.16 ± 0.02 | 142.5 ± 15.3 | 22.8 ± 2.4 |
| 7.9 174 | 0.08 ± 0.01 | 387.5 ± 40.3 | 31.0 ± 3.3 | |
Продукты гидролиза иприта оказывали влияние не только на синтез хлорофилла а и экзополисахаридов, но и на синтез и выделение в среду таких важнейших для водных экосистем метаболитов, как цианотоксины.
В условиях 50 % подавления роста культуры Microcystis aeruginosa 973 наблюдалось ингибирование токсинообразования единицей биомассы на 68 % и, как следствие, снижение содержания в среде микроцистина-LR на 84 % (рис. 2).
Однако в условиях стимулирования роста токсигенной цианобактерии Microcystis aeruginosa 973 низкими концентрациями ПГИ выявлено увеличение содержания микроцистина-LR в среде на 39 %, что связано, главным образом, с образованием повышенного количества биомассы цианобактерий (см. рис. 2).
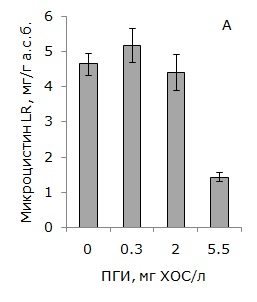
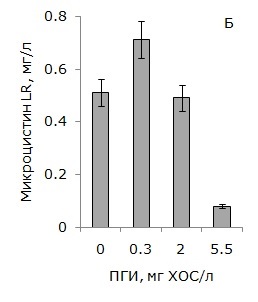
Рис. 2. Влияние ПГИ на продуктивность биомассы Microcystis aeruginosa 973 по микроцистину-LR (А) и содержание микроцистина-LR в среде (Б)
Fig. 2. Effect of mustard gas hydrolysis products on microcystin LR production on M. aeruginosa 973 (A) and microcystin LR concentrations in the medium (Б)
Обсуждение
Продукты гидролиза иприта оказывают влияние как на рост цианобактерий, так и на их физиологическое состояние.
Ингибирующее действие продуктов гидролиза иприта на рост цианобактерий проявляется в диапазоне концентраций, превышающих 2.0–6.0 мг ХОС/л в зависимости от культуры. Наиболее устойчивой к воздействию ПГИ является цианобактерия Aphаnizomenon flos-aquae (ЕС50 – 11.2 мг ХОС/л, 246 мг ТДГ/л). Наибольшую чувствительность к токсиканту проявила токсигенная цианобактерия Microcystis aeruginosa 973, 50 % ингибирование роста которой наблюдалось при содержании продуктов гидролиза иприта в среде 5.5 мг ХОС/л.
В условиях стресса, вызванного действием ПГИ, у всех исследованных культур цианобактерий наблюдалось ингибирование синтеза хлорофилла а. Степень ингибирования зависела от вида цианобактерии. Следует отметить различную чувствительность пигментного аппарата у исследованных культур цианобактерий к действию ПГИ. По степени чувствительности образования хлорофилла а к ПГИ цианобактерии можно расположить в следующем порядке: Microcystis aeruginosa > Nodularia spumigena > Aphаnizomenon flos-aquae > Trichormus variabilis. Различная чувствительность синтеза хлорофилла а в зависимости от вида цианобактерий ранее наблюдалась и в отношении других поллютантов как неорганической, так и органической природы (Poliak et al., 2011; Zaytseva et al., 2015).
Ингибирование роста цианобактерий сопровождалось увеличением синтеза и экскреции в среду экзополисахаридов. Повышенный синтез экзополисахаридов является одним из возможных механизмов защиты цианобактерий от негативного воздействия продуктов гидролиза иприта.
Полученные результаты согласуются с представленными нами ранее результатами и данными других исследователей по усилению синтеза экзополисахаридов под действием неблагоприятных факторов, в том числе токсикантов. Изменения в продукции экзополисахаридов цианобактериями отмечены под действием различных токсикантов как неорганической, так и органической природы, причем эффект зависел как от природы токсиканта и его концентрации, так и от культуры водорослей (Otero, Vincenzini, 2003; Poliak et al., 2011; Zhao et al., 2011; Deng et al., 2012; El-Sheekh et al., 2012; Medvedeva et al., 2017).
Увеличение синтеза внеклеточных полисахаридов под действием ПГИ отмечалось ранее и у представителей другой таксономической группы микроорганизмов – микромицетов (Кузикова и др., 2007; Орлова, 2007).
Повышенный синтез микробных полисахаридов существенным образом влияет на формирование качества воды в водоемах, т. к. они влияют на углеродный цикл и микробное разнообразие, служат субстратом для питания других микроорганизмов, развитие которых, с одной стороны, важно для процессов самоочищения, с другой – усиливает биологическое и химическое загрязнение воды за счет клеток микроорганизмов и их метаболитов (El-Sheekh et al., 2012).
Продукты гидролиза иприта оказывают влияние не только на синтез хлорофилла а и экзополисахаридов, но и на синтез токсигенным штаммом Microcystis aeruginosa 973 высокотоксичного микроцистина-LR и его содержание в среде.
Следует отметить, что в низкой концентрации 0.3 мг/л (по ХОС) продукты гидролиза иприта оказывали стимулирующее действие на рост токсигенного штамма Microcystis аeruginosa 973. В присутствии низких доз ПГИ прирост биомассы цианобактерии превышал контрольные значения на 25 % (Медведева и др., 2016). В условиях стимулирования роста цианобактерии Microcystis aeruginosa 973 продуктивность биомассы по микроцистину-LR оставалась на уровне контрольных значений, однако общее количество токсина в единице объема культуральной жидкости возрастало на 39 %. С увеличением концентрации ПГИ в среде (до 5.5 мг ХОС/л) происходило ингибирование роста культуры на 50 %, снижение продуктивности биомассы по микроцистину-LR на 68 % и уменьшение его концентрации в среде на 84 %.
Изменение синтеза микроцистинов цианобактериями р. Microcystis ранее было отмечено под влиянием условий культивирования, наличия в среде биогенных элементов, поллютантов органической и неорганической природы (Oh et al., 2000; Wang et al., 2007; Волошко и др., 2008; Poliak et al., 2011; Polyak et al., 2013; Zaytseva et al., 2015).
Заключение
Иприт и продукты его гидролиза оказывают существенное влияние на массовые виды цианобактерий.
В ходе проведенных исследований выявлено, что продукты гидролиза иприта оказывают токсическое действие на рост и синтез хлорофилла а всеми исследованными культурами цианобактерий.
Токсигенный штамм цианобактерий Microcystis aeruginosa 973 в случае стимулирования роста низкими концентрациями ПГИ накапливает в среде повышенное количество микроцистина-LR. Ингибирование роста Microcystis aeruginosa сопровождалось снижением синтеза и экскреции токсина в среду.
В условиях стресса, вызванного воздействием на цианобактерии продуктов гидролиза иприта, происходит повышенный синтез и выделение в среду таких протекторных соединений, как полисахариды.
Повышенное выделение таких метаболитов, как полисахариды и цианотоксины, является серьезным негативным последствием загрязнения водных объектов продуктами гидролиза иприта.
Библиография
Александров В. Н., Емельянов В. И. Отравляющие вещества . М.: Военное изд-во, 1990. 271 с.
Волошко Л. Н., Пиневич А. В., Копецкий И., Титова Н. Н., Хроузек П., Зелик П. Продуцируемые цианобактериями токсины в период «цветения» воды в нижнем Cуздальском озере (Санкт-Петербург, Россия) ⁄⁄ Альгология. 2008. Т. 20. № 2. С. 210–223.
Завьялов Е. В. Эколого-токсикологическое воздействие кожно-резорбтивных отравляющих веществ на фауну: Автореф. дис. … канд. биол. наук . Волгоград, 1995. 18 c.
Зайцева Т. Б. Микробиологическая деструкция иприта и продуктов его гидролиза: Дис. … канд. биол. наук . СПб., 2000. 129 с.
Конешов С. А. Эколого-гидробиологическая диагностика токсикантов в зонах экологического риска: Автореф. дис. … канд. биол. наук . Саратов, 1999. 18 с.
Кузикова И. Л., Медведева Н. Г., Сухаревич В. И., Орлова О. Г., Рыбальченко О. В. Влияние продуктов гидролиза иприта на микромицеты // Микология и фитопатология. 2007. Т. 41. Вып. 3. С. 252–260.
Куценко С. А., Бутомо Н. В., Гребенюк А. Н., Ивницкий Ю. Ю., Мельничук В. П., Преображенская Т. Н., Рыбалко В. М., Саватеев Н. В. Военная токсикология, радиобиология и медицинская защита / Под ред. С. А. Куценко. СПб.: ООО «Издательство Фолиант», 2004. 526 с.
Медведева Н. Г., Поляк Ю. М., Зайцева Т. Б., Жариков Г. А. Деструкция продуктов гидролиза иприта морскими и почвенными бактериями // Известия РАН. Серия биологическая. 2012. № 1. С. 91–99.
Медведева Н. Г., Зайцева Т. Б., Кузикова И. Л., Зиновьева С. В. Оценка токсичности продуктов гидролиза иприта для водных микроорганизмов // Вода: химия и экология. 2016. № 1. С. 76–81.
Орлова О. Г. Влияние продуктов гидролиза иприта на микроорганизмы: Дис. … канд. биол. наук . СПб., 2007. 115 с.
Andrulewicz E. Chemical weapons dumped in the Baltic Sea / I. E. Gonenc et al. (eds.). Assessment of the fate and effect of toxic agents in water. Springer, 2007. P. 301–321.
Chemiche Kampftoffamunition in der sudlichen und westlichen Ostsee Herausgegeben vom Bundesamt fur Seeschiffahrtung Hydrographie. Hamburg, 1993. 66 s.
Debouzy J. C., Aous S., Dabouis V., Neveux Y., Gentilhomme E. Phospholipid matrix as a target for sulfur mustard (HD): NMR study in model membrane systems // Cell Biol. Toxicol. 2002. Vol. 18. № 6. P. 397–408.
Deng X., Gao K., Sun J. Physiological and biochemical responses of Synechococcus sp. PCC7942 to Irgarol 1051 and diuron // Aquat. Toxicol. 2012. Vol. 122–123. P. 113–119.
El-Sheekh M. M., Khairy H. M., El Shenody R. A. Algal production of extra and intra-cellular polysaccharides as an adaptive response to the toxin crude extract of Microcystis aeruginosa // Iran. J. Environ. Health. Sci. Health. 2012. Vol. 9. № 1. Article number В 10.
Jeffrey S. W., Humprhray G. E. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton // Biochim. and Physiol. Pflanz. Bd. 1975. Vol. 167. № 2. P. 191–194.
Herbert D., Phipps P. J., Strange R. E. Chapter III. Chemical Analysis of Microbial Cells // Methods in Microbiology. 1971. Vol. 5. Part B. P. 209–344.
Ichinotsubo D., Mower H. F., Setliff J., Mandel M. Use of Rec-bacteria for testing of carcinogenic substances // Mutat. Res. 1977. Vol. 46. P. 53–62.
Kircher M., Brendel M. DNA alkylation by mustard gas in yeast Saccharomyces cerevisiae strains of different repair capacity // Chem.-Biol. Interact. 1983. Vol. 44. P. 27–39.
Lawton L. A., Edwards C., Codd G. A. Extraction and high-performance liquid chromatographic method for the determination of microcystins in raw and treated waters // Analyst. 1994. Vol. 119. № 7. P. 1525–1530.
Medvedeva N., Polyak Yu., Kuzikova I., Orlova O., Zharikov G. The effect of mustard gas on the biological activity of soil // Environ. Res. 2008. Vol. 6. № 3. P. 289–295.
Medvedeva N., Polyak Yu., Kankaanpää H., Zaytseva T. Microbial responses to mustard gas dumped in the Baltic Sea // Marine Environ. Res. 2009. Vol. 68. P. 71–81.
Medvedeva N., Zaytseva T., Kuzikova I. Cellular responses and bioremoval of nonylphenol by the bloom-forming cyanobacterium Planktothrix agardhii 1113 // J. Marine Syst. Available online 16 January 2017. In Press. URL: http://dx.doi.org/10.1016/j.jmarsys.2017.01.009.
Nyholm N., Sørensen P. S., Kusk K. O., Christensen E. R. Statistical treatment of data from microbial toxicity tests // Environ. Toxicol. Chem. 1992. Vol. 11. Issue 2. P. 157–167.
Oh H. M., Lee S. J., Jang M. H., Yoon B. D. Microcystin production by Microcystis aeruginosa in a phosphorus-limited chemostat // Appl. Environ. Microbiol. 2000. Vol. 66. P. 176–179.
Otero A., Vincenzini M. Extracellular polysaccharide synthesis by Nostoc strains as affected by N source and light intensity // J. Biotechnol. 2003. Vol. 102. № 2. P. 143–152.
Plunkett G. Chemical Warfare Agent Sea Dumping off Australia. Commonwealth of Australia, 2003. 44 p.
Poliak Yu. M., Zaytseva T. B., Petrova V. N., Medvedeva N. G. Development of mass cyanobacteria species under heavy metals pollution // Hydrobiol. J. 2011. Vol. 47. № 3. P. 75–90.
Polyak Y., Zaytseva T., Medvedeva N. Response of toxic cyanobacterium Microcystis aeruginosa to environmental pollution // Water Air Soil Pollut. 2013. Vol. 224. P. 1494.
REDCOD Project. Reseach of environmental damage caused by chemical ordnance dumped at sea. Contract № B4-3070/2003/368585/SUB/D.3. Final Scientific Report. ICRAM, 2006. 277 p.
Rippka R., Deruelles J., Waterbury J. B., Herdman M., Stanier R. Y. Genetic assignments, strain histories and properties of pure cultures of cyanobacteria // J. Gen. Microbiol. 1979. Vol. 111. P. 1–61.
Rosenblatt D. H., Miller T. A., Dacre J. C., et al. Problem definition studies on potential environmental pollutants. II. Physical, chemical, toxicological, and biological properties of 16 substances. Fort Detrick, MD: U.S. Army Medical Bioengineering Research Development Laboratory, 1975. 291 p.
Rosenblatt D. H., Small M. J., Kimmell T. A., et al. Background chemistry for warfare agents and decontamination processes in support of delisting waste streams at the U.S. Army Dugway Proving Ground, Utah. Environmental Assessment Division, 1996. 88 p.
Small M. J. Compounds formed from the chemical decontamination of HD, GB, and VX and their environmental fate. U.S. Army Research and Development Command. Frederick, Maryland, 1984. 175 p.
Wang J., Xie P., Guo N. Effects of nonylphenol on the growth and microcystin production of Microcystis strains // Environ. Res. 2007. Vol. 103. № 1. P. 70–78.
Zaytseva T. B., Medvedeva N. G., Mamontova V. N. Peculiarities of the effect of octyl- and nonylphenols on the growth and development of microalgae // Inland Water Biol. 2015. Vol. 8. № 4. P. 406–413.
Zhao L., Lu L., Li M., Xu Z., Zhu W. Effects of Ca and Mg levels on colony formation and EPS content of cultured Microcystis aeruginosa(Conference Paper) // Procedia Environ. Sci. 2011. Vol. 10. Part B. P. 1452–1458.
Zhu M. The influence of mustard gas and other chemical agents to the kinds oceanic livings // Acad. J. Second Mil. Med. Univ. 1985. Vol. 6. Issue 5. P. 332–335.

 © 2011 - 2026
© 2011 - 2026