


Introduction
Currently, commercial hunting of wild animals has finally given way hunting trophy shooting. Hunting trophy tourism, especially in the CIS countries, is one of the most intensively developing and prestigious forms of using of natural resources and, as a branch of human economic activity, requires a rational approach and should be governed by scientific data on the number and gender-age structure of the trophy species populations. One of the most valuable and desirable objects of trophy hunting are various species of ungulates, including mountain sheep, which occupy one of the leading places.
This study, carried out in the conditions of the Kazakh Upland, is a part of a series studies on the state of populations of separate subspecies of mountain sheep in the Middle and Central Asia, where trophy hunting for argali is practiced. The practical focus of the research was determined by the need to develop a long-term program for rational use and management of their populations under the growing influence of various forms of human impact on the population. Along with practical purposes, the data obtained express theoretical aspect and are necessary for understanding the features of formation and functioning of separate components of population structures, that was practically not done for such rare and poorly understood species as mountain sheep. It is known that structural organization of the population is formed under the influence of various kinds of external and internal influences and reflects an integrated response of the population to these effects. Therefore, it acts as one of the most important general population adaptations providing the optimal functioning mode and ultimately, the stability of populations in complex and dynamical environmental conditions.
It is the sex-age and spatial-temporal structure of animal populations and their dynamics amid the environmental changes is the least studied aspect of populational regulations. There is not a single generalized summary in the ecological literature on this topic, even within the single class of animals. Some authors on the example of separate species considered only the interdependence of changes in the sexual composition of populations and their numbers, often with very contradictory results.
The overall goal of our work was to assess the current state and identify patterns of the dynamics of structural and functional organization of the Kazakh argali population depending on spatial-temporal dynamics of environmental factors, as well as various forms and intensities of anthropogenic impacts.
Structural-functional organization of a population is understood by us as a relatively stable in time ratio of sex and age groups in a population, distributed in a certain way in space and interconnected into a single functional whole (Magomedov et al., 2001). As parameters of the structural and functional organization of the Kazakhstan argali population, were considered the features of changing in size and composition of separate herds of population groups, the spatial-temporal type of the territory use, and also sexual and age composition and characteristics of changes in these and other parameters of populations in depending on the specific environmental conditions.
Materials
The sheep inhabiting the Kazakh Upland are considered by most researchers to be the Kazakhstan or North Kazakhstan form of argali (Ovis ammon collium, Severtzov, 1873) (Calkin , 1951; Berber, Kalmykov, 1994; Baydavletov, 1996; Bekenov et al., 1999; Fedosenko, 2000). Kazakhstani argali lives in Central and Eastern Kazakhstan, where it occupies most of the Kazakh highlands, in the south it goes to the Bektaut mountains and Katan-Emel (northern Balkhash), to the east, in Semipalatinsk region, it is common in the Karachingiz mountains, Akchatau and others. In Eastern Kazakhstan, it is found in Kalbinsk Altai, Tarbagatai and Saur. Its range also extends beyond Kazakhstan.
The areas of our research were in the central part of the Kazakh Upland (Central Kazakhstan Upland) in the territory of the Karaganda and partially Semipalitinsk regions. The field material collection plots covered the mountains of Kyzyltau, Kalmakkyrgan, Arkalyk, Edrey, Myrzhik. The climate is sharply continental, expressed in the severity of winter, high summer temperatures, the short duration of spring and autumn, large annual and daily amplitudes of air temperature, dry air, and low precipitation (Al'pat'ev et al., 1976). Rainfall varies greatly from year to year, averaging around 300 mm per year. The best moisturization is in the mountain ranges, on which about 400 mm of precipitation falls during the year. The main precipitation falls on the summer period (70–80%), therefore winters are with predominantly light amount of snow (about 20–25 cm). The winter period is characterized by frequent and severe snowstorms (up to 40 days) (Gvozdeckiy, Mihaylov, 1978).
Fescue - feather grass steppes are distributed predominantly, in some places xerophytic feather grass (Stipa sareptana), June grass (Koeleria gracilis), oat-grass (Helictotrichon desertorum) can play a greater role in the grass stand. In the spring, ephemera and ephemeroids develop, and thickets of steppe shrubs – peashrub and spirea – are found along depressions and ravine (Gvozdeckiy, Mihaylov, 1978). In the mountain ranges of the Upland, altitudinal zonality appears. Differences in herbage are observed depending on the exposure of the slope. On the northern slopes the motley grass are well represented, while on the southern slopes – the more xerophytic species with narrow and rather hard leaves.
Due to the strong decline in livestock numbers in recent years, the pasture load on vegetation in the research area is poorly expressed. As a consequence of this is the accumulation of junk and steppe fires, often covering vast territories. Fires strongly affect the forage base of argali in the winter, the most severe in the forage relation period. Judging by the polling data, poaching is also flourishing in this territory.
During the research period (November 4–18, 2002), 449 argali individuals were visually observed on an area of 1544 km2.
Methods
For a number of reasons, the route method of accounting for numbers with using cars was chosen as the main. Animal were counted in daylight at the same time in three cars on predefined daily routes. In a number of cases, in parallel the accounting sites were used, which were laid every 3-5 km along the main line of the route (transects).
The total number of counted animals and their gender and age groups, places of their detection, direction of movement and time of detection were plotted on a plan or large-scale map of the area and were fixed in the memory of electronic media.
When studying the sexual and age structure of populations, females, males and yearlings of both sexes were distinguished. Certain difficulties arose in determining of one and a half year old individuals; at this age, males and females can be very difficultly distinguished from a distance. Therefore, when collecting empirical material, we conditionally attribute them to females, and later they separate while the analysis of the demographic structure. Due to the fact that the horns of females are small in size and poorly visible at a distance, empirical data on the number of animals of older ages were obtained only for males. Thanks to the annual rings on the horn covers and the shape of the horns, the age of the males, even in nature, was determined with an accuracy of one year.
To determine the age structure of the population, the horns of dead animals were also used. From the rings on the horn covers, it is possible to determine accurately not only the age of males, but also females, and even the time of year when the animal died. We found and examined 37 skulls of males and only 2 skulls of females. Also was used the material on dead animals (skulls of males), which is kept in the museum of the Academy of Sciences of Kazakhstan (74 skulls). This method is widely used in the analysis of the age structure of bovids (Caughley, 1979; Riklefs, 1979; Pianka, 1981). In addition, we obtained material on the ages of the animals shot (a total of 126 animals, 2 from which were females), but we did not use it in the analysis of the age structure, since the shooting is selective in sex and age and therefore does not reflect the real demographic structure in the population. An insignificant number of skulls of females and young, apparently, is due to the fact that their lighter skulls are pulled apart by predators.
Alignment of age rows and construction of the age distribution in the argali population (by sex and age) were calculated on the basis of special logarithmic-polyminal and integral equations (Ahmedov, Magomedov, 2000; Magomedov et al., 2001).
The spatial-temporal structure and nature of the territory use by the argali population was studied on the basis of material obtained from the analysis of animal meetings depending on the nature of the habitat. Altitude, orientation of the slopes to the sun, as well as individual relief elements – valleys, edges of valleys and foots of slopes, the slopes themselves and plateau-like elevations with crests of ridges were considered as separate characters of habitats.
Data on the general nature of the relief, the degree of its dissection (the number of ravines per area unit), the average steepness of the slopes, their length, the ratio of aligned (flat areas) and rugged slopes, their relative position, etc., were used as criteria for the protective properties of sheep habitats.
Results
As was shown by our research, the distribution of argali on the territory was relatively uniform. The population density for separate mountain ranges did not differ significantly and fluctuated within 0.24–0.35 individuals per 1 km2, averaging 0.28 ± 0.02 individuals per 1 km2 (table. 1).
Table 1. Population densities of argali at research sites
| № | Research sites (mountain range) | Number of marked animals, (n) | Area of accounting, (km2) | Population density, |
| 1 | Kyzyltau | 101 | 315 | 0,32 |
| 2 | Kalmakkypgan | 48 | 184 | 0,26 |
| 3 | Arkalyk | 25 | 103 | 0,24 |
| 4 | Erdey | 96 | 342 | 0,28 |
| 5 | Myrzhik | 74 | 296 | 0,25 |
| 6 | Konyrtemyrshi | 105 | 304 | 0,35 |
| Total | 449 | 1544 | 0,28 |
Nevertheless, the distribution of animals in the territory revealed certain patterns. Although argali was also noted by us in open completely even intermountain valleys during the research period, nevertheless most animals adhered to uplands and rocks outcrops, i.e., territories with dissected relief. Direct dependences of argali density on the area and height of mountain massifs are noted (Fig. 1). The larger the area of the mountain range and the higher it is, the greater the length and steepness of the slopes and the more pronounced network of ravines are. Obviously, this is due to the fact that the argali find the most optimal feeding and protective conditions on the crossed territory.

Fig. 1. The dependence of the number of marked animals on the area of the mountain range (r = 0.97; P < 0.05) (A) and altitude (r = 0.75; P > 0.05) (B). The X-axis: A – the area of the mountain range (km2); B – markы of the absolute heights of the mountain range (m); the Y-axis: A – the number of marked animals (individuals); B – the population density of argali (individuals/km2)
The noted dependences of the number and density on the area of the mountain massif and the degree of partition of the territory, in addition to the feeding factor, are largely determined by the large possibilities of protection from predators and humans. On more dissected and larger mountain formations, i.e., less passable for vehicles, the population number is higher than in areas more accessible to humans. The role of rugged terrain as protective conditions of the sheep’s habitat is also indicatet by the behavior of animals, which in danger always rushed to the nearest mountain ranges.
An integral part of the social structure of ungulates, which largely determines the type and effectiveness of the population’s functioning is the herdness. In general, the herd index in the argali population on the territory of research during the rut period was 4.24 ± 0.30. In general, the herdness index in the argali population in the research area during the rut period was 4.24 ± 0.30. The largest number of animals in the research period was concentrated in harems (39.2 %), then in mixed (31.4%) and female groups (23.4%). A high herdness index is characteristic for mixed groups, and for female groups and harems it is much lower (table. 2).
Table 2. Types of argali groups and their herdness indices
| Groups type | The number of groups | The number of individuals | The herdness index | ||
| N | % | n | % | ||
| Loners | 25 | 23.6 | 25 | 5.6 | 1.0 |
| Females | 24 | 22.6 | 105 | 23.4 | 4.38±0.61 |
| Males | 1 | 0.9 | 2 | 0.4 | 2.0 |
| Harems | 36 | 34.0 | 176 | 39.2 | 4.89±0.38 |
| Mixed | 20 | 18.9 | 141 | 31.4 | 6.81±0.72 |
| Total | 106 | 449 | 4.24±0.30 | ||
The largest number of animals was concentrated in groups ranging in size from 3 to 8 individuals. In general, 65.7% of the animal population are kept in groups of this size. The largest group of argali we noted in this territory consisted of 17 animals (Fig. 2).

Fig. 2. The distribution of Kazakhstan argali into groups depending on group sizes. The X-axis – the size of groups; the Y-axis – the number of individuals in groups
It is shown that among the many possible factors, the herdness index depends on two parameters: the height of the mountain massif above sea level and the relative number of males in the population. With an increase in the height of the massif, which indirectly reflects the protective properties of the habitat, the herdness index regularly decreases (r = -0.85; P <0.05), which is typical for many mountain ungulates (Wilson, 1975; Баскин, 1976; Коли, 1979; Дарман, 1990). Another parameter affecting the argali herdness index is the relative number of males in the population: during the rutting season, males separate females from the main groups and form harems characterized by low herdness indices. The single males remaining without harems also reduce the overall herdness index (Fig. 3).
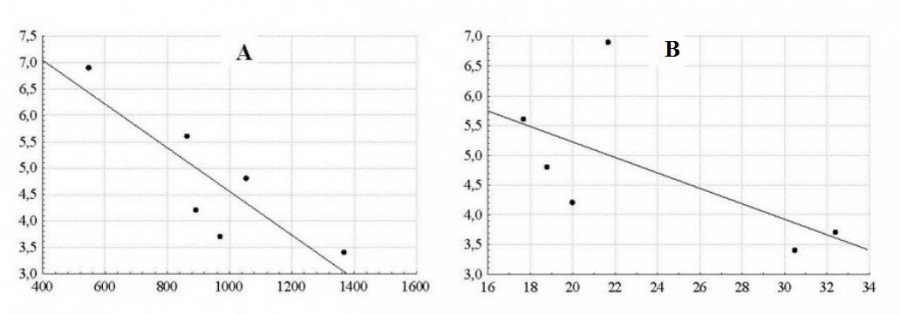
Fig. 3. The dependence of Kazakhstan argali herd population index on habitat height (A): Y = 8.701 - 0.004x; (r = -0.85; P < 0.05) and relative number of males in the population (B): Y = 7 822 - 0.13 x; (r = -0.63; P > 0.05). The X-axis: A – altitude (meters), B – the relative number of males in the population (%); the Y-axis – herd index
It is known that argali, like most mountain ungulates, are characterized by age and gender differences in the use of the territory, which are most pronounced in the summer. The period of our research coincided with the rutting period. And at this time, most animals are in harems and mixed groups, and males and females practically do not differ in the nature of the territory use. In the study area, almost the entire argali population was kept on uplands – 412 individuals (91.8%), in the intermountain valleys only 37 individuals were recorded (8.2%). Of all the animals noted in the uplands, the majority were found on the mountain’s slopes (78.6%), much less in the intermountain valleys (17.5%) and very few on the ridges of the hills (3.9%).
On the slopes of mountain ranges (uplands), we noted 324 argali, of which 43.8% adhered to the northern slopes, 29.0% to the slopes of the eastern exposure, 14.5 and 12.7% respectively to the western and southern slopes. The argali preference for the northern slopes, apparently, may be explained by the dominance of mesophytic plant species, well eaten by sheeps.
Argali were found in the altitude range from 400 to 1300 m. at. m., where their main population (83.3%) prefer an altitude of 550-850 m. at. m., and the maximum number of met animals was at a height of 650 m n. at. m. (19.6%). These heights correspond to the middle part of the slopes and occupy the largest square in the research area.
Thus, at the beginning of the winter season, the most preferred by argali plots are the middle parts of the northern slopes of mountain ranges. The most important factors determining the spatial population structure are the habitats’ orographic conditions, the distribution character of vegetation cover, as well as the meteorological conditions of this period.
The most important population characters of animals are their sex and age structure, reflecting the current state of the species as a whole and its separate populations, the intensity of their reproduction and mortality for different age groups. As in many other species, and especially in ungulates, it also acts as the main populational parameter, which must be taken into account during its exploitation. Therefore, when conducting research, we tried to pay special attention to the age and sex composition of the studied populations and their groups.
Unfortunately, during the research period argali rush had already begun, therefore both males and female groups could be met on the same territory, but most of the noted groups were mixed.
As can be seen from the table. 3, adult females prevail in argali populations on almost all mountain ranges in the central part of the Kazakh Upland; they make up from 36.0 to 52.1% of the entire population (average 44.28 ± 3.07). The number of males older than 2 years and yearlings is approximately equal (23.52 ± 2.58% and 21.00 ± 3.82%, respectively). The smallest age and sex group are one-year-old animals (9.33 ± 0.62%). The sex ratio among adults (over 2 years old) is 1.9: 1.0 in favor of females, i.e. in a population there is on average about 2 females per male. The proportion of yearlings per female is generally 0.43.
Table 3. Sex and age structure of argali population in separate mountain ranges of the central part of the Kazakh Upland
| Research sites (mountain range) | Females | Yearlings | One-year-old | Males | ||||
| n | % | n | % | N | % | n | % | |
| 1. Kyzyltau | 52 | 51.1 | 19 | 18.8 | 11 | 10.9 | 19 | 18.8 |
| 2. Kalmakkypgan | 18 | 37.5 | 13 | 21.7 | 4 | 8.3 | 13 | 21.7 |
| 3. Erdey | 9 | 36.0 | 9 | 36.0 | 2 | 8.0 | 5 | 20.0 |
| 4. Arkalyk | 50 | 52.1 | 19 | 19.8 | 10 | 10.4 | 17 | 17/7 |
| 5. Myrzhik | 37 | 50.0 | 5 | 6.8 | 8 | 10.8 | 24 | 32/4 |
| 6. Konyrtemyrshi | 41 | 39.0 | 24 | 22.9 | 8 | 7.6 | 32 | 30.5 |
Analysis of the sex and age structure of populations depending on the height of the massif above the level and on the total area of the mountain massif revealed some relationships between these parameters. The relative number of females and one-year-olds shows a positive relationship with the area of the mountain massif (r = 0.80; P> 0.05 for females; r = 0.62; P> 0.05 for one-year-olds), while the number of yearlings is negative (r = -0.70; P> 0.05). Thus, during the first year of life on such massifs, it is slightly higher (r = 0.40; P> 0.05).
Although sex and age segregation of females and males is a widespread phenomenon, a weak manifestation of the patterns noted above is most likely associated with the rutting season when mountain sheep males roam the territory and show no attachment to a certain territory.
To analyze the demographic structure of the population, we used materials on the skulls of dead animals (111 skulls of males aged 1 to 11 years) and visual observations. The analysis shows that the proportion of males in the population begins to decrease sharply from the 4 age and disappears up to 15 years, while the decrease in the proportion of females in the populations is much slower, and females of 18 years old are found in the population (Fig. 4).

Fig. 4. Age pyramid of the population of Kazakhstan argali. The X-axis – the number of animals, %; the Y-axis –age (years)
The noted differences in the distribution of females and males by age clusters are the result of differences in their specific mortality at different age periods (Fig. 5). Relatively high specific mortality rates are characteristic for yearlings and animals of the older age groups, the lowest - for young animals aged 1–4 years. Higher rates of specific mortality of males older than 3 years are distinguished, which can be explained by the predominant legal or illegal shooting of males by hunters. According to interrogation data, at least 100 animals per year from 7 to 14 years old are hunted in this territory during trophy hunting.

Fig. 5. Specific mortality of females and males in argali populations depending on age in the Central part of the Kazakh upland. X-axis – specific mortality; Y-axis – age (years)
Conclusions
Due to the strong reduction in livestock over the past years, the pasture load on the vegetation in the research area is practically not expressed. Apparently, steppe fires, often spanning vast territories and depriving animals of winter food, already have a significant impact on argali populations.
One of the main factors limiting the number of populations undoubtedly is the poaching of animals. The degree of influence of poaching on the population size depends on the orographic features of the habitats that determine the protective conditions of the territory. It must be said that the evolutionary confinement of animals to territories with protective conditions was formed as a response to the activity of predators. In modern conditions, although the predation factor continues to retain a certain significance in the distribution of animals, anthropogenic factors, namely poaching, come to the fore. These two reasons of animals’ death have much in common in the sense that the degree of their influence on the population is directly determined by the degree of the territory protection. However, while predators attack, even small protective elements of landscapes (separate hills, rocks, or even rocky outcrops) allow animals to avoid death, then when exposed to humans, more stringent requirements are imposed on protective conditions (remoteness from settlements and roads, impassability for humans and vehicles, good visibility, etc.). It should also be noted that the nature of elimination of ungulate animals when exposed by predators is more “accessible”, most often due to the fact that predators remove defective, sick, or old individuals from the population. During poaching, there is poor selectivity and clear selectivity for large males stallions during trophy hunting.
The Kazakh argali population studied by us, by its nature and the nature of the occupied habitats has always seemed to be the most vulnerable, compared to other subspecies of Central Asian mountain sheep, where hunters plays a large role in the restriction of their abundance.
According to the literature, the number of argali within the Kazakh Uplands until the mid-1960s. was high and stable and amounted to about 16,000 individuals. Then the population began to decline, and up to 1970 it had decreased by more than half (about 7000 individuals) (Savinov, 1974). The process of population decline continued in subsequent years. In the 1980s there were already 5,000 individuals (Fedosenko, Kapitonov, 1983). The first half of the 1990s also was characterized by a decrease in numbers (2082–2428), and by the end of the 1990s. it barely exceeded 1000 animals (1997), although in some areas the density continued to remain high (Bekenov, Baydavletov, 1997; Bekenov et al., 1999; Fedosenko, 2000). A sharp decrease in the number of Kazakhstan argali from the second half of the 1960s to the beginning of the 1990s. was associated exclusively with human economic activity – the intensification of agriculture in this region (the development of virgin lands and the increase in the number of domestic animals). After the 1990s – an unprecedented increase in poaching against the background of the collapse of the USSR and the loss of law obedience, the difficult material and financial situation of environmental organizations, and the emergence of highly effective rifled weapons among the people. Factors such as a sharp increase in the number of wolves, annual steppe fires depriving sheep of winter food, conducting trophy hunts during the rutting season, began to play an increasingly noticeable role.
All this is confirmed in our studies: a clear positive relationship was noted between the number and density of argali, on the one hand, and the area of the mountain massif and the degree of dissection of the territory, on the other. In more dissected and larger mountain ranges, i.e., less passable for vehicles, the population is higher than in more accessible for humans areas. The role of the rugged relief as protective conditions for the habitats of mountain sheep is also indicated by the behavior of animals: although we met animals in the open steppe, seeing us, they always rushed to the nearest mountain ranges.
In the argali population, almost in all mountain ranges in the central part of the Kazakh Upland the adult females which make up one third to half of the entire population dominate. The number of males older than 2 years on average did not exceed 25% of the entire population. The proportion of males begins to decline sharply from 4 years old and fades to 15 years. The natural determining cause of changes in the population demographic structure is the feeding conditions of the winter and summer periods. We have previously shown that summer feeding conditions determine the fatness of ungulates to the beginning of the winter period (Ахмедов, Магомедов, 1995, 2000). Males during the rutting season, which in most species of ungulates takes place at the beginning of winter, lose almost all the fat reserves accumulated in the summer-autumn period. As a result, their survival is completely dependent on the availability of food in the winter. Females practically do not lose fat reserves during the rut, and their survival in the winter period depends on the feeding conditions of not only the winter, but also the summer period. So, in winter, when the availability and nutritional value of feed is sharply reduced, the survival rate of males is lower than that of more fat females. An additional confirmation of this is the decrease in the proportion of males from south to north. Due to the deeper snow cover, the availability of food in the winter is lower on the northern regions of the range than in the southern. We believe that in mountain sheep this is one of the reasons that determine the age and sex structure of the population. The consequences of steppe fires superimpose on this, after that large areas are practically unsuitable for wintering of animals. Fires in the northern regions cover large areas due to the better development of steppe vegetation here. In addition, leaving the burned places, animals are forced to use areas that have been preserved for winter pastures of domestic animals and where, in addition to competing with domestic sheep, they become more accessible to wolves and hunters.
According to the literature, by the 2000s. the wolf’s role in argali mortality on the territory of the Kazakh Upland increased significantly (Bekenov, Baydavletov, 1997; Bekenov et al., 1999; Fedosenko, 2000). According to A.K. Fedosenko (Fedosenko, 2000), the share of wolves in the mortality of argali reach from 40 to 73% of all dead animals.
In the territory under consideration, trophy shooting of animals is carried out, in the period from 1990 to 2001, 124 males from 7 to 14 years old were shot. Thus, along with the availability and seasonal dynamics of feed conditions (fires, grazing, climatic conditions, pressure of wolves, etc.), selective trophy shooting of animals acts as a new factor influencing the populations’ demographic structure.
References
Ahmedov E. G., Magomedov M. Dependence of the dynamics of the sex-age structure of the populations of the Dagestan tour (Capra cylindricornis Blyth) on the living conditions and population density, Zoologicheskiy zhurnal. 1995. T. 74. Vyp. 7. P. 109–118.
Ahmedov E. G. Magomedov M. Patterns of formation of the demographic structure of the population of the Dagestan tour (Capra cylindricornis), Zoologicheskiy zhurnal. 2000. T. 79. Vyp. 4. P. 461–470.
Al'pat'ev A. M., Arhangel'skiy A. M., Podoplelov N. Ya., Stepanov A. Ya. Physical Geography of the USSR (Asian part). M.: Vyssh. shk., 1976. 357 p.
Baskin L. M. The behavior of ungulates. M.: Nauka, 1976. 293 p.
Baydavletov R. Zh. The current state of argali population in Central Kazakhstan, Sostoyanie teriofauny v Rossii i blizhnem zarubezh'e. M., 1996. P. 21–26.
Bekenov A. B., Baydavletov R. Zh., Fedosenko A. K. Veynberg P. I. On the state of argali population in the Karaganda region, Problemy ohrany i ustoychivogo ispol'zovaniya bioraznoobraziya zhivotnogo mira Kazahstana. Almaty, 1999. P. 13–14.
Bekenov A. B., Baydavletov R. Zh. Scientific experiment "argali", Redkie vidy mlekopitayuschih Rossii i sopredel'nyh stran. M., 1997. P. 10.
Berber A. P., Kalmykov I. B. Arkhar in the Karaganda region, Ohota i ohotnich'e hozyaystvo. 1994. No. 11. P. 12–14.
Calkin V. I. Mountain sheep of Europe and Asia. M.: Izd-vo MOIP, 1951. 343 p.
Darman Yu. A. Flocking of roe deer as a population indicator, V s'ezd Vsesoyuznogo teriologicheskogo obschestva AN SSSR (29 yanv. – 2 fevr. 1990 g., Moskva): Tez. dokl. M., 1990. T. 2. P. 141–142.
Fedosenko A. K., Kapitonov V. I. Arkhar, Mlekopitayuschie Kazahstana. Alma-Ata, 1983. T. 3. Ch. 3. P. 144–209.
Fedosenko A. K. Arkhar in Russia and neighboring countries. M., 2000. 291 p.
Gvozdeckiy N. A., Mihaylov N. I. Physical Geography of the USSR (Asian part). M.: Mysl', 1978. 508 p.
Koli G. Analysis of vertebrate populations. M.: Mir, 1979. 362 p.
Magomedov M., Ahmedov E. G., Yarovenko Yu. A. Dagestan tour (population and trophic aspects of ecology). M.: Nauka, 2001. 138 p.
Pianka E. Evolutionary Ecology. M.: Mir, 1981. 396 p.
Riklefs R. Fundamentals of General Ecology. M.: Mir, 1979. 424 p.
Savinov E. F. Results of aerial survey of argali in the Kazakh Highlands, Sovremennoe sostoyanie i puti razvitiya ohotovedcheskoy nauki v SSSR. Kirov, 1974. P. 168–169.
Wilson E. O. Sociobiology: The new synthesis. Cambridge: Harvard Univ. Press, 1975. 696 p.

 © 2011 - 2026
© 2011 - 2026