



Введение
Насекомые – обязательные участники круговорота веществ и энергии в природных биогеоценозах. Циклы их развития связаны со многими компонентами экосистемы. Наблюдения за насекомыми позволили сформулировать в 1905 г. одну из классических теорий в науке – о «приливах и отливах жизни». По мнению автора данной теории, в работе «отчасти удалось указать на те постоянные, вечные и глубокие изменения, какие испытывает фауна какой-либо местности под влиянием непрерывно поднимающихся и опускающихся волн видовой жизни» (Четвериков, 2009). «Динамические характеристики популяции – это величины, оценивающие интенсивность происходящих в ней процессов. Межгодовые колебания численности часто оказываются тесно коррелированными с климатическими условиями, воздействующими на изучаемые организмы» (Гиляров, 1990). Общеизвестно, что на живую природу воздействуют такие космические ритмы, как периодические изменения солнечной активности (Чернова, Былова, 2004). Вспышки на Солнце – закономерное явление (Стародубцев, 2011), оказывающее влияние на природные циклы (Ягодинский, 1975, 1981; Кривенко, Виноградов, 2008). Идея о циклических изменениях всех земных процессов под влиянием энергии Солнца нашла отражение в научных исследованиях различных групп живых организмов. Связь между популяционными волнами и периодичностью солнечных вспышек изучена для практически значимых насекомых – вредителей сельского и лесного хозяйства. Прогнозы динамических циклов насекомых необходимы для регуляции их численности. Отмечено, что солнечная активность создает циклический фон массовых размножений для непарного шелкопряда – Lymantria dispar (Linnaeus, 1758) (Бенкевич, 1990; Станкевич и др., 2019; Лямцев, 2019). Проблемы динамики численности насекомых подробно рассмотрены на примере клопа вредной черепашки – Eurygaster integriceps Puton, 1881 (Викторов, 1967; Капусткина, Хилевский, 2020). Исторические обзоры о развитии идеи зависимости колебаний динамики численности (волн жизни) в биологических системах от разных факторов рассмотрены А. Н. Фроловым (2017) и Е. Н. Белецким с соавт. (2017). Прогнозирование численности популяций кровососущих комаров (Diptera: Culicidae) представляет научный и практический интерес. Ранее было отмечено, что для получения корректных данных по экологии кровососущих комаров необходимо проведение стационарных наблюдений за динамикой их численности (Николаева, 1994). Информация о биоритмах необходима для биологического обоснования профилактических и истребительных мероприятий против комаров и прогнозирования их численности (Виноградова, Карпова, 2010).
Обзор особенностей экологии на разных фазах развития кровососущего комара Aedes communis (De Geer, 1776) представлен в статье Медведева с соавт. (2010). Этой работой были заложены методологические основы изучения данного вида в стационарных условиях на Северо-Западе России (Панюкова, Мади, 2014). Известно, что комар A. communis имеет практическую значимость как потенциальный переносчик лимфоцитарного хориоменингита и туляремии (Гуцевич и др., 1970; Остроушко и др., 2007; Панюкова, Грицай, 2019). В Новгородской области ранее были отмечены волнообразные изменения относительной численности A. communis в многолетней динамике с 1991 по 1998 г.: в 1991 г. процентное соотношение A. communis в составе лесного комплекса комаров составляло 39.2 %, в 1992 г. – 40.4, 1993 г. – 10, 1994 г. – 4.4, 1995 г. – 13, 1996 г. – 26.4, 1997 г. – 32, 1998 г. – 48 % (Кункова, Федорова, 1999).
Цель настоящей работы – провести анализ особенностей многолетних ритмов A. communis в сравнении с циклами солнечной активности на примере стационарных исследований в пределах лесной зоны Республики Коми. В задачи исследований входило наблюдение за фенологией преимагинальных стадий и сезонной активностью нападений на человека (пищевой активностью) имаго A. communis.
Материалы
Многолетние исследования относительной численности комара A. communis выполнены в окрестностях г. Сыктывкара (с. Выльгорт Сыктывдинского р-на Республики Коми, 61°38ʹ с. ш., 50°43ʹ в. д.). Климат данной территории умеренно-континентальный, среднегодовое количество осадков составляет 700 мм, что превышает испарение с поверхности суши (500 мм в год). При избыточном увлажнении здесь преобладают дерново-подзолистые и торфянистые почвы. Территория относится к центральной части подзоны средней тайги. Характерной растительной ассоциацией для этой подзоны считается ельник чернично-зеленомошный (Мартыненко, 1999).
Стационарные наблюдения проведены с 2005 по 2021 г. в ельнике чернично-зеленомошном с примесью березы, где был выбран модельный водоем на окраине леса (ежегодно образующаяся снеговая лужа, 50 × 25 × 0.5 м). В работе проанализированы данные о 4672 личинках и 2068 имаго кровососущего комара A. communis. Наблюдения вели ежедекадно с апреля по июль. С момента образования водоема наблюдали за изменениями температуры воды и окружающего воздуха. Фенологические наблюдения за личинками комаров проводили с момента появления личинок первого возраста еженедельно до прекращения их развития в водоемах (окукливания и вылета имаго). Отмечали даты развития личинок (всех возрастов), куколок, а также сроки начала и окончания нападения имаго комаров на человека.
Методы
Измерения температуры воды в водоеме проводили спиртовым термометром в момент сбора преимагинальных стадий. Сборы личинок и куколок вели кюветой (18 × 22 см), брали по 5 проб в разных частях водоема, как и ранее (Panyukova et al., 2018). За единицу измерения относительной численности личинок принята плотность экземпляров на 1 м2 водной поверхности (Николаева, 1980). Под бинокулярным микроскопом (ЛОМО МССО или МБС-10) личинок старшего возраста определяли до вида прижизненно, подтверждая затем достоверность определений по признакам выведенных имаго, как это принято при составлении определительных таблиц и в общих методиках (Сазонова, 1958; Халин и др., 2021).
Имаго собирали на предплечье левой руки учетчика в 5-миллилитровые пробирки, на 2/3 заполненные 70%-ным этанолом. При каждом учете отмечали время, в течение которого на учетчика нападали 10 самок. Затем рассчитывали число особей каждого вида в перерасчете на стандартный 20-минутный учет, которое принималось за единицу измерения относительной пищевой активности имаго. Время сбора 10 имаго составляло от нескольких секунд до 20 минут в зависимости от погодных условий и числа нападающих особей. Комаров определяли в лаборатории под бинокулярным микроскопом, используя классические определители (Гуцевич и др., 1970; Гуцевич, Дубицкий, 1981; Becker et al., 2010).
Записи наблюдений вели в дневнике учетов, затем вносили в электронную таблицу базы данных в программе Microsoft Excel. Ряды полученных данных были использованы для построения графиков годовой динамики относительной численности личинок A. communis и относительной пищевой активности имаго (самок). Статистический анализ собранного материала выполнен в программе PAST версий 2.15 и 3.17 (Hammer et al., 2001), рассчитывали коэффициент Спирмена (r-Spearman's) при уровне значимости α = 0.05, чем ближе значение данного коэффициента к 1, тем выше достоверность связи. Результат считали статистически значимым, если уровень вероятности (р) не превышал 0.05 (p ≤ 0.05 > 0.01 – низкая, p ≤ 0.01 > 0.001 – средней силы, p ≤ 0.001 – высокая статистическая значимость). Расчеты коррелятивных связей A. communis с солнечной активностью проведены с помощью кросс-корреляционного анализа, как и ранее Фроловым при изучении связей с гелиоциклической активностью обработок посевных площадей против лугового мотылька – Loxostege sticticalis Linnaeus, 1761 (Фролов, 2015). Для статистического анализа связей выбраны данные (число Вольфа) за май и апрель с 2005 по 2021 г. в связи с устойчивыми датами перехода среднесуточных температур воздуха через 0 °С (21.04 – для апреля) и +5 °С (1.05 – для мая). Данные месяцы связаны с установлением положительных температур воздуха на изучаемой территории (Братцев, 2011а, б), именно в этот период начинается развитие A. communis. В анализе использовали климатические показатели о среднемесячных температурах воздуха и количестве осадков по наблюдениям на метеостанции в г. Сыктывкаре (табл. 1).
Таблица 1. Климатические характеристики в весенне-летний период в годы исследований (г. Сыктывкар)
| Год | Апрель | Май | Июнь | |||
| температура (°С) | осадки (мм) | температура (°С) | осадки (мм) | температура (°С) | осадки (мм) | |
| 2005 | -9.4 | 82 | 1.7 | 31.4 | 12.2 | 53 |
| 2006 | -8 | 18.7 | 2.4 | 63.1 | 9.7 | 52.4 |
| 2007 | -0.8 | 60.7 | 2.3 | 89.5 | 9.3 | 33.4 |
| 2008 | -3.1 | 52.8 | 1.5 | 57.6 | 6.4 | 40 |
| 2009 | -3.8 | 23.7 | -0.8 | 43.6 | 8.8 | 126.8 |
| 2010 | -6.3 | 22.4 | 4.6 | 35 | 13.1 | 96.3 |
| 2011 | -4.7 | 61.7 | 2.9 | 47 | 10.9 | 32.8 |
| 2012 | -7.9 | 66.3 | 4.3 | 25.8 | 10.8 | 143 |
| 2013 | -14.4 | 15.3 | 2.2 | 43.7 | 8.9 | 34.9 |
| 2014 | -2 | 24.4 | 1 | 46.3 | 10.7 | 106 |
| 2015 | -1.8 | 35.7 | 2.9 | 40.3 | 13.6 | 57.5 |
| 2016 | -4.7 | 55.9 | 4.8 | 35.6 | 11.4 | 53 |
| 2017 | -1.2 | 76.3 | 1 | 61 | 5 | 95.7 |
| 2018 | -10.8 | 26 | 1.6 | 80 | 8 | 75 |
| 2019 | -3.5 | 36.8 | 2.1 | 84.3 | 10.9 | 94.1 |
| 2020 | -1.3 | 47.7 | 1 | 63.4 | 10.2 | 41.2 |
| 2021 | -6 | 16.8 | 4.9 | 62.5 | 12.1 | 62.6 |
Для характеристики солнечной активности взяты литературные данные из опубликованного каталога солнечных пятен с 1874 по 2020 г. (Veronig et al., 2021). За 2021 г. данные о числе солнечных пятен (в апреле и мае) взяты с сайта Королевской бельгийской обсерватории (WDC-SILSO, Royal Observatory of Belgium).
Начиная с 2009 г. проведение стационарных наблюдений совпало с уникальным 24-м циклом солнечной активности, который длился ровно 11 лет. «Под циклом понимается законченный процесс, элементы которого (фазы, стадии, этапы и т. д.), следуя друг за другом или чередуясь, составляют единое целое» (Белецкий и др., 2017). Структура 11-летнего цикла солнечной активности состоит из более кратковременных элементов – импульсов солнечной активности (Витинский, 1983). Исследования сезонной активности Aedes communis включали завершающий период 23-го цикла солнечной активности (2005–2008 гг.) и период уникального, рекордно слабого 24-го цикла солнечной активности, который начался в январе 2009 г. и закончился в декабре 2019 г. (по данным Института земного магнетизма, ионосферы и распространения радиоволн РАН (ИЗМИ РАН)).
Результаты
Динамика относительной пищевой активности имаго на стационаре, в ельнике чернично-зеленомошном с примесью березы в окрестностях г. Сыктывкара (с. Выльгорт), представляет собой волнообразные циклы или волны жизни (рис. 1).
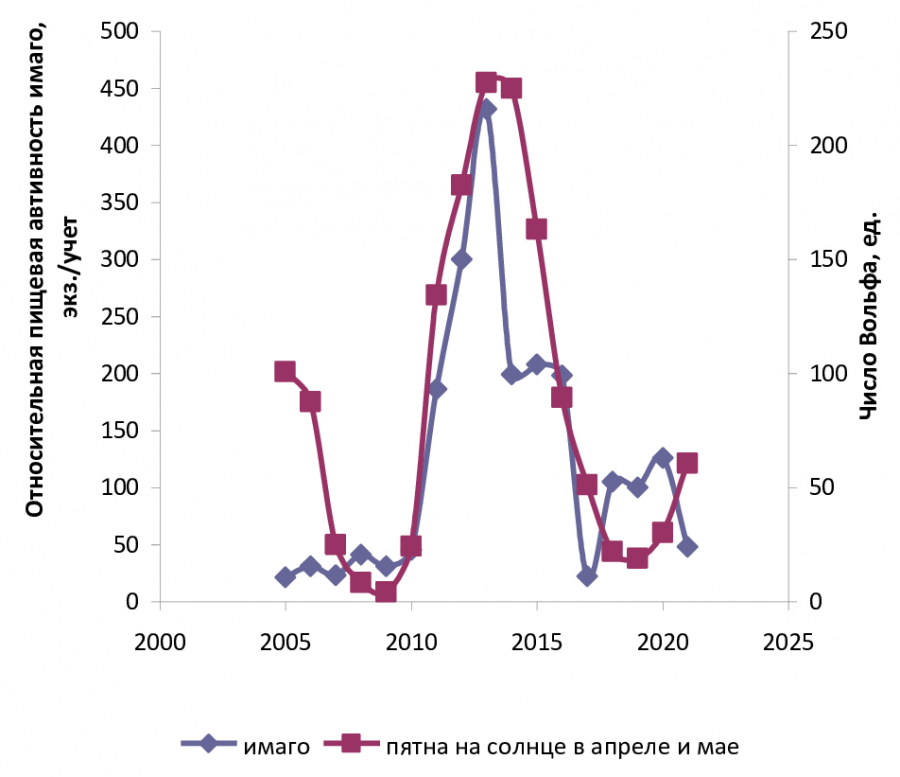
Рис. 1. График динамики популяции A. communis: относительная пищевая активность самок в сравнении с ритмами солнечной активности (Республика Коми, с. Выльгорт, 61°38ʹ с. ш., 50°43ʹ в. д.)
Fig. 1. Graph of A. communis population dynamics: relative food activity of females in relation to the rhythms of solar activity (Komi Republic, Vylgort village, 61°38ʹ N, 50°43ʹ E)
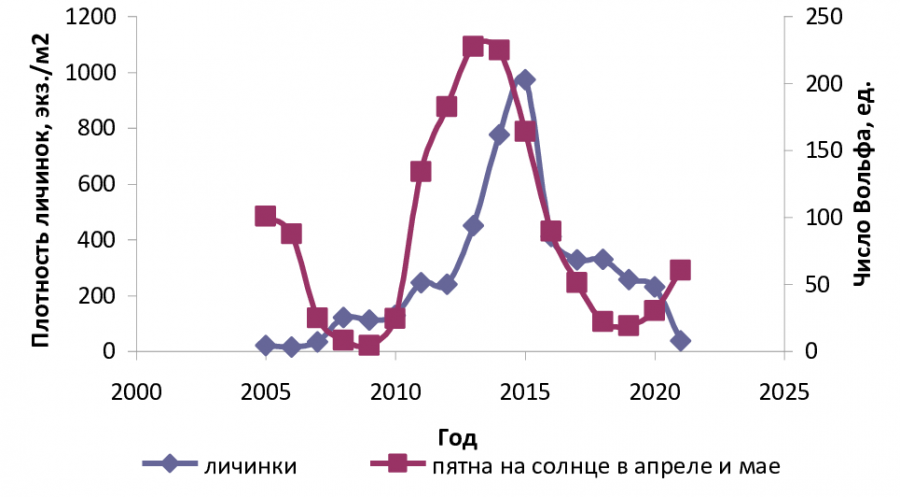
Рис. 2. График динамики популяции A. communis: плотность личинок в модельном водоеме в сравнении с ритмами солнечной активности (Республика Коми, с. Выльгорт, 61°38ʹ с. ш., 50°43ʹ в. д.)
Fig. 2. Graph of A. communis population dynamics: density of larvae in the model water body in relation to the rhythms of solar activity (Komi Republic, Vylgort village, 61°38ʹ N, 50°43ʹ E)
Росту относительной численности личинок в модельном водоеме предшествовал пик активности нападений A. communis, который приходился на 2011‒2013 гг., наибольшее число личинок данного вида отмечено в последующие 2014 и 2015 гг. (рис. 2). Низкая активность нападений на человека имаго комаров была отмечена в 2005–2009, 2017 и 2021 гг. при низкой солнечной активности.
На начало 25-го цикла солнечной активности (2020‒2021 гг.) отмечались спад относительной численности личинок и последовавший за этим спад нападений имаго A. communis летом 2021 г.
По полученным данным, цикл популяционной динамики комара A. communis отстает от цикла солнечной активности более чем на год, т. е. определяется предыдущим годом или двумя годами цикла солнечной активности. Большее влияние Солнце оказывает на имаго A. communis, чем на личинок, относительная численность которых зависит от успешности имаго самок при откладке яиц в предыдущие годы.
Волнообразные многолетние циклы личиночной популяции и активность нападений имаго A. communis различались по годам, пики и спады их не накладывались на цикличность солнечной активности, а закономерно изменялись: сначала наблюдался пик пищевой активности имаго, затем пик солнечной активности и следующий за ним пик плотности личиночной популяции. Периоды подъема волн жизни A. communis ярко выражены, максимальные значения плотности личинок наблюдаются с отставанием от цикла солнечной активности на 1–2 года (табл. 2).
Таблица 2. Кросс-корреляционный анализ зависимости цикличности популяции A. communis на преимагинальных стадиях (личинки) от солнечной активности в апреле в 2005‒2021 гг.
| Лаг (отставание), год | Коэффициент корреляции | р |
| ‒5 | ‒0.659 | 0.021 |
| ‒4 | ‒0.459 | 0.113 |
| ‒3 | ‒0.221 | 0.445 |
| ‒2 | 0.045 | 0.871 |
| ‒1 | 0.271 | 0.309 |
| 0 | 0.591 | 0.012 |
| 1 | 0.786 | 0.0003 |
| 2 | 0.746 | 0.001 |
| 3 | 0.519 | 0.056 |
| 4 | 0.091 | 0.766 |
| 5 | ‒0.405 | 0.190 |
Период развития личинок в модельном водоеме составлял от двух недель до месяца в зависимости от погодных условий весеннего сезона. Начало развития личинок соответствовало температурному порогу в 4 °С, при этом понижение температуры воды до 2 °С не вызывало их гибели. Колебания температуры воды в водоеме оказывали влияние на скорость развития личинок A. communis. По нашим наблюдениям, повышение температуры воды в водоеме до 20 °С и понижение до 2 °С приводили к изменению двигательной активности личинок A. communis – они медленнее, чем обычно, опускались на дно при беспокойстве.
Примененный анализ не позволил выявить связи между температурами воздуха, количеством осадков и относительной численностью особей комаров на разных стадиях развития. Среднемесячные температуры воздуха и количество осадков в период активного развития и питания A. communis (с апреля по июль) не коррелировали с полученными данными об относительной численности личинок и пищевой активности (активности нападений) имаго самок. Отсутствуют также связи между среднемесячными температурами воздуха и солнечной активностью в указанные месяцы. Отмечена корреляция между относительной пищевой активностью самок комаров данного вида и солнечной активностью для апреля (r = 0.56) и мая (r = 0.59). Корреляция между значениями относительной численности личинок и пищевой активности имаго самок достаточно высокая (r = 0.73). Существует тенденция зависимости относительной плотности личинок A. communis от солнечной активности в апреле (r = 0.44) и мае (r = 0.36).
В отдельные годы исследований наблюдали влияние весенних заморозков на личинок изучаемого вида, которые начали свое развитие в предшествовавшую похолоданию оттепель. Так, во второй декаде мая 2011 г. весенние заморозки привели к образованию на поверхности водоема корки льда (рис. 3). Погибли только вмерзшие в лед личинки A. communis в краевых участках водоема. Большинство личинок сохранило жизнеспособность, они медленно двигались подо льдом. В апреле 2021 г. (30.04) вновь наблюдали развитие личинок первых возрастов A. communis подо льдом при температуре воды 2 °С.

Рис. 3. Модельный водоем в период весеннего похолодания (12.05.2011), развитие личинок Aedes communis (De Geer, 1776) подо льдом
Fig. 3. Model reservoir during spring cold snap (12.05.2011), development of Aedes communis (De Geer, 1776) larvae under ice
Преимагинальные стадии изучаемого вида обнаруживались в водоемах в течение нескольких недель (табл. 3). Появление первого возраста личинок в водоемах отмечено при температуре воды 2‒4 °C. Четвертый возраст личинок встречался при температуре воды 10 °C. При температуре воды 15‒20 °C рост личинок и куколок A. communis заканчивался, отмечался массовый вылет имаго. Первыми окукливались и вылетали самцы, через 2‒4 дня вылетали самки. Взрослые особи самцов встречались на траве, кустарничках и стволах деревьев, недалеко от водоема. Самки начинали нападать на человека через несколько дней после обнаружения в природе самцов.
Таблица 3. Фенодаты событий жизненного цикла популяции комара A. communis за годы исследований (2005–2021) (Республика Коми, г. Сыктывкар, с. Выльгорт)
| Стадия | Начало фенопериода (первые находки стадии) | Окончание фенопериода (последние находки стадии) | Амплитуда (дни) | Продолжительность фазы (дни) |
| Первый возраст личинок | 26.04 | 10.05 | 14 | 3‒7 |
| Второй возраст личинок | 11.05 | 21.05 | 10 | 3‒7 |
| Третий возраст личинок | 22.05 | 26.05 | 4 | 3‒7 |
| Четвертый возраст личинок | 27.05 | 1.06 | 5 | 3‒7 |
| Куколка | 15.05 | 1.06 | 16 | 1‒5 |
| Всего дней в водной среде | 26.04 | 1.06 | 37 | 13‒33 |
| Первые нападения имаго | 03.05 | 10.06 | 38 | ‒ |
| Последние нападения имаго | 1.07 | 19.07 | 18 | ‒ |
| Всего дней в воздушной среде | 03.05 | 19.07 | 78 | 40 |
Периоды нападения на человека самок A. communis связаны со скоростью протекания весенних явлений, а именно характером весны: ранняя или поздняя, быстрая или затяжная. В разные годы отмечены различия по датам первых и последних нападений имаго на человека. Амплитуда температур при сборе имаго изменялась от 6 до 25 °С. Оптимальная температура, при которой отмечено максимальное число нападений имаго данного вида, составляла около 10 °С.
Продолжительность обнаружения исследуемого вида в водной и воздушной среде различалась. Нестабильность существования временного водоема ограничивала сроки нахождения преимагинальных стадий A. communis в водной среде по сравнению с воздушной. Имаго в природе нападали на человека более двух месяцев, тогда как преимагинальные стадии встречались в водоемах около месяца. Периодичность цикла развития комаров данного вида определялась отчасти погодными, в первую очередь температурными, условиями. Начало и окончание фенопериода смещалось по годам максимально на 4 недели (в 2011, 2021 гг.), минимально на 2 недели (в большинстве случаев).
Смена фенологических явлений связана со скоростью прогревания водоема, в котором происходит развитие личинок A. communis. Постоянное поступление талых вод действует охлаждающе на общую массу талой воды в водоеме и поддерживает низкие температуры. Холодоустойчивость A. communis способствует сокращению числа конкурентов и хищников на начальных этапах развития личинок данного вида во временных водоемах.
По нашим наблюдениям, в лесной зоне личинки A. communis чаще встречаются в затененных деревьями водоемах, где создаются благоприятные условия для их развития. Затенение исключает резкие суточные изменения температуры водной среды.
В разные годы в исследуемом водоеме развивались кровососущие комары преимущественно лесного и частично пойменного (Aedes cinereus) комплекса видов: в 2005–2009 гг. мы наблюдали развитие только A. communis; в 2010 г. водоем заселяли 6 видов (A. cinereus, A. communis, A. diantaeus, A. hexodontus, A. intrudens и A. pionips), в 2011 г. – A. communis и A. punctor, в 2012–2014 гг. – 4 вида (A. communis, A. punctor, A. diantaeus и A. intrudens). C 2015 по 2020 г. в водоеме развивались личинки A. communis, в 2021 г. были обнаружены A. communis и A. diantaeus, при этом по численности личинок преобладал A. diantaeus.
Обсуждение
Важной особенностью годовой ритмичности изученной выборки из популяции комара A. communis мы считаем ежегодно стабильно высокую относительную численность личинок младших возрастов в модельном водоеме как типичном месте развития данного вида в таежной зоне. Ежегодно в процессе таяния снега весной происходит неравномерное поступление в водоем холодных талых вод, охлаждающих водоем. Вместе с водой в водоем, в котором уже идет развитие личинок A. communis, добавляются новые партии перезимовавших яиц комаров с окружающих территорий, в результате чего в водоеме поддерживается разновозрастной состав преимагинальных стадий комаров. Личинки комаров дышат воздухом, при покрытии водоема льдом этот тип дыхания становится невозможным. Подо льдом при низких температурах (2 °С) происходит переход личинок на дыхание всей поверхностью тела, альтернативное воздушному дыханию (Тыщенко, 1986). Устойчивость личинок младших возрастов к понижению температуры воды и их выживание в толще воды подо льдом гарантируют сохранение части популяции при ее разновозрастном составе. В таежных биоценозах период развития личинок A. communis растянут во времени. Одной из причин может быть неравномерное поступление в водоем яиц комаров с талыми водами, что вызывает одновременное разновозрастное развитие личинок.
Заключение
Волны жизни массового в лесной зоне A. communis в период наблюдений с 2005 по 2021 г. имели два спада и один подъем. Пик относительной пищевой активности самок отмечался с 2013 по 2015 г., и его начало совпало с началом пика солнечной активности в 2013 г., с 2015 г. наблюдалось максимальное повышение плотности личинок данного вида после базового накопления числа яйцекладок за предыдущие годы. Таким образом, солнечная активность стимулировала пищевую активность самок комара A. communis, что подтверджено статистически (r = 0.56‒0.59).
Фенологические периоды повторяются ежегодно, периоды их наступления и продолжительность определяются внешними факторами. Период развития личинок в модельном водоеме составлял от двух недель до месяца в зависимости от температурных условий сезона и степени прогревания воды в водоеме. Нападение имаго самок A. communis на человека во многом зависит от времени наступления весенних явлений (характера и скорости протекания весны: ранняя или поздняя, быстрая или затяжная). В разные годы отличия по времени первых нападений на человека имаго составляли от двух до четырех недель. Продолжительность встречи имаго данного вида в природе превышает два месяца. Отмечена особенность фенологии A. communis – высокая устойчивость личинок младших возрастов к понижению температуры воды и их выживание в толще воды подо льдом. Разновозрастность популяции обеспечивает ее сохранение при пороговых для данного вида условиях среды.
Наблюдения за волнами жизни комара A. communis позволит делать успешные ежегодные прогнозы активности нападения кровососущих комаров для населения на текущий теплый период. Плотность личинок, а также связанная с ней активность нападения имаго на человека имеют достоверные связи с гелиоциклической активностью.
Библиография
Белецкий Е. Н., Станкевич С. В., Немерицкая Л. В. Современные представления о динамике популяций насекомых: прошлое, настоящее, будущее. Синергетический подход // Вісник Харківського національного аграрного університету. Серія «Фітопатологія та ентомологія». 2017. № 1−2. С. 22−33.
Бенкевич В. И. Солнечно-климатические ситуации и реализация вспышек массового размножения непарного шелкопряда Lymantria dispar L. (Lepidoptera, Lymantriidae) Московской обл. за 120 лет // Энтомологическое обозрение. 1990. Т. 69, № 1. С. 3−13.
Братцев В. М. Климат. Общая характеристика. Даты перехода среднесуточных температур воздуха через 0 °С весной и осенью // Атлас Республики Коми / Под ред. В. Н. Гайзер. М.: Феория, 2011а. С. 106.
Братцев В. М. Климат. Общая характеристика. Даты перехода среднесуточных температур воздуха через 5 °С весной и осенью // Атлас Республики Коми / Под ред. В. Н. Гайзер. М.: Феория, 2011б. C. 106.
Викторов Г. А. Проблемы динамики численности насекомых на примере вредной черепашки . М.: Наука, 1967. 271 с.
Виноградова Е. Б., Карпова С. Г. Сезонные и суточные ритмы кровососущих комаров . СПб.: Зоологический институт РАН, 2010. 237 с.
Витинский Ю. И. Солнечная активность . М.: Наука, 1983. 192 с.
Гиляров А. М. Популяционная экология . М.: Изд-во МГУ, 1990. 191 с.
Гуцевич А. В., Мончадский А. С., Штакельберг А. А. Комары (Семейство Culicidae). Фауна СССР. Насекомые двукрылые . Л.: Наука, 1970. Т. 3. Вып. 4. 384 с.
Гуцевич А. В., Дубицкий А. М. Новые виды комаров фауны Советского Союза // Паразитологический сборник Зоологического института РАН. 1981. Т. 30. С. 97–165.
Капусткина А. В., Хилевский В. А. Динамика численности и вредоносности вредной черепашки Eurygaster integriceps Put. (Heteroptera, Scutelleridae) в посевах зерновых культур степной зоны Предкавказья // Энтомологическое обозрение. 2020. Т. 99, № 1. С. 71–78.
Кривенко В. Г., Виноградов В. Г. Птицы водной среды и ритмы климата Северной Евразии . М.: Наука, 2008. 587 с.
Кункова Е. В., Федорова В. Г. Волны жизни доминирующей популяции комаров Aedes communis лесной зоны Новгородской области // Материалы XXVI межвузовской научно-практической конференции по проблемам биологии и медицинской паразитологии, посвященной памяти академика Е. Н. Павловского / Под ред. А. Ф. Никитина. СПб.: Изд-во Военно-медицинской академии им. С. М. Кирова, 1999. С. 63–64.
Лямцев Н. И. Динамика популяции непарного шелкопряда в лесостепных дубравах Европейской России // Лесоведение. 2019. № 5. С. 366–374.
Мартыненко В. А. Растительность и флора // Леса Республики Коми / Под ред. Г. М. Козубова, А. И. Таскаева. М.: ДиК, 1999. С. 54–61.
Медведев С. Г., Айбулатов С. В., Панюкова Е. В. Экологические особенности и распространение Aedes communis (De Geer, 1776) на территории Северо-Запада европейской части России // Паразитология. 2010. Т. 44, № 5. С. 441–460.
Николаева Н. В. Экология личинок кровососущих комаров Южного Ямала: Препринт / Под ред. Н. Н. Данилова. Свердловск: Институт экологии растений и животных, 1980. 66 с.
Николаева Н. В. Экология кровососущих комаров: популяционные и биоценологические аспекты // Экология. 1994. № 3. С. 60–69.
Остроушко Т. С., Панюкова Е. В., Пестов С. В. Двукрылые насекомые (Insecta: Diptera) комплекса Гнус фауны европейского Северо-Востока России // Труды Коми научного центра УрО РАН. № 183. Беспозвоночные европейского Северо-Востока России / Под ред. М. М. Долгина. Сыктывкар: Изд-во Коми научного центра УрО РАН, 2007. C. 190–235.
Панюкова Е. В., Грицай А. Б. Эпидемиологическая характеристика видового состава кровососущих комаров (Diptera, Culicidae) европейского северо-востока России // Материалы XVII Всероссийской конференции с международным участием «Биодиагностика состояния природных и природно-техногенных систем». Кн. 2 / Под ред. Т. Я. Ашихминой. Киров: Изд-во Вятского гос. ун-та, 2019. С. 356–359.
Панюкова Е. В., Мади Е. Г. Волны жизни доминирующей популяции кровососущего комара (Ochlerotatus communis De Geer, 1776) в подзоне средней тайги Республики Коми // Материалы XII Всероссийской конференции с международным участием «Биодиагностика состояния природных и природно-техногенных систем». Кн. 1 / Под ред. Т. Я. Ашихминой. Киров: ООО «Веси», 2014. С. 220–221.
Сазонова О. Н. Таблицы для определения самок комаров Aedes Mg. (Diptera, Culicidae) лесной зоны СССР // Энтомологическое обозрение. 1958. Т. 37, № 3. С. 741–752.
Станкевич С. В., Белецкий Е. Н., Забродина И. В. Циклически-нелинейная динамика природных систем и проблемы прогнозирования . Ванкувер: Accent Graphics Communications & Publishing, 2019. 232 c.
Стародубцев С. А. Космические лучи и турбулентность солнечного ветра: особенности 23-го цикла солнечной активности // Солнечно-земная физика. 2011. № 17. С. 86–91.
Тыщенко В. П. Физиология насекомых: Учебное пособие для студентов университетов, обучающихся по специальности «Биология» . М.: Высшая школа, 1986. 303 с.
Фролов А. Н. Луговой мотылек Loxostege sticticalis L. (Lepidoptera, Crambidae) в фокусе проблем сельскохозяйственной энтомологии. I. Периодичность массовых размножений // Энтомологическое обозрение. 2015. Т. 94, № 1. С. 3–16.
Фролов А. Н. Динамика численности и прогноз массовых размножений вредных насекомых: исторический экскурс и пути развития. Аналитический обзор // Вестник защиты растений. 2017. Т. 4, № 94. С. 5–21.
Халин А. В., Айбулатов С. В., Пржиборо А. А. Методы сбора двукрылых насекомых комплекса гнуса (Diptera: Culicidae, Simuliidae, Tabanidae) // Паразитология. 2021. Т. 55, № 2. С. 134–173.
Чернова Н. М., Былова А. М. Общая экология: Учебник для студентов педагогических вузов . М.: Дрофа, 2004. 416 с.
Четвериков С. С. Волны жизни. (Из лепидоптерологических наблюдений за лето 1903 г.). Повторное издание // Русский орнитологический журнал. 2009. Т. 18, № 519. C. 1822–1829. (Первая публикация: Известия Императорского общества любителей естествознания, антропологии и этнографии. Труды Зоологического общества. Дневник Зоологического отделения. 1905. Т. 3, № 6. С. 1–5.)
Ягодинский В. Н. Космический пульс биосферы . М.: Знание, 1975. 144 с.
Ягодинский В. Н. Космические циклы и ритмы жизни . М.: Знание, 1981. 64 с.
Becker N., Petric D., Zgomba M., Boase C., Madon M., Dahl C., Kaiser A. Mosquitoes and their Control. Second Edition. Heidelberg; Dordrecht; London; New York: Springer, 2010. 608 p.
Hammer O., Harper D. A. T., Pyan P. D. Paleontological statistics software package for education and data analysis // Palaentologia Electronica. 2001. Vol. 4, No 1. P. 1–9.
Panyukova E. V., Novakovskaya I. V., Patova E. N. Larval algae nutrition of Aedes communis (De Geer, 1776) // Russian Entomological Journal. 2018. Vol. 27, No 4. P. 443–449. DOI: 10.15298/rusentj.27.4.13.
Veronig A. M., Jain S., Podladchikova T., Pötzi W., Clette F. Hemispheric sunspot numbers 1874–2020 // Astronomy end Astrophysics. 2021. No 652. DOI: 10.1051/0004-6361/20214119/.
Благодарности
Автор благодарит к. б. н. И. И. Шуктомову и к. б. н. Э. И. Бознака (Институт биологии ФИЦ Коми научного центра УрО РАН, Сыктывкар) за помощь в подготовке рукописи к публикации, а также Е. А. Гутченко (отдел метеорологии филиала ФГБУ Северное УГМС «Центр гидрометеорологии и мониторинга окружающей среды Республики Коми», г. Сыктывкар) за любезно предоставленные метеорологические данные. Большое спасибо к. ф.-м. н. А. А. Абунину (Центр прогнозов космической погоды ИЗМИРАН, Москва) за консультации по современным исследованиям солнечной активности. Искренне благодарю д. б. н. Б. А. Коротяева и д. б. н. А. Н. Фролова за критические замечания и ценные рекомендации к рукописи статьи.
Исследование выполнено в рамках темы отдела экологии животных Института биологии Коми НЦ УрО РАН «Разнообразие фауны и пространственно-экологическая структура животного населения европейского северо-востока России и сопредельных территорий в условиях изменения окружающей среды и хозяйственного освоения» (Гос. регистрационный номер: 122040600025–2).

 © 2011 - 2026
© 2011 - 2026