



Введение
Характеристика размеров – не только неотъемлемая составляющая описания любого вида животных. Морфометрические показатели используют при выделении подвидов, сравнении популяций, определении возраста и пола, расчетах энергетических затрат на жизнеобеспечение и т. п. Нередко прежде всего на них опираются при отсутствии у вида диморфизма в окраске оперения и поведении.
Перевозчик Actitis hypoleucos (Linnaeus, 1758) – мономорфный представитель отряда Ржанкообразных (Charadriiformes) с обширным гнездовым ареалом, который простирается от Атлантического до Тихого океана, зимовочный – охватывает огромные пространства от Африки до Юго-восточной Азии (Гладков, 1951; Козлова, 1961). Птицы, гнездящиеся в Европе, в том числе на северо-западе России, зимуют в Африке (Штифель и др., 1985; Резвый и др., 1995; Wernham et al., 2002; Valkama et al., 2014).
Генетические исследования не выявили наличия у вида четко дифференцируемых географических популяций (Zink et al., 2008; Hung et al., 2013). Вместе с тем вероятное разделение зимовок птиц из восточных и западных частей ареала, а также достаточно высокая степень гнездового консерватизма (Holland, Yalden, 1991; Dougall et al., 2005; Хохлова, Лунина, 2016) создают предпосылки для формирования территориальных группировок, которые могут отличаться в том числе и биометрическими показателями. Однако надежных показателей, которые позволяли бы судить об их вариациях в пределах ареала, пока очень немного, поскольку в большинстве случаев при расчетах использовали либо малые выборки музейных экземпляров, либо данные измерений живых птиц, отловленных во время сезонных миграций на местах остановки и смешения особей из разных регионов.
В ходе многолетних исследований в восточном Приладожье были использованы методы отлова, исключающие возможность попадания в выборку мигрантов, что позволило получить представление о морфометрических показателях локальной гнездовой популяции перевозчиков южной части Карелии.
Методы
Работы проводили на стационаре Института биологии КарНЦ РАН «Маячино» (60º 46' с. ш., 32° 48' в. д.) в 1990–2007 гг. Ежедневный контроль птиц и поиск гнезд и выводков на 5 км участке побережья Ладожского озера начинали в середине мая и заканчивали в августе.
Изучены морфометрические показатели взрослых размножающихся перевозчиков. Птиц ловили и кольцевали после вылупления птенцов, чтобы не привлекать к кладкам внимание серых ворон Corvus corone и других разорителей гнезд, постоянно присутствующих на побережье. Это давало возможность сразу выявлять родителей, но затрудняло определение их пола из-за отсутствия у вида полового диморфизма не только в окраске оперения, но и в поведении в период насиживания и вождения выводков. Кроме того, к этому времени уже нельзя было определить пол и по форме клоакального выступа отловленных особей. Он был установлен лишь у части маркированных птиц, вернувшихся с зимовок, по их поведению в предбрачный период. Данный способ визуального определения пола считают наиболее надежным (Dougall et al., 2010), хотя и здесь трудно избежать ошибок из-за большой подвижности птиц во время брачных игр. Современный способ определения пола генетическим методом не применяли.
Для отловов использовали лучки, в центр которых помещали птенцов, в редких случаях – паутинные сети (Хохлова, Лунина, 2015а). В зависимости от окраски и состояния оперения, пойманных птиц относили к младшей (годовалые) или старшей возрастной группе (Meissner et al., 2015; Blasco-Zumeta, Heinze, 2016). Всего индивидуальным сочетанием цветных колец помечено 140 взрослых птиц, 45 % которых возвращались в течение 1–7 лет (Хохлова, Лунина, 2016). Если обе птицы в паре были визуально идентифицированы по индивидуальной маркировке, их повторно не ловили.
У пойманных птиц измеряли длину крыла, хвоста, цевки и клюва по стандартной методике (Виноградова и др., 1976). Кроме того, снимали промеры клюва от его кончика до уголка рта (клюв2) и до ноздри (клюв3). Измерения проводили с точностью до 0.5 мм, используя штангенциркуль или линейку с упором для крыла и хвоста. Все или большая часть размеров получены от 129 птиц (табл. 1), причем 36 особей, возвращавшихся после зимовки, были измерены по 2–4 раза в разные годы (всего 178 промеров для крыла). В табл. 2 приведены морфометрические характеристики вида, вычисленные с использованием как всех промеров, так и средних значений для особей, измеренных повторно.
Анализ данных и расчеты производили с использованием возможностей Access и Excel. При обработке применены обычные статистические методы (Ивантер, Коросов, 2011). Средние величины указаны с ошибкой средней.
Результаты
Перевозчик – самый массовый вид куликов на побережьях водоемов Карелии в период размножения. На контролируемом участке Ладожского озера его средняя численность колебалась по годам от 2.6 до 6.3 пар/км, достигая на каменистых пляжах 12 пар/км береговой линии. Прилет происходил в конце апреля – начале мая. Период откладки яиц растянут на 1.5 месяца: крайние даты начала кладок – 7.05–23.06, вылупления птенцов – 2.06–12.07 (Хохлова, Лунина, 2015б). Все местные маркированные птицы, как взрослые, так и молодые, покидали побережье к началу августа.
Результаты повторных измерений птиц, вернувшихся с зимовок, не всегда совпадали со значениями предшествующего сезона (см. табл. 1). Это расхождение было минимальным для цевки и клюва, тогда как длина крыла и хвоста у некоторых птиц значимо колебалась по годам, составив в среднем 1.3 мм для крыла и около 2 мм для хвоста (см. табл. 1), что позволяет предполагать, что наряду с возможными ошибками измерения оно могло быть обусловлено разной степенью обношенности перьев и межгодовыми колебаниями их длины из-за особенностей условий протекания предбрачной линьки.
Таблица 1. Изменения длины крыла и хвоста перевозчика, измеренных повторно в разные годы (без разделения по полу)
Table 1. Changes in the wing and tail length of the Common Sandpipers Actitis hypoleucos measured repeatedly in different years (sexes combined)
| Количество измерений (разные годы) | Число особей с данным диапазоном изменений длины крыла / хвоста, мм | Средняя величина изменений длины, M ± m | ||||||||
| 0 | 1 | 2 | 3 | 4 | 5 | 8 | всего | крыла | хвоста | |
| 2 | 8 / 4 | 13 / 10 | 1 / 5 | 2 / 4 | 1 / 1 | - / 1 | - / 1 | 25 / 26 | 1.0 ± 0.21 | 1.6 ± 0.25 |
| 3 | - / - | 4 / 2 | 4 / 2 | 1 / 2 | - / 1 | - / - | - / - | 9 / 7 | 1.7 ± 0.71 | 2.3 ± 0.45 |
| 4 | - / - | - / 1 | 1 / - | 1 / - | - | - / 1 | - / - | 2 / 2 | 2.5 ± 0.71 | 3.0 ± 2.83 |
| Всего | 8/4 | 17/13 | 6/7 | 4/6 | 1/2 | -/2 | -/1 | 36/35 | 1.3 ± 0.17 | 1.9 ± 0.23 |
При сравнении морфометрических характеристик, рассчитанных с использованием всех промеров и средних значений для особей, измеренных повторно, различия между ними оказались незначительными и статистически недостоверными для всех показателей.
Большинство отловленных птиц (95 %) имели длину крыла 108–118 мм, хвоста – 52–63, цевки – 23–27 мм, клюва – 22–27 мм, и лишь у отдельных особей размеры выходили за эти границы (см. табл. 2). Предельные значения хорошо согласуются с данными авторов, работавших с гнездовым населением вида как в Англии (Holland et al., 1982; Yalden, 2012), так и в Норвегии (Løfaldli, 1981).
Таблица 2. Морфометрические характеристики перевозчика A. hypoleucos, вычисленные с использованием всех промеров (I) и средних показателей для особей, измеренных повторно (II), без разделения по полу
Table 2. Morphometrics of the Common Sandpiper A. hypoleucos (sexes combined) calculated using all measurements (I) and averages for individuals measured repeatedly (II)
| Показатель | Длина, мм | ||
| lim | I. M ± m (N) | II. M ± m (N) | |
| Крыло | 104–120 | 112.8 ± 0.22 (178) | 112.7 ± 0.26 (129) |
| Хвост | 49–65 | 57.3 ± 0.22 (172) | 57.2 ± 0.25 (127) |
| Цевка | 21–29 | 24.9 ± 0.12 (119) | 24.9 ± 0.14 ( 85) |
| Клюв | 21–29 | 24.8 ± 0.12 (159) | 24.9 ± 0.14 (118) |
| Клюв2 | 24–32 | 28.2 ± 0.11 (159) | 28.3 ± 0.13 (119) |
| Клюв3 | 12–22 | 17.9 ± 0.11 (144) | 17.9 ± 0.12 (107) |
Кривая распределения птиц по наиболее популярному показателю – длине крыла – имеет два пика: основной (32 %) на значениях 111–112 мм и небольшой (15 %) на значениях 114 мм, что можно связать с присутствием в выборке птиц разного возраста и пола.
Годовалые птицы уступают в размерах птицам старших возрастных групп (табл. 3), по длине крыла различия статистически значимы (t = 2.33, p = 0.05). Тенденции к дальнейшему увеличению линейных показателей после 2-летнего возраста не выявлено.
Таблица 3. Морфометрические показатели перевозчиков A. hypoleucos разных возрастных групп (мм) без разделения по полу
Table 3. Morphometric characteristics of different age groups of the Common Sandpiper A. hypoleucos (in mm, sexes combined)
| Возраст | Крыло | Хвост | Цевка | ||||||
| N | lim | M ± m | N | lim | M ± m | N | lim | M ± m | |
| 1 год | 44 | 104–119 | 111.9 ± 0.47 | 41 | 50–62 | 57.2 ± 0.39 | 21 | 21–28 | 24,9 ± 0,31 |
| ≥2 года | 75 | 108–120 | 113.3 ± 0.37 | 76 | 50–64 | 57.4 ± 0.27 | 62 | 23–29 | 25,0 ± 0,10 |
| ?? | 59 | 107–119 | 113.0 ± 0.41 | 56 | 49–65 | 57.3 ± 0.43 | 23 | 21–27 | 24,4 ± 0,28 |
| Возраст | Клюв | Клюв2 | Клюв3 | ||||||
| N | lim | M ± m | N | lim | M ± m | N | lim | M ± m | |
| 1 год | 42 | 22–29 | 24.9 ± 0.25 | 42 | 24–32 | 28.2 ± 0.24 | 40 | 15–20 | 17.9 ± 0.19 |
| ≥2 года | 70 | 22–29 | 24.9 ± 0.18 | 69 | 25–32 | 28.3 ± 0.18 | 69 | 15–22 | 18.1 ± 0.17 |
| ?? | 47 | 21–28 | 24.6 ± 0.22 | 47 | 25–31 | 28.1 ± 0.20 | 36 | 12–20 | 17.6 ± 0.24 |
В течение сезона оперение птиц постепенно обнашивается, что приводит к сокращению длины крыла и хвоста. У гнездящихся птиц, отловленных до середины июня и после 25 июня, эта разница составила: для крыла – 1.5 мм (t = 2.59, p = 0.01), для хвоста – 0.6 мм (табл. 4). Кроме того, слегка увеличилась длина клюва по коньку из-за обноса оперения у его основания, тогда как длина цевки не изменилась.
Таблица 4. Морфометрические показатели перевозчиков A. hypoleucos, отловленных в Приладожье у выводков в разные сроки без разделения по полу (мм)
Table 4. Morphometric characteristics of the Common Sandpiper A. hypoleucos caught near broods in the Ladoga area in different time periods (in mm, sexes combined)
| Сроки | Крыло | Хвост | Цевка | ||||||
| N | lim | M ± m | N | lim | M ± m | N | lim | M ± m | |
| 30.05–5.06 | 52 | 108–119 | 113.4 ± 0.37 | 50 | 49–65 | 57.5 ± 0.41 | 46 | 23–29 | 24.9 ± 0.18 |
| 16–25.06 | 87 | 107–120 | 112.9 ± 0.34 | 84 | 50–64 | 57.3 ± 0.32 | 52 | 22–29 | 24.9 ± 0.19 |
| 26.06–1.07 | 39 | 104–118 | 111.9 ± 0.47 | 39 | 50–63 | 56.9 ± 0.50 | 21 | 21–28 | 24.9 ± 0.31 |
| Сроки | Клюв | Клюв2 | Клюв3 | ||||||
| N | lim | M ± m | N | lim | M ± m | N | lim | M ± m | |
| 30.05–5.06 | 51 | 22–28 | 24.9 ± 0.20 | 51 | 24–32 | 28.2 ± 0.24 | 50 | 16–20 | 18.1 ± 0.16 |
| 16–25.06 | 71 | 21–29 | 24.7 ± 0.18 | 71 | 25–31 | 28.1 ± 0.20 | 67 | 12–20 | 17.8 ± 0.16 |
| 26.06–1.07 | 37 | 21–29 | 25.2 ± 0.28 | 37 | 26–32 | 28.5 ± 0.22 | 27 | 10–22 | 17.9 ± 0.41 |
Тенденция к уменьшению размеров в течение лета прослеживается во всех возрастных группах (рисунок, А) и не связана с изменением их соотношения в выборках. Средняя длина крыла птиц всех возрастов, в соответствии с уравнением регрессии (y = 2919.99–0.008x), за 1.5 месяца сокращается примерно на 3 мм с достаточно высоким уровнем значимости (F = 5.96; p = 0.02). Рулевые перья обнашиваются медленнее (рисунок, В): их сокращение на 2 мм не значимо (F = 2.02, p = 0.16).
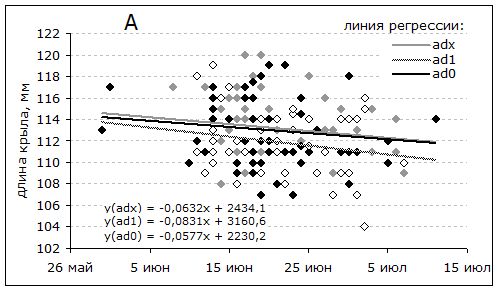
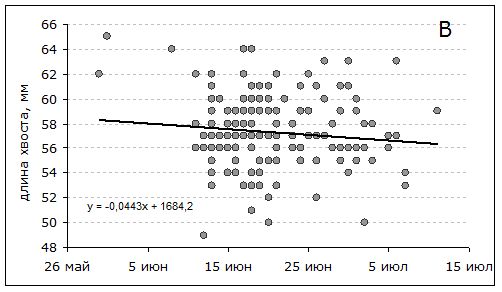
Зависимость длины крыла (А) и хвоста (В) перевозчиков A. hypoleucos в Приладожье от даты отлова в гнездовой период. Возраст: ad1 – 1 год, adx – ≥2 года, ad0 – не известен
Dependence of the wing and tail length of the of Common Sandpiper A. hypoleucos on the date of catching in the Ladoga area during the breeding season. age: ad1 – 1 year, adx – ≥2 years, ad0 – is not known
Размеры самок и самцов, отловленных в Приладожье, почти полностью перекрывались. Две птицы с самыми длинными крыльями (120 мм), пойманные в середине июня, оказались самкой (хвост 59 мм) и самцом (61 мм) из разных пар. Длина крыла самых мелких особей (2 самца, 1 самка, 1 с неопределенным полом) составила 107 мм (хвост 54–58 мм). Мельче них была только 1 годовалая птица с сильно обношенным оперением, отловленная в поздние сроки (2 июля) и определенная как самка: крыло – 104 мм, хвост – 50 мм. В то же время у всех этих птиц, как мелких, так и крупных, показатели длины клюва и цевки были близки к средним.
При определении пола иногда пытаются использовать разницу в размерах гнездящихся партнеров (Yalden, 2012). В Приладожье в 65 случаях удалось одновременно поймать и измерить обеих птиц, что позволило исключить влияние сроков обследования на степень изношенности их оперения (табл. 5). В паре из наиболее мелких птиц длина крыла одного партнера составила 107 мм, хвоста – 58 мм, другого соответственно 109 и 56 мм. В паре из самых крупных особей длина крыла обеих птиц составила 119 мм, хвоста – 59 мм у самца и 62 мм у самки.
Таблица 5. Частота обнаружения (число) птиц с разной длиной крыла в 65 парах перевозчиков A. hypoleucos, отловленных одновременно
Table 5. Occurence (number) of birds with different wing length in 65 pairs of the Common Sandpiper A. hypoleucos caught at the same time
| Мелкий партнер | Число птиц с данной длиной крыла, мм | ||||||||||||
| Крупный партнер | |||||||||||||
| Длина крыла | 109 | 110 | 111 | 112 | 113 | 114 | 115 | 116 | 117 | 118 | 119 | 120 | Всего |
| 107 | 1 | 1 | 2 | ||||||||||
| 108 | 1 | 1 | 1 | 3 | |||||||||
| 109 | 2 | 2 | 1 | 3 | 1 | 9 | |||||||
| 110 | 3 | 1 | 2 | 1 | 7 | ||||||||
| 111 | 3 | 1 | 2 | 4 | 6 | 4 | 1 | 21 | |||||
| 112 | 1 | 3 | 2 | 1 | 1 | 8 | |||||||
| 113 | 2 | 1 | 1 | 4 | |||||||||
| 114 | 2 | 2 | 2 | 1 | 7 | ||||||||
| 115 | 1 | 1 | |||||||||||
| 116 | 1 | 1 | 2 | ||||||||||
| 117–118 | – | ||||||||||||
| 119 | 1 | 1 | |||||||||||
| Всего | 1 | 3 | 7 | 4 | 13 | 8 | 14 | 8 | 4 | 2 | 1 | 65 | |
Средняя длина крыла крупного партнера составила 114.92 ± 0.28 мм, мелкого – 111.25 ± 0.27 мм (t = 9.4; p = 0.001). Однако результаты повторных отловов и наблюдений за маркированными особями в Приладожье показали, что при формировании пары кулики не принимают во внимание размеры друг друга: независимо от пола, у крупных особей партнеры обычно мельче, у мелких – крупнее. У особей средних размеров они могут быть как крупнее, так и мельче: из 17 птиц, сменивших в течение жизни нескольких партнеров, которых отлавливали одновременно с ними, 4 особи гнездились то с более крупной, то с более мелкой птицей (табл. 6, № 1, 3, 5, 6). Из-за этого по относительным размерам птиц в паре безошибочно определить пол партнеров невозможно.
Таблица 6. Результаты измерений длины крыла и хвоста птиц (I) и их партнеров (II), пойманных одновременно с ними
Table 6. The results of the wings and tails measurements of the birds (I) and their partners (II) caught at the same time
| № | Дата отлова | I. | II. | ||||||
| Кольцо | Возраст | Крыло, мм | Хвост, мм | Кольцо | Возраст | Крыло, мм | Хвост, мм | ||
| 1 | 28.06.1992 | xc713472 | ad0+1 | 109 | 56 | xd126942 | ad1? | 107 | 58 |
| 19.06.1995 | ad0+4 | 111 | 55 | xd505470 | ad0 | 112 | 57 | ||
| 2 | 26.06.1992 | xd126932 | ad0 | 113 | 57 | xd126931 | ad1 | 109 | 52 |
| 28.06.1993 | ad0+1 | 113 | 58 | xc713300 | ad1? | 109 | 55 | ||
| 3 | 19.06.1994 | б/с207619 | ad0 | 110 | 54 | б/с207624 | ad0 | 112 | 62 |
| 16.06.1995 | ad0+1 | 111 | 55 | xd505459 | ad0 | 109 | 54 | ||
| 4 | 21.06.1999 | xc714716 | ad x | 111 | 56 | xc714717 | ad1 | 112 | 58 |
| 14.06.2003 | ad x+4 | 111 | 57 | xd053230 | ad x? | 117 | 57 | ||
| 14.06.2004 | ad x+5 | 112 | 60 | xd053255 | ad0+1 | 116 | 57 | ||
| 5 | 01.07.2001 | xc714753 | ad1 | 114 | 60 | xc714754 | ad x | 113 | 59 |
| 15.06.2003 | ad3 | 111 | 59 | xd053255 | ad0 | 117 | 59 | ||
| 6 | 22.06.2001 | xd701200 | ad0 | 111 | xd701999 | ad1 | 108 | - | |
| 19.06.2004 | ad0+3 | 110 | 53 | Pb157399 | ad1 | 114 | 55 | ||
| 7 | 29.06.2002 | xd053058 | ad1 | 114 | 59 | xd053057 | ad0 | 111 | 56 |
| 13.06.2003 | ad2 | 116 | 58 | xd053203 | ad0 | 111 | 54 | ||
| 18.06.2004 | ad3 | 114 | 60 | -«- | ad0+1 | 111 | 51 | ||
| 8 | 11.06.2002 | б/с207800 | ad0 | 117 | 65 | xc714827 | ad1 | 111 | 58 |
| 12.06.2003 | ad0+1 | 117 | 57 | xd053086 | ad x | 111 | 56 | ||
| 02.07.2002 | ad1 | 116 | 56 | xd053069 | ad1 | 115 | 58 | ||
| 9 | 15.06.2003 | xd053068 | ad2 | 114 | 57 | xd053250 | ad0 | 114 | 58 |
| 13.06.2004 | ad3 | 115 | 56 | -«- | ad0+1 | 114 | 58 | ||
| 10 | 13.06.2003 | ad x | 113 | 61 | xd053209 | ad1 | 110 | 55 | |
| 18.06.2004 | xd053208 | ad x+1 | 116 | 59 | -«- | ad2 | 111 | 57 | |
| 17.05.2005 | ad x+2 | 115 | 62 | xd053315 | ad1 | 111 | 57 | ||
| 11 | 13.06.2003 | xd053221 | ad1 | 116 | 60 | xd053220 | ad1? | 113 | 62 |
| 13.06.2004 | ad2 | 116 | 58 | Pb157370 | ad x | 112 | 55 | ||
| 12 | 14.06.2003 | xd053239 | ad x | 115 | 56 | xd053245 | ad x | 108 | 56 |
| 16.06.2004 | ad x+1 | 114 | 55 | Pb157377 | ad0 | 112 | 54 | ||
| 13 | 14.06.2003 | xd053245 | ad x | 108 | 56 | xd053239 | ad x | 115 | 56 |
| 17.06.2004 | ad x+1 | 109 | 53 | xc714832 | ad3 | 116 | 58 | ||
| 14 | 15.06.2003 | xd053255 | ad0 | 117 | 59 | xc714753 | ad3 | 111 | 59 |
| 14.06.2004 | ad0+1 | 116 | 57 | xc714716 | ad x+5 | 112 | 60 | ||
| 15 | 18.06.2003 | xd053258 | ad0 | 110 | 54 | xd053259 | ad0 | 116 | 56 |
| 19.06.2004 | ad0+1 | 113 | 53 | xc714834 | ad2 | 120 | 59 | ||
| 16 | 18.06.2003 | xd053259 | ad0 | 116 | 56 | xd053258 | ad0 | 111 | 52 |
| 20.06.2004 | ad0+1 | 115 | 59 | Pb157400 | ad x | 110 | 54 | ||
| 17 | 30.06.2005 | xd053331 | ad1 | 109 | 56 | xd053332 | ad1 | 114 | 61 |
| 16.06.2007 | ad3 | 110 | 56 | xd053365 | ad1? | 112 | 57 | ||
Обсуждение
Как и у других птиц, индивидуальные размеры перевозчика варьируют в довольно широких пределах. У птиц европейских популяций величина наиболее популярного показателя – длины крыла – варьирует между 102 и 121 мм, средние значения изменяются между 113.1 мм у особей, отловленных весной в свежем оперении, до 110.1 мм – осенью (Brown, 1973; Glutz von Blotzheim et al., 1977; Meissner, 1997; Yalden, 2012; Meissner, Krupa, 2016 и др.). Гнездовую популяцию из центральной Норвегии характеризует средний показатель 112.4 мм (Løfaldli, 1981), британскую – 113.3 мм (Robson, 1977), однако расхождение между ними может быть обусловлено не только популяционными особенностями, но и большей обношенностью оперения птиц на севере из-за поздних сроков прилета и отловов, а также способом измерения.
По морфометрическим показателям гнездовая группировка перевозчиков Приладожья не выделяется среди европейских популяций, а индивидуальные размеры варьируют в тех же широких пределах. Предельные значения для длины крыла – 104 и 120 мм, средний показатель – 112.7 мм.
Судя по величине партнеров в парах из самых мелких и самых крупных птиц (см. табл. 5), размеры самцов и самок перекрываются почти полностью, как и у перевозчиков других европейских популяций. Предельные значения длины крыла птиц разного пола, фигурирующие в литературных источниках, – 102–119 и 107–120 мм соответственно, клюва – 20.9–27.6 и 22.9–27.5 мм, цевки – 20.8–27.1 и 21.6–27.1 мм. При этом разные авторы, констатировавшие, что определить пол, используя промеры частей тела птиц, можно только у экстремально мелких или крупных особей, указывают разные границы отсечения (Meissner et al., 2015). При работе с гнездовой популяцией в Англии при определении пола наряду с другими способами использовали относительные размеры птиц в паре (Yalden, 2012). Указанные в данной статье характеристики хорошо согласуются с показателями для крупных и мелких партнеров в Приладожье: длина крыла самок составила 114.4 мм (107–120, N = 107), самцов – 110.8 мм (103–119, N = 92). Однако, поскольку размеры птиц не играют роли при формировании пары (см. табл. 6), различия между полами, вероятно, значительно меньше.
На примере берингийских популяций чернозобика Calidris alpina было показано, что средние размеры самок и самцов куликов могут отличаться не только у разных подвидов, но и у птиц одного подвида в разных точках отлова (Gates et al., 2013). По литературным данным, в Западной Европе самки перевозчика обычно в среднем крупнее самцов (Løfaldli, 1981; Yalden, 2012; Meissner, Krupa, 2016 и др.), в Китае – мельче (Sandpiper…, 2016), объединение птиц из Европы и Азии дало одинаковые цифры (Cramp, Simmons, 1983).
В западной части ареала вида наибольшие различия в размерах птиц разного пола демонстрируют перевозчики, гнездящиеся в Норвегии: длина крыла – 108.7 и 115.3 мм (Løfaldli, 1981), и относительно небольшие, по музейным экземплярам, в центре и на севере европейской части России – 110.7 и 109.2 мм (Glutz von Blotzheim et al., 1977). Однако делать выводы о половых особенностях морфометрии птиц разных географических популяций по имеющимся данным не вполне корректно. Все авторы приводят средние показатели без учета соотношения в выборках птиц из разных возрастных групп и сроков их отлова. Вместе с тем у годовалых птиц они меньше, чем у особей старших возрастов, и у всех значимо меняются к концу сезона из-за обноса оперения. Картину искажают и ошибки при прижизненном определении пола, неизбежные из-за отсутствия у вида надежных половых маркеров.
Появление молекулярных методов исследований открыло новые возможности (Нестеренко и др., 1996; Griffiths et al., 1998 и др.). В 2005–2015 гг. в 3 пунктах Польши ПЦР-методом определили пол и промерили 247 самцов и 111 самок перевозчиков, отловленных на миграции с начала июня по середину сентября (Meissner, Krupa, 2016). По всем показателям самцы, мигрировавшие через Балтику, лишь немного уступали самкам, и только по средней длине крыла разрыв оказался значительным – 111.3 и 114.8 мм. Интересно, что эти цифры практически совпали с показателями для крупных и мелких партнеров в Приладожье – 111.3 и 114.9 мм (см. табл. 5). Проведя дискриминантный анализ по трем параметрам, включая размеры крыла, авторы вычислили индексы, отличавшие пол большинства отловленных ими особей. Однако сравниваемые ими выборки не вполне равноценны, поскольку самки были многочисленными лишь в начале миграции и, соответственно, имели менее обношенное оперение. В регионе, где проводились исследования, массовый пролет идет позднее – в конце июля – середине августа (Meissner, 1996). С июня по август, судя по линии регрессии, длина крыла перевозчиков может сократиться за счет обноса более чем на 5 мм (Iwajomo, Hedenström, 2011). Исходя из этого «базовые» различия между полами по данному показателю все же не столь значительны. Поэтому применение предложенных критериев для гнездящихся птиц с близкими сроками отлова может оказаться не столь эффективным и требует предварительной проверки.
Из-за более ранних сроков отлова самок складывается впечатление, что они мигрируют раньше. Однако обращает на себя внимание существенное нарушение соотношения полов среди птиц, пойманных в Польше: во всех трех пунктах работы число самок резко уступало числу самцов (1 : 2.2). Кроме того, в отличие от самцов их распределение в выборках по длине крыла было далеким от нормального, а доля особей с минимальными размерами (111 мм) сравнительно велика – около 7 %. Все это указывает на то, что самки этого трансконтинентального мигранта, способного к дальним броскам, могут использовать иные, чем у самцов, пути или другую миграционную стратегию, транзитом пересекая многие районы Европы. Подобное разделение на пролете особей разного пола уже обнаружено у некоторых видов Ржанкообразных (Nebel, 2007).
Не исключено также, что самки и самцы перевозчика из разных регионов летят разными путями, используя иные места для остановок. Предположение о присутствии на стоянках в Польше птиц разных географических популяций соответствует «слегка бимодальному» (slightly bimodal) распределению особей по длине крыла в обеих выборках (Meissner, Krupa, 2016). В европейской части ареала осенняя миграция перевозчиков в том числе из соседней с Карелией Ленинградской области идет в юго-юго-западном направлении (Резвый и др., 1995). Судя по обнаружению в мае 1986 г. в Муезерском районе Карелии (63.30 N 31.40 E) годовалой птицы, окольцованной под Хельсинки 11.07.85 во время миграции, перевозчики, по крайней мере, из северной Карелии придерживаются этого же направления. Вполне вероятно, что часть «карельских» птиц после пересечения Балтики останавливается на отдых в Польше, однако доказать это на основе морфометрических показателей пока не представляется возможным.
Заключение
Перевозчики, гнездящиеся на северо-западе России и попадающие на зимовки в Африку через Западную Европу, по-видимому, достаточно тесно контактируют с птицами европейских популяций. Почти идентичные морфометрические характеристики гнездового населения восточного Приладожья хорошо согласуются с данными для других точек Европы. Их сближают близкие предельные и средние значения всех параметров, одинаково широкая амплитуда их колебаний, сходные темпы сокращения длины крыла и хвоста к концу сезона по мере обноса оперения. Показано также, что в Приладожье годовалые впервые гнездящиеся птицы мельче особей старших возрастов, а после 2-го года длина крыла и хвоста одной особи может варьировать по годам без определенной тенденции. Прижизненное определение пола после окончания предбрачного периода затруднено: размеры самок и самцов почти полностью перекрываются, а формирование пар происходит без учета размеров партнеров. Эффективность применения для гнездовых популяций вида морфометрических индексов, предлагаемых для определения пола (Meissner, Krupa, 2016), нуждается в проверке, поскольку они вычислены на основе измерений пролетных птиц без учета наличия в выборках птиц разных возрастов и разной степени обноса оперения особей, мигрирующих в разные сроки.
Библиография
Виноградова Н. В., Дольник В. Р., Ефремов В. Д., Паевский В. А. Определение пола и возраста воробьиных птиц фауны СССР . М.: Наука, 1976. 189 с.
Гладков Н. А. Перевозчик Actitis hypoleucos L. // Птицы Советского Союза / Под ред. Г. П. Дементьева и Н. А. Гладкова. М.: Сов. наука, 1951. Т. 3. С. 241–255.
Ивантер Э. В., Коросов А. В. Введение в количественную биологию . Петрозаводск: Изд-во ПетрГУ, 2011. 304 с.
Козлова Е. В. Ржанкообразные. Подотряд Кулики . Серия: Фауна СССР. Птицы. Т. 2. Вып. 1. Ч. 2. М.; Л.: АН СССР, 1961. 502 с.
Нестеренко О. Н., Антоненкова В. Г., Алискеров С. В. Определение пола птиц по хромосомным препаратам // Научные исследования в зоологических парках. М., 1996. № 6. С. 80–86.
Резвый С. П., Носков Г. А., Гагинская А. Р. и др. Атлас миграций птиц Ленинградской области по данным кольцевания . Л.: СПб об-во естествоиспытателей, 1995. 236 с.
Хохлова Т. Ю., Лунина Т. Л. Методы и основные результаты контроля популяции перевозчика Actitis hypoleucos в восточном Приладожье (Карелия) // Материалы юбилейной научно-практ. конф. «Роль заповедников России в сохранении и изучении природы»: Труды Окского гос. природного биосферного заповедника. Вып. 24. Рязань: НП «Голос губернии», 2015а. С. 172–175.
Хохлова Т. Ю., Лунина Т. Л. Особенности гнездовой биологии и территориального поведения перевозчика Actitis hypoleucos L. в Восточном Приладожье // Вопросы экологии, миграции и охраны куликов Северной Евразии: Материалы 10-й юбилейной конф. рабочей группы по куликам Северной Евразии / Ред. И. И. Черничко, В. Н. Мельников. Иваново: Изд-во Ивановского гос. ун-та, 2016. С. 396–399.
Штифель А., Приклонский С. Г., Постельных В. А. Перевозчик – Actitis hypoleucos L. // Миграции птиц Восточной Европы и Северной Азии: Журавлеобразные – ржанкообразные / Ред. В. Д. Ильичев. М.: Наука, 1985. С. 126–143.
Blasco-Zumeta, Heinze. Common Sandpiper Ageing and sexing by // Ibercaja Aula en Red. URL2: http://aulaenred.ibercaja.es/wp-content/uploads/Ahypoleucos.pdf (дата обращения 13.12.2016).
Brown S. C. Common Sandpiper biometrics // Wader Study Group Bull. 1973. Vol. 11. P. 18–23.
Cramp S., Simmons K. E. L. (eds.). The Birds of the Western Palearctic. Vol. III. Waders to Gulls. Oxford; New York: Oxford University Press, 1983. 913 p.
Dougall T. W., Holland P. K., Mee A., Yalden D. W. Comparative population dynamics of Common Sandpipers Actitis hypoleucos: living at the edge: Capsule Common Sandpiper populations at the edge of their range do not recruit so well // Bird Study. 2005. Vol. 52. № 1. Р. 80–87. DOI: 1080/00063650509461376.
Gates H. R., Yezerinac S., Powell A. N., Tomkovich P. S., Valchuk O. P., Lanctot R. B. Differentiation of subspecies and sexes of Beringian dunlin using morphometric measures // Journal of Field Ornithology. 2013. Vol. 84. P. 389–402.
Glutz von Blotzheim U. N., Bauer K. M., Bezzel E. Handbuch der Vögel Mitteleuropas. Vol. 7. Wiesbaden: Aula-Verlag, 1977. 895 p.
Griffiths R., Double M. C., Orr K., Dawson R. J .G. A DNA test to sex most birds // Molecular Ecology. 1998. № 7. P. 1071–1075.
Holland P. K., Robson J. E., Yalden D. W. The breeding biology of the common sandpiper Actitis hypoleucos in the Peak Distric // Bird Study. 1982. Vol. 29. № 2. P. 99–110.
Holland P. K., Yalden D. W. Population dynamics of Common Sandpipers Actitis hypoleucos breeding along an upland river system // Bird Study. 1991. Vol. 38. P. 151–159.
Hung C.-M., Drovetski S., Zink R. Multilocus test of the absence of mtDNA phylogeographic structure in a widespread wader, the Common Sandpiper (Actitis hypoleucos) // Journal of Ornithology. 2013. Vol. 154. P. 1105–1113.
Iwajomo S. B., Hedenström A. Migration patterns and morphometrics of common sandpipers Actitis hypoleucos at Ottenby, southeastern Sweden // Ringing and Migration. 2011. Vol. 26. № 1. P. 38–47.
Løfaldli L. On the breeding season biometrics of the common sandpiper // Ringing and Migration. 1981. Vol. 3. № 3. P. 133–136.
Meissner W. Timing and phenology of autumn migration of common sandpiper (Actitis hypoleucos) at the Gulf of Gdansk // The Ring. 1996. Vol. 18. P. 59–72.
Meissner W. Autumn migration and biometrics of the Common Sandpiper Actitis hypoleucos caught in the Gulf of Gdańsk // Ornis Fennica. 1997. Vol. 74. P. 131–139.
Meissner W., Holland P. K., Cofta T. Ageing and sexing series 11: Ageing and sexing the Common Sandpiper Actitis hypoleucos // Wader Study. 2015. Vol. 122. № 1. P. 54–59.
Meissner W., Krupa R. Identifying the Sex of the Common Sandpiper (Actitis hypoleucos) by Linear Measurements // Ann. Zool. Fennici. 2016. Vol. 53. P. 175–182.
Nebel S. Differential migration of shorebirds in the East Asian-Australasian Flyway // Emu. 2007. Vol. 107. P. 14–18.
Robson J. E. Brief note on British breeding Common Sandpipers // Wader Study Group Bull. 1977. Vol. 21. P. 7–8.
Sandpiper // Shared Encyclopedia URL1: http://www.et97.com/view/254418.htm (дата обращения 13.12.2016).
Valkama J., Saurola P., Lehikoinen A., Lehikoinen E. Piha M., Sola P., Velmala W. Suomen rengastusatlas. Osa II. Helsinki: Luonnontieteellinen keskusmuseo & ympäristöministeriö, 2014. 784 s.
Wernham C. V., Toms M. P., Marchant J. H., Clark J. A., Siriwardena G. M., Baillie S. R. (eds.). The Migration Atlas: Movements of the Birds of Britain and Ireland. London: T. & A. D. Poyser, 2002. 900 p.
Yalden D. W. Wing area, wing growth and wing loading of Common Sandpipers Actitis hypoleucos // Wader Study Group Bull. 2012. Vol. 119. № 2. P. 84–88.
Zink R. M., Pavlova A., Drovetski S., Rohwer S. Mitochondrial phylogeographies of five widespread Eurasian birds // Journal of Ornithology. 2008. Vol. 149. P. 399–413.
Благодарности
Выражаю глубокую признательность сотрудникам лаборатории зоологии КарНЦ РАН В. Б. Зимину†, А. В. Артемьеву, Н. В. Лапшину, студентам ПетрГУ Т. Л. Янкелович, С. А. Барановой, Л. Г. Корвяковой, Н. А. Улицкой и многим другим, кто оказывал помощь в поиске гнезд, отловах птиц и их ежедневном контроле на стационаре «Маячино». Благодарю П. С. Томковича, взявшего на себя труд ознакомиться с рукописью, за внимательное отношение, ценные замечания и комментарии, которые позволили дополнить и значительно улучшить текст статьи.

 © 2011 - 2026
© 2011 - 2026