


Researches on the thermobiology of reptiles began abroad in the first half of the 19th century. Snakes, lizards, turtles and crocodiles abundantly inhabited (and now, of course, also inhabit) tropical countries (in the 19th century it was most often colonies), in which these studies were originally carried out by scientists and simply interested people (both of them are mostly representatives of the metropolises). In addition, among the reptiles of the tropics, many those animals are really dangerous for humans (a lot of poisonous snakes, dangerous large boas and pythons, crocodiles, etc.). Therefore, especially earlier, they played a serious role in people's life and a lot of attention was paid to them. This attention, activated, probably, initially to a large extent by fear, later grew into curiosity, and then into scientific interest.
STUDY OF THERMOBIOLOGY OF REPTILES IN FOREIGN COUNTRIES
The first work we know about the heat balance of reptiles was published in the first half of the 19th century (Valensiennes, 1841). In this article it was said that incubating python female significantly increase their body temperature, especially in the place where they cover the clutch. The next publication on the same subject was published a year later (Lamarre-Pacquot, 1842). But the earliest oral report by this author to the French Academy of Sciences that incubating female pythons in Bengal increase their body temperature by several degrees took place as early as 1832.
The first concrete mention about the reptiles’ body temperature, which we know, dates from the middle of the 19th century (Agassiz, Gould, 1856). At the end of the XIX century some scientists began to use thermocouples to measure body temperature (Sutherlanad, 1896). This expanded the possibilities of field and laboratory studies. Observations on behavior associated with the use of heat and with the thermal endurance of reptiles have been published since the end of the 19th century (Werner, 1891; Richet, 1889; Soetbeer, 1898).
The accumulation of primary data in the same direction appeared from the very beginning of the 20th century (Gadow, 1901; Grinnell, 1908; Camp, 1916; Weese, 1917, 1919) and continued until the end of the 30s. (Buxton, 1923, 1924; Hall, 1922; Reese, 1923; Hesse, 1924; Parker, 1925; Kanitz, 1925; Baldwin, 1925a, b; Kammerer, 1926; Pearse, Hall, 1928; Krüger, 1924, 1929; Krüger , Kern, 1924; Weigmann, 1929; Franz, 1930; Pearse, 1931; Benedict, 1932; Blum, Spealman, 1933; Mosauer, Lazier, 1933; Mcilhenny, 1935; Parker, 1935; Mosauer, 1930, 1936; Lufti, 1936 ; Warden et al., 1936; Hesse et al., 1937; Atsatt, 1939; Bogert, 1939; Cowles, 1939; Klauber, 1939). Moreover, the number of publications in the 30s. beginning to increase significantly. If from the middle of the XIX to the end of the 20s. XX century we know only 31 publications, then only for the 30s. – already 39 (figure). In the late 30s. scientists who later become classics in the field of thermobiology appear on the list of authors – these are R. B. Cowles and C. M. Bogert.
Publications, in one way or another related to the thermal biology of reptiles, were associated with a description of the ecology of these animals, their living conditions, behavioral reactions aimed to avoid the temperature death from overheating, choosing the most appropriate and safe microbiotopes, activity time, etc. But most of the materials dealt with the temperature endurance of reptiles. Probably, this was due to the fact that many of the first observations on the biology of reptiles were carried out in places where they were most numerous and where they were visually more accessible – in deserts. Researchers were very impressed that they met primarily lizards and sometimes snakes a) at very high ambient temperatures and/or b) in places where such temperatures were periodically present. Moreover, paragraph a was often mistakenly identified with paragraph b. Nevertheless, this is the first thing that caught the eye, sometimes caused bewilderment and, therefore, scientific interest. and experiments on the studying of temperature endurance could initially be put even at the most primitive way, which does not require serious equipment and scientific knowledge (for example: in how many minutes a snake, placed on the ground with a temperature of 40°, 42°, etc., dies).

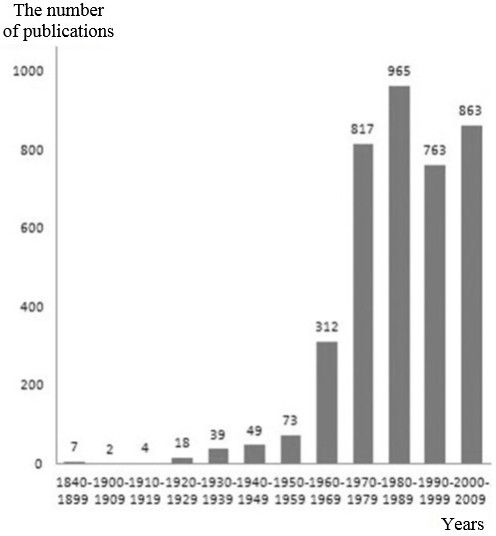
The number of known to us foreign scientific publications concerning the thermal biology of reptiles and the nearest adjacent areas, by years
However, already in this initial period some single articles appeared that showed the attempts to understand the value of temperature in animals’ life, the ratio of ecto- and endothermy (Pearse, Hall, 1928; Benedict, 1932; Warden et al., 1936, etc.).
Thus, from the middle of the XIX century. to the end of the 30s. XX century in foreign studies of reptiles’ thermal biology, there was a period of primary accumulation of scientific material according to the most striking reptile biology-related features associated with temperature.
Publications of the 1940s (we know 49 of them, see the figure) demonstrated the desire of researchers to penetrate deeper into the essence of the reptiles’ biology. In addition to ecology topics that have already become commonplace (Lueth, 1941; Lowe, 1942; Cowles, 1944; Bogert, 1949a, b, etc.) and thermal endurance (Cowles, 1940, 1942, 1945; Cole, 1943; Cowles, Burleson, 1945; Colbert et al., 1946; Woodbury, Hardy, 1948 and others), there were works devoted to other important aspects: wintering of reptiles and thermal endurance at low temperatures (Cowles, 1941; Bailey, 1948, 1949), the influence of temperature on the development of morphological characters in snakes (Klauber, 1941; Fox, 1948), general problems of thermoregulation (Herter, 1941; Cowles, 1942; Bogert, 1949a, b).
But, of course, there is the article devoted to general questions of reptiles’ thermal biology that is probably of particular importance (Cowles, Bogert, 1944). By most herpetologists, it is considered to be the first serious work on the thermal biology of reptiles, which began the development of this scientific course. In it, the authors showed the results of rather detailed studies of thermal biology of twelve species of diurnal reptile and seven species of nocturnal reptiles (lizards and snakes) in the southwestern United States. They clearly indicated (unfortunately only in passing) a number of very important aspects that unfortunately were not noticed by their scientific followers. So, they determined the state of “normal activity”, although they did not focus their attention on this important methodological aspect. In fact, this activity state is fully consistent with the state of “full activity [1]”, which was described a year earlier by S. S. Liberman and N. V. Pokrovskaya (1943), although R. B. Cole and K. M. Bogert were clearly not yet familiar with the last article. In addition, R. B. Cole and K. M. Bogert clearly demonstrated (although, alas, they did not focus on this) that data on activity temperatures, and especially on preferred temperatures, should be allocated only from samples of body temperatures at normal activity. This is extremely important, because if you do not take into account the circumstances mentioned above, then the researchers will receive incorrect, erroneous primary data, and this, in turn, will entail errors in the analysis of the material and in the conclusions regarding even the most common (physiological, evolutionary) problems (Черлин, 2013, 2014a; Cherlin, 2015a).
As a result, the thermal biological characters of reptiles’ activity and thermal preference showed by R. B. Cole and K. M. Bogert in 1944, turned out to be methodically more correctly determined (both in sense and in numerical values) than in most of the researchers who followed them. In other words, the publication of R. B. Cole and K. M. Bogert (Cowles, Bogert, 1944) has much more scientific, but primarily – methodological significance than is now commonly believed. However, it is not the first important article on reptile thermal biology. This primacy should rightfully belong to the works of I. D. Strelnikov (1934, and others), as well as S. S. Liberman and N. V. Pokrovskaya (1943).
Publications of the 50s (we know 73 of them, see the figure) had a different scientific orientation: there were studies on the ecology and thermoregulatory behavior of reptiles (Carpenter, 1952; Norris, 1953; Pearson, 1954; Fitch, 1954; Cowles, 1956; Inger, 1959 and others.), in terms of their body temperature in nature and its dynamics (Bogert, 1953; Carpenter, 1956; Fitch, 1956), on thermal physiology and thermal endurance (Lowe, Vance, 1955; Parker, 1956; Reichling, 1957; Cowles, 1958b; Evans, Cowles, 1959; Musacchia, 1959), on thermoregulation (Saint-Girons, Saint-Girons, 1956; Reichling, 1957; Cowles, 1958a; Bogert, 1959).
Thus, for the 40s and 50s. of XX century, the foreign countries are characterized by a gradual, slow increase in interest and the number of publications on reptiles’ thermal biology, intensification of instrumental component in research, especially in the 50s. The most general ideas about the connection of physiology and ecology of reptiles with temperature also began to form during this period. In foreign science, this was the time of the foundation of reptile's thermal biology as a new, developing science, basically a circle of problems was defined, a set of concepts, terms, methodology, methods, the first summaries and generalization attempts appeared.
Since the beginning of the 1960s, and especially in the 1970s. the number of publications on thermal biology of reptiles began to increase in an avalanche-like manner (we know 312 for the 60s, and 817 publications in the foreign scientific press for the 70s, see figure). This time is characterized by the manifestation of maximum interest in problems of thermal physiology (metabolism, energy metabolism, immunology, salt and water balances, food digestion, the importance of blood circulation in thermoregulation, etc.) (Bennett, 1972; Bennett, Ruben, 1979; Bennett et al., 2000; Bligh, Johnson, 1973; Pearson, Brandford, 1976, etc.), and then decreasing – in thermoregulation (the first serious works began to appear on thermoregulation in reptiles – Cowles, 1962; Brattstrom, 1965; McGinnis, Moore, 1969 and others, as well as the principles of organizing thermoregulation, which are sometimes described by different mathematical models – Hardy, 1965 ; Heath, 1970; Berk, Heath, 1975; Barber, 1977, 1979; Barber, Crawford, 1977), and in thermoregulatory behavior, body temperatures in nature and adaptive ecology of different species of reptiles, reproduction, temperature endurance and research methodology. It was during this period that the peak of the scientific activity of the remarkable American scientist Paul Licht fell. He studied the effect of temperature on various physiological systems of reptiles, the responses of the endocrine system to temperatures during the regulation of various body functions, the interaction of various external factors on activity, digestion of food, reproductive functions, etc., etc. Paul Licht made an invaluable contribution to the development of our knowledge and understanding of ecological physiology, physiological ecology, ecology and other aspects of the reptiles’ life, their adaptations to the environment, etc.
Thus, the 1960s and 1970s. in foreign countries have become, first of all, a time of deepening of knowledge about the relationship of internal and external, physiological and environmental components in reptiles’ thermobiology.
In the 1980-1990s. foreign scientists showed a very high interest in reptiles’ thermal biology (for the 80s, we know 965, for the 90s – 763 publications in the foreign scientific press, see figure). Firstly, during this time a large number of specific materials of field and laboratory studies in the field of thermal ecology of species has been accumulated. Most likely, this is due to the fact that a large number of researchers in Western countries have the opportunity to make numerous expeditions to tropical regions. In addition, many new technical means and electronic devices have appeared that make it possible to study the body temperature of animals more quickly, simply and efficiently, use remote methods for recording activity and body temperature, etc. This has made it possible to increase the rate of accumulation of primary data on the thermal ecology and thermal physiology of certain reptile species, faster collecting materials for articles and dissertations and has become attractive to herpetologists.
Secondly, extensive materials have been accumulated on thermal physiology, thermoregulation, and thermal biological indicators (Nelson et al., 1984 and others). Much and serious attention was paid to metabolism and energy, various problems of thermal physiology (the effect of temperature on different physiological functions, etc.), thermal ecology and thermoregulation (electronic technical means of tracking and control began to be actively used), the effect of incubation temperature on the sex of hatchlings, etc. Here also, probably, a great role was played by the appearance of new, modern technical tools. In addition, due to the accumulation of a large number of results of specific studies, during this period it became possible to begin a real analysis of these materials: comparing the thermal biological parameters of reptiles from different natural and climatic regions, thermal physiological characteristics, activity structures, thermal ecological features, options for organizing of thermoregulation, some generalizations on the evolution of the thermobiological sphere. Over these years, the thermal biology of reptiles as a whole scientific trend has formed, strengthened its scientific basis, more or less – the conceptual apparatus and methodological research approaches (unfortunately, only “more or less” – this aspect we will discuss later).
In the zero years of the XXI century there was a continuation of accumulation of the field data in thermal ecology of certain reptile species and their relationships with environmental conditions, study and comparison of thermal biological indicators, various aspects of thermal physiology, and most importantly – an increasing number of analytical publications appeared, sometimes concerning problems and models of thermoregulation, evolution of the thermal biological sphere (for this time, we know near 900 publications in the foreign scientific press, see figure).
However, these analytical articles did not always have an objectively high scientific value due to a number of standard methodological incorrectnesses, as well as insufficient development of the methodological base – standardization of the conceptual apparatus, etc. It seems that the main achievements corresponding to the level of development of science in this area were received back in the 1960–1990s, and now the deep understanding and development, the appearance of “breakthrough” ideas and developments were required, but this was not as good as we would like.
Since the beginning of the 10s. of XXI century the number of publications on reptiles’ thermal biology in the foreign press began to decline. The interest in this area gradually faded. Most likely, this can be explained by the fact that over all previous years a great deal of material has been accumulated on the thermal biology of certain reptile species, but no adequate general view, general conception of this trend of science, reaching the general biological level, has been proposed. Under these conditions, the further accumulation of data gradually lost its meaning and significance.
Thus, the 1980s, 1990s and zero years of the XXI century in foreign countries have become the golden age of reptiles’ thermal biology. The main databases of primary data on the thermal biology of reptiles and more or less generalizing ideas were accumulated precisely at this period.
Many foreign researchers who have studied ecology, physiology, ecological physiology, physiological ecology and other areas of reptiles’ thermal biology in different years have made a huge contribution to research on trend of science. These scientists cannot be ignored here: S.C. Anderson, R.A. Anderson, R.A. Avery, R.M. Andrews, F.H. Pough, R.B. Cowles, C.M. Bogert, P. Licht, G.A. Bartholomew, A.F. Bennett, S.D. Bradshaw, B.H. Brattstrom, T.J. Case, J.L.Cloudsley-Thompson, D. Crews, P.M.C. Davies, W.R. Dawson, B.T. Firth, E. Frankenberg, C. Gans, B. Green, O.E. Greenwald, P.T. Gregory, G.C. Grigg, L. Gvozdik, A. Hailey, J.J. Harlow, J.E. Heath, H. Heatwole, P.E. Hertz, R.B. Huey, V.H. Hutchison, H.B. John-Alder, W.H. Karasov, D. King, C.B. De Witt, J.C. Lee, H. Lillywhite, W.W. Mainne, S.M. McGinnis, R. Meek, K.A. Nagy, G. Naulleau, K.S. Norris, J.W. Patterson, E.R. Pianka, F.H. Pough, C.L. Prosser, P.J. Regal, H. Saint-Girons, K. Schmidt -Nielsen, F. Seebacher, R. Shine, V.H. Shoemaker, I.F. Spellerberg, R.D. Stevenson, J.R. Templeton, C.R. Tracy, H. Underwood, S. Waldschmidt, G.J.W.Webb, Y.L.Werner, J.M. Whittier, and many others.
The state of knowledge and problems in studies of thermal biology of reptiles in foreign countries
During more than 180 years of studying the thermal biology of reptiles, huge factual material has been accumulated from field and laboratory observations and special laboratory experiments. The ecology, geographical distribution, biotopical location, thermal biology, physiology, ecological physiology and physiological ecology of many species of reptiles were studied. A large number of articles and many books have been published on these topics. Much in the biology of reptiles has become more understandable and scientifically proved. It is simply impossible to describe all the achievements in this field. They are very diverse and multifaceted. But we regret to note that in the works of foreign researchers in reptiles’ thermal biology of the last time, despite the abundance of publications, there are some negative, in our opinion, aspects.
One of the serious problems in ecological, physiological, physiology-ecological and other studies is the widespread use of standard research methods that are familiar from the middle of the last century, which, as recent works have shown, contain certain methodological errors. Unfortunately, they distort the results and lead to erroneous biological conclusions (Черлин, 2010, 2013, 2014a).
The objectivity of the presence of these problems is partly proved by the materials of detailed ecological, naturalistic observations and studies, partly by the fact that it is precisely and only with a methodologically correct variant of work that the thermobiological indicators demonstrate quite definite regularities. Thus, the body temperatures range at full activity and the thermostabilization range, got by the methodically correctly performed work, appear to be stable within the species, the same in spring, summer and autumn, and even in biotopes with different climatic conditions (cool and hot if animals of this species inhabit them). In fact, during all seasons of the year, reptiles of the same species during time periods when they actually choose their body temperature try to keep it in the same range (review by Черлин, 2014a).
But most often there is another option in the work of herpetologists: the data are collected simply "in activity", i.e. when an animal is met by researcher in the activity zone and moves: for example, when diurnal lizard is meet in the daytime on the soil surface, the researcher does not allocate a state of full activity and does not take into account how the lizard behaves in relation to heat (tries to fasten the heating, chooses a specific range for high body temperature, do not select the body temperature, strive to avoid overheating or to maximize time spent at high body temperatures, etc.). In other words, in this variant the work is performed methodically incorrectly. In this case, the body temperatures range while such activity will be significantly wider than the body temperature range while full activity. Moreover, the preferred temperature (the average of this sample) will not bear any serious meaning, and will show variability: it will be higher in warmer climate and weather conditions (e.g. summer in the lowlands, in the south regions) and lower in the cooler (e.g., spring and autumn, in the mountains or in the North). In this case, the same characters obtained correctly or incorrectly (for example, the temperature of thermal stabilization and preferred body temperatures), can differ not in 1-2°, but in cool periods can reach up to 10° (!). So, for the same species, different researchers, using incorrect or correct methods for collecting primary data, can show the preferred body temperature respectively 22 or 32° (I do not want to refer to the concrete source, so as not to offend anyone)! And these are the fundamental, fatal differences!
Thus, there is a confusion in concepts, in research methods, in indicators of activity and thermal preferences (optimal, preferred, voluntary, average), in methods of obtaining primary data and data processing options, etc. These facts show objectively the need for standardization of the conceptual apparatus, accounting for activity statuses in reptiles’ thermal biology, correction of research methods, etc. And, despite the assertion of many famous researchers that “we don’t need to complicate everything, now it is just understandable”, the situation is quite obvious: the organization in order the conceptual apparatus, methodological and methodical parts of thermal biological research is absolutely necessary!
The part of the general problems in thermal biological studies in reptiles is also related to the fact that a unified, scientifically substantiated system of thermal biological terms and concepts has not yet been adopted. Unfortunately, a majority of herpetologists does not recognize this circumstance, believing that everything is just clear enough without paying special attention to it. But in fact, the different authors use the same terms to denote different thermobiological characters obtained by various methods. As a result, such identical terms carry different semantic loads, i.e., they simply reflect different biological phenomena, although researchers try to compare them and draw far-reaching general biological conclusions from this analysis. One does not need to be a genius to understand that in this case all the scientific constructions obtained in this way are far from the truth.
Despite the apparent obviousness of the situation, the vast majority of herpetologists involved in researches on reptiles’ thermal biology, especially in the field, do not distinguish statuses and forms of activity with all the incorrectnesses and errors resulting from this. Worse, if in any article there are given the tables that summarize a huge number of measurements of body temperatures of different reptile species, but the measurement method is not described, i.e. it is not indicated in what activity status these measurements were taken, or at least it is not simply described in details, all these data lose their scientific value, since it is impossible to understand what and how was received and processed, whether the methodical errors were made or not, and even what the data mean in sense. Nevertheless, the resulting erroneous conclusions are used by researchers in separate and general biological constructions, which as a result also naturally turn out to be incorrect.
Unfortunately, both foreign and domestic researchers suffer from this "disease", and our attempts to explain the situation are often perceived "without enthusiasm", with resentment, or even as a "brazen encroachment" on "indisputable authorities" and "unconditional and already established axioms".
In this regard, I think, that the most important laws related to the organization and regulation of the reptiles’ life are probably still undescribed and remains misunderstood by the scientific community.
The situation with the understanding of such a crucial aspect of the reptiles’ life as their thermal adaptation is also unfavorable. Many scientific publications have been devoted to them, and recently a book on this subject has even been published (Angilletta, 2009). However, in practice it turns out that they consider certain aspects of reptile biology (ecological physiology, physiology, ecology, general thermal biology of reptiles, etc.), but they have very little to do with the adaptations themselves. Here, in our opinion, it is important to understand that many regularities, dependencies and relationships of various phenomena in biochemistry, physiology, ecology and other fields of reptiles’ biology with temperature actually exist and are undeniable. The problem is not that they are gone or that we reject them. The problem is in that not all of these dependencies and relationships can be qualified as thermal adaptations (Черлин, 2015). Most likely, when investigating thermal adaptations, one would have to deal with, on the one hand, biochemical and physiological thermal compensations, and, on the other – two major aspects of the phenomenon of adaptation: 1) a complex of stable, immutable, species-specific characters of thermal homeostasis, and 2) a set of reactions that subtly "adjust" the spatial-temporal structure of the daily and seasonal activity in the direction of maintaining the invariance of the complex of characteristics of thermal homeostasis in different environmental conditions.
A number of papers have been performed and published on the principles of organization of thermoregulation mechanisms in reptiles, that sometimes was described by different mathematical models (Brattstrom, 1965; Heath, 1970; Berk, Heath, 1975; Barber, 1977; Barber, Crawford, 1977; Barber, 1979 ; Huey, 1982; Adolph, 1990; Adolph, Porter, 1993, etc.). These and other publications slightly lift the veil on the subject, describe some external manifestations of the thermoregulation mechanisms. But, despite their seriousness, unfortunately, they do not reveal the mechanisms of this phenomenon. Moreover, due to the lack of a common concept of thermal biology of reptiles, specialists have not yet developed an idea of what temperature reptiles regulate, for what, in what cases, etc. It is quite obvious that these animals regulate the body temperature not always. Indeed, the phenomenon of body temperature regulation actually turns out to be heterogeneous: sometimes reptiles maintain their body temperature in a certain range, limited above and below by dangerous (or unfavorable, "repulsive") boundaries; sometimes they maintain a certain temperature level (optimal, preferred, "eccritic", etc., or more precisely – the thermostabilization level or range (Черлин, 2014a; Cherlin, 2015b)), but this type of regulation is not always evident; they also maintain a certain body temperature during inactive states of (during periods of sleep, hibernation, and/or estivation). But it is still unclear: do animals maintain one certain temperature level in different physiological states or not? There is even no a single, generally accepted list of those thermobiological parameters that are affected by the mechanisms of regulation; it is also not clarified whether different thermal biological parameters are regulated by the same mechanisms or not... Reading many even very serious publications, it becomes obvious that scientists most often do not even ask to themselves these most important questions, and, therefore, do not try to answer on them. And without this, the whole system of relations of reptiles’ vital activity with a temperature factor will not be understood.
The problems with the evolution of the thermobiological sphere are also rather difficult. Such important and seemingly consistent with the modern understanding of the evolution process statements, such as, in particular, that the changes in the thermal environment entails a corresponding adaptive shift of the thermal biological norm of the reptiles’ population, in fact, are not obvious. And if it does exist, it requires special, experimental or other evidence. Our studies just show the controversy of such a claim (Черлин, 2015). It is quite obvious that in different geographical regions and climatic conditions there may be reptiles with very similar or just the same basic thermal physiological characters of activity, and in the same geographical regions and climatic conditions there can be reptiles with very different thermal biological characteristics. And this is due to the use of different spatial-temporal structures of daily and seasonal activity. In addition, we have clearly shown that the system of behavioral modifications of the spatial-temporal of activity structure is extremely effective in preventing any changes in the thermal physiological species norm. In fact, these behavioral responses are directed to preserve the characteristics of the thermal physiological species norm. In such a situation, it is difficult to assume the possibility of an adaptive change in the species thermal physiological norm, the immutability of which is so securely protected. But the change in this norm still happened. In our opinion, this may be a consequence, for example, of some nonadaptive purely biochemical, physiological, morphological, genetic, etc. changes in the nervous mechanism of temperature regulation, etc., which we do not yet know about. At the same time, the system of behavioral modifications, changing the time and place of activity of the animals, "adjusts", adapts a new physiological norm to the specific climatic conditions of the region, or causes the animals with this new norm to change the biotopical distribution and/or even geographical distribution (i.e. change the temporal and spatial placement). Then the natural selection helps to preserve successful combinations of a new species-specific norm, a complex of behavioral responses and external conditions. In this case, the new species norm is indeed well adapted to the climatic conditions of the region that the animal occupies, i.e. the adaptation process occurs primarily due to behavioral responses. But it can hardly be argued that a new species norm has emerged as a result of adaptation to these climatic conditions.
In addition, unfortunately, in researchers, in herpetologists, the healthy and very important for zoologists naturalism weakens significantly, i.e. they cease to be real naturalists. Sometimes the inordinate desire to bring the obtained primary data into different models leads to a discussion of the major aspects of animal life only in the terms of mathematical formulas. These models often become an end in themselves, and model comparisons turn into "games of mind" that have little contact with the real life of reptiles. I draw your attention to the fact that we just do not oppose mathematical data processing and the use of mathematical models. Simply mathematical processing should be a help, sometimes - very important, but still – а help for the scientist on the way to understanding how and why animals live this way, and not otherwise. But sometimes it is more important and valuable to notice some small element of animal behavior (which can only be noticed by a good naturalist), revealing a whole layer of his life, than, for example, to compare theoretical models of regulation of any physiological parameters, poorly imagining by what real laws the physiological system and ecology of these animals is organized and how these laws manifest themselves.
An example of the importance of subtle naturalistic observations is the description of poorly, implicitly manifested behavioral features that revealed the state of full activity in reptiles behavior and opened, after that, a whole layer of extremely important thermal physiological processes, phenomena, characteristics, and regularities (Либерман, Покровская, 1943; Cowles, Bogert, 1944; Cherlin, 2015b; Черлин, 2016). And as an example of a very successful application of mathematical modeling not detached from the object of study, we can mention the works of prof. A.V.Korosov (Коросов, 2008, 2010) on the modeling of thermoregulation of the common adder, Pelias berus.
***
Despite the fact that since the 1940s. several books and summaries have been published on the thermal biology of reptiles or related to it topics (Cowles, Bogert, 1944; Brattstrom, 1965; Tempelton, 1970; Cloudsley-Thompson, 1971; Comparative ..., 1973; Schmidt-Nielsen, 1964, 1997, etc.) and many articles, unfortunately, there is no generally accepted systemic understanding of this area of science. While researches, analysis of materials and conclusions are largely determined by long-established methods and concepts and follow the traditional, “knurled track” (using sometimes beautiful and modern technical means and methods of material processing, “automatically” providing for many researchers unconditional acceptance of the obtained data and publications by the scientific public). In order to get finally out of this situation, it is necessary to introduce into the research a “fresh stream”.
Thus, we can conclude that the abundance of publications on the thermal biology of reptiles does not provide itself an objective picture of this scientific direction. It is necessary to formulate, finally, a common concept in the thermal biology of reptiles, to standardize and scientifically substantiate a system of concepts, terms and indicators, methods of collecting and processing materials, etc., which the scientific "thermobiological community" is actively resisting, clearly mistakenly believing that all this is already there, or that it is possible to do without it.
It seems to us that in this case the problems we have described in this field of science better and more clearly characterize the state of study of the thermal biology of reptiles than the listing of separate achievements that are not connected and united by a single idea.
[1] Full activity – the state of activity, where animals are in a physiological state, fully, without significant physiological constraints, ensuring the efficient course of all major behavioral acts (locomotion, thermoregulation, nutrition, sexuality, territoriality, protection, communication, etc.). In this state, the movements of reptiles are active, rapid (adequate for species characteristics) and accurate (Либерман, Покровская, 1943; Dawson, 1975; Cowles, Bogert, 1944; Черлин, 2014b; Cherlin, 2015a).
[2] Temperature (range) of thermostabilization – is a level or rather narrow range of high body temperatures that reptiles, being in a full activity state, direct and promptly maintain for more or less long periods of time. Numerically, it almost corresponds to what was previously called in the literature "optimal" or "preferred" temperatures, defined in a methodologically correct way (which happened, alas, infrequently), but in fact, this range is most often not optimal for any physiological processes in the body, and not preferred, because basically it is a compromise between the desired and possible levels of temperatures. Simply reptiles for a number of reasons (not until yet fully understood) stabilize in it their body temperature. Therefore, we found it reasonable and appropriate to change its name to the “temperature (range) of thermostabilization” or “thermostabilization range”, removing from it an assessment of the reasons for this stabilization (Черлин, 2014a).
References
Bligh J., Johnson K. G. Glossary of terms for thermal physiology, J. Appl. Physiol. 1973. No 35. P. 941–961.
Cherlin V. A. Why the complex of thermobiological characters is so and not another?, Some important aspects of thermal biology of reptiles. Saarbrücken, Lambert Academic Publishing, 2015b. P. 55–116.
Adolph S. C. Influence of behavorial thermoregulation on microhabitat use by two Sceloporus lizards, Ecology. 1990. No 71. P. 315–327.
Adolph S. C., Porter W. P. Temperature, activity, and life histories, The American Naturalist. 1993. Vol. 142. No 2. P. 273–295.
Agassiz L., Gould A. A. Principles of zoology: Touching the structure, development, distribution and natural arrangement of the races of animals, living and extinct. Part I. Comparative physiology. Sheldon, Blakeman and Co., New York, 1856. 250 p.
Angilletta M. J. Thermal Adaptation: A Theoretical and Empirical Synthesis. Oxford: Oxford University Press, 2009. 289 r.
Atsatt S. R. Color changes as controlled by temperature and light in the lizards of the desert regions of southern California, Univ. Calif. Los Angeles Publ. Biol. Sci. 1939. No 1. P. 237–276.
Bailey R. M. Temperature toleration of garter snakes in hibernation, Ecology. 1949. Vol. 30. P. 238–242.
Bailey R. M. Winter mortality in the snake Storeria dekay, Copeia. 1948. No 3.
Baldwin F. M. Body temperature changes in turtles and their physiological interpretations (Chrysemys marginata belli, C. Gray and Chelydra serpentina, Lin.), Amer. Jour. Physiol. 1925a. No 72. P. 210–211.
Baldwin F. M. The relation of body to environmental temperatures in turtles Chrysemys marginata belli (Gray) and Chelydra serpentina (Lin.), Biol. Bull. 1925b. No 48. P. 432–445.
Barber B. J. A theoretical and experimental analysis of behavioral temperature regulation in the lizard, Dipsosaurus dorsalis, Dissertation Abst. Int. 37. 1977. R. 4330.
Barber B. J. Dual threshold control of peripheral temperature in the lizard Dipsosaurus dorsalis, Physiol. Zool. 1979. Vol. 52. P. 250–263.
Barber B. J., Crawford E. C. Jr. A stochastic dual-limit hypothesis for behavioral thermoregulation in lizards, Physiol. Zool. 1977. Vol. 50. No 1. P. 53–60.
Benedict F. G. The physiology of large reptiles with special reference to the heat production of snakes, tortoises, lizards and alligators. Washington: Carnegie Institution, 1932. No 425. P. 539.
Bennett A. F. A comparison of activities of metabolic enzymes in lizards and rats, Comp Biochem. Physiol. 1972. 42B. P. 637–647.
Bennett A. F., Hicks J. W., Cullum A. J. An experimental test of the thermoregulatory hypothesis for the evolution of endothermy, Evolution. 2000. Vol. 54. No 5. P. 1768–1773.
Bennett A. F., Ruben J. A. Endothermy and activity in vertebrates, Science. 1979. Vol. 206. P. 649–654.
Berk M. L., Heath J. E. An analysis of behavioral thermoregulation in the lizard, Dipsosaurus dorsalis, J. Therm. Biology. 1975. No 1. P. 15–22.
Blum H. F., Spealman C. R. Note on the killing of rattlesnakes by "sunlight", Copeia. 1933. P. 150–151.
Bogert C. M. Body temperatures of the tuatara under natural conditions, Zoologica. 1953. No 38. P. 63–64.
Bogert C. M. How reptiles regulate their body temperature, Scientific American. 1959. No 200. P. 105–120.
Bogert C. M. Reptiles under the sun, Nat. Hist. 1939. Vol. 24. P. 26–37.
Bogert C. M. Thermoregulation and eccritic body temperatures in Mexican lizards of the genus Sceloporus, Ann. Inst. Biol. Mexico. 1949b. No 20. P. 415–426.
Bogert C. M. Thermoregulation in reptiles, a factor in evolution, Evolution. 1949a. No 3. P. 195–211.
Brattstrom V. N. Body temperatures of reptiles, Amer. Midl. Naturalist. 1965. Vol. 73. No 2. P. 376–492.
Buxton P. A. Animal life in deserts: A study of the fauna in relation to environment. London: Edward Arnold and Co., 1923. 176 p.
Buxton P. A. Heat, moisture and animal life in deserts, Proc. Roy. 1924. Soc. B 96. P. 123–131.
Camp C. L. Notes on the local distribution and habits of the amphibians and reptiles of southeastern California in the vicinity of the Turtle Mountains, Univ. Calif. Publ. Zool. 1916. Vol. 12. P. 503–544.
Carpenter C. C. Body temperatures of three species of Thamnophis, Ecology. 1956. Vol. 37. P. 732–735.
Carpenter C. C. Comparative ecology of the common garter snake (Thamnophis s. sirtalis), the ribbon snake (Thamnophis s. sauritus), and Butler's garter snake (Thamnophis butleri) in mixed populations, Ecol. Monogr. 1952. Vol. 22. P. 235–258.
Cherlin V. A. About the article of S.S.Lieberman and N.V.Pokrovskaya on the thermal biology of sand lizard, Principy ekologii. 2014b. T. 3. No. 3. P. 25–32.
Cherlin V. A. Difficulties and possible errors in field investigations on the thermal biology of reptiles, Sovremennaya gerpetologiya: problemy i puti ih resheniya: Stat'i po materialam dokladov Pervoy mezhdunarodnoy molodezhnoy konferencii gerpetologov Rossii i sopredel'nyh stran (Sankt-Peterburg, Rossiya, 25–27 noyabrya 2013 g.), Zoologicheskiy institut RAN. SPb., 2013. P. 32–39.
Cherlin V. A. Heat adaptation of reptiles and their forming mechanisms, Principy ekologii. 2015. T. 4. No. 1. P. 17–76.
Cherlin V. A. Modern view on thermal biology from the position of studying reptiles, Biosfera. 2016. T. 8. No. 1. P. 47–67.
Cherlin V. A. Reptiles: temperature and ecology. Saarbrücken: Lambert Academic Publishing, 2014a. 442 p.
Cherlin V. A. The activity statuses and their significance in physiological ecology of reptiles and other ectothermic animals, Some important aspects of thermal biology of reptiles. Saarbrücken, Lambert Academic Publishing, 2015a. P. 11–54.
Cherlin V. A. Thermal biology of reptiles. General information and experimental technique (manual). SPb.: Russko-Baltiyskiy informacionnyy centr «BLIC», 2010. 124 p.
Cloudsley-Thompson J. L. The Temperature and Water Relations of Reptiles. Merrow Publishing Co., Ltd., Watford, Herts., England, 1971. 159 p.
Colbert E. H., Cowles R. B., Bogert C. M. Temperature tolerances in the American alligator and their bearing on the habits, evolution and extinction of the dinosaurs, Bull. Am. Mus. Nat. Hist. 1946. No 86. P. 329–373.
Comparative animal physiology. (L. Prosser, Ed.). Philadelphia, London, Toronto: W. B. Saunders Company, 1973. 888 p.
Cowles R. B. Additional implications of reptilian sensitivity to high temperature, Amer. Nat. 1940. Vol. 75. P. 542–561.
Cowles R. B. Critical thermal levels and thermal regulation in desert reptiles, 25th Ann. Meeting Amer. Soc. Ichthyologists and Herpetologists, Amer. Mus. Nat. Hist. Abstr. 1942. No 10 (mimeographed).
Cowles R. B. Heat-induced sterility and its possible bearing on evolution, Ibid. 1945. Vol. 79. P. 160–175.
Cowles R. B. Parturition in the yucca night-lizard, Xantusia vigilis, Copeia. 1944. No 1. P. 98–100.
Cowles R. B. Possible implications of reptilian thermal tolerance, Science. 1939. Vol. 90. P. 465–466.
Cowles R. B. Possible origin of dermal temperature regulation, Evolution. 1958a. No 12. P. 347–357.
Cowles R. B. Semantics in biothermal studies, Science. 1962. No 135. P. 670.
Cowles R. B. Sidewinding locomotion in snakes, Copeia. 1956. No 4. P. 211–214.
Cowles R. B. The effect of temperature on thyroid histology and survival in the lizard, Sceloporus occidentalis, Copeia. 1958b. P. 265–276.
Cowles R. B. Winter activities of desert reptiles, Ecology. 1941. Vol. 22. P. 125–140.
Cowles R. B., Bogert C. M. Preliminary study of the thermal requirements of desert reptiles, Bulletin of the American Museum of Natural History. 1944. No 83. P. 261–296.
Cowles R. B., Burleson G. L. The sterilizing effect of high temperature on the male germ-plasm of the yucca night lizard, Xantusia vigilis, Am. Nat. 1945. Vol. 79. P. 417–435.
Ctrel'nikov I. D. Light as a factor in the ecology of animals. The effect of solar radiation on the body temperature of some poikilothermic animals., Izvestiya Nauchnogo instituta imeni P. F. Lesgafta. 1934. T. 17–18. P. 313–372.
Dawson W. R. On the physiological significance of the preferred body temperatures of reptiles, Perspectives of biophysical ecology. Ecological studies. Vol. 12. (D. M. Gates and R. B. Schmerl, Eds.). New York, 1975. P. 443–473
Evans E. E., Cowles R. B. Effect of temperature on antibody synthesis in the reptile Dipsosaurus dorsalis, Proc. Soc. Exp. Biol. Med. 1959. No 101. P. 482–483.
Fitch H. S. Life history and ecology of the five-lined skink, Eumeces fasciatus, Univ. Kansas. Publ. Mus. Nat. Hist. 1954. No 8. P. 1–156.
Fitch H. S. Temperature responses in free-living amphibians and reptiles of northeastern Kansas, Univ. Kansas Publ. Mus. Nat. Hist. 1956. No 8. P. 417–476.
Fox W. Effect of temperature on development of scutellation in gater snake, Thamnophis elegans atratus, Copeia. 1948. No 4. P. 252–262.
Franz N. Untersuchungen über den Wärmebaushalt der Poikilothermen, Biolog. Zentralblatt. 1930. Bd. 50. S. 158–181.
Gadow H. Amphibia and reptiles, The Cambridge Natural History (C. S. F. Harmer and A. E. Shipley, Eds.). London: Mc-Millan and Co, 1901. Vol. 8. P. 1–668.
Grinnell J. The biota of the San Bernardino Mountains, Univ. Calif. Publ. Zool. 1908. No 5. P. 1–170.
Hall F. G. The vital limit of exsiccation of certain animals, Biol. Bull. 1922. Vol. 42. P. 31–51.
Hardy J. D. The "set-point" concept in physiological temperature regulation, Physiological controls and regulations. Philadelphia, 1965. P. 98–116.
Heath J. E. Behavioral regulation of body temperature in poikilotherms, The Physiologist. 1970. Vol. 13. P. 339–410.
Herter K. Die Vorzungstemperaturen bel Landtieren, Naturwissenschaften. Berlin, 1941. Vol. 29. P. 155–164.
Hesse R. Tiergeographie auf oekologischer grundlage. Jena: Gustav Fischer, 1924. 613 p.
Hesse R., Allee W. C., Schmidt K. P. Ecological animal geography. New York: J. Wiley and Sons, 1937. 597 p.
Huey R. Temperature, physiology, and the ecology of reptiles, Biology of the Reptilia. Physiology (C). Vol. 12. (C. Gans and F. H. Pough, Eds.). London: Academic Press, 1982. P. 25–91.
Inger R. F. Temperature responses and ecological relations of two Bornean lizards, Ecology. 1959. Vol. 40. No 1. P. 127–136.
Kammerer P. Der Artenwandel auf Inseln. Vienna; Leipzig: Franz Deuticke, 1926. 324 p.
Kanitz A. Temperatur und Lebens-Vorgange im Allgemeinen, Tabulae Biol. 1925. Vol. 2. P. 9–25.
Klauber L. M. Four papers on the application of the statistical methods to herpetolo-gical problems. III. The correlation between scalation and life zones in San Diego County snakes, Bull. Zool. Soc. San Diego. 1941. Vol. 17. P. 73–79.
Klauber L. M. Studies of reptile life in the and southwest, Bull. Zool. Soc. San Diego. 1939. Vol. 14. P. 1–100.
Korosov A. V. Ecology of common adder in the North. Petrozavodsk: Izd-vo PetrGU, 2010. 264 p.
Korosov A. V. Simple model of basking of common adder, Sovremennaya gerpetologiya. 2008. T. 8. Vyp. 2. P. 118–136.
Krüger P. Uber die Bedeutung der ultraroten Strahlen für den Wärmehaushalt der Poikilothermen, Biol. Zbl. 1929. Vol. 49. P. 65–82.
Krüger P., Kern H. , Arch. Physiol. 1924. No 202. S. 119–138.
Lamarre-Picquot P. Troisieme memoirs sur l’incubation et qutre phenomenes observes ches les ophidiens, Comp. Rend., Acad. Sci. Paris, 1842. Vol. 14. P. 164.
Liberman S. S. Pokrovskaya I. V. Materials on the ecology of sand lizard, Zoologicheskiy zhurnal. 1943. T. 22. No. 2. P. 247–256.
Lowe C. H., Vance V. J. Acclimation of the critical thermal maximum of the reptile Urosaurus ornatus, Science. 1955. No 122. P. 73–75.
Lowe C. Notes on the mating of desert rattlesnakes, Copeia. 1942. P. 261–262.
Lueth F. X. Effects of temperature on snakes, Copeia. 1941. P. 125–132.
Lufti M. Das termotaktische Verhalten einiger Reptilien: Inaugural-Diss. Munchen, 1936.
McGinnis S. M., Moore R. G. Thermoregulation in the Boa constrictor, Herpetologica. 1969. No 25. P. 38.
Mcilhenny E. A. The alligator's life history. Boston: The Cristopher Publishing House, 1935. 117 p.
Mosauer W. A note on the sidewinding locomotion of snakes, Amer. Nat. 1930. Vol. 64. P. 178-183.
Mosauer W. The toleration of solar heat in desert reptiles, Ecology. 1936. Vol. 17. P. 56–66.
Mosauer W., Lazier E. L. Death from insolation in desert snakes, Copeia. 1933. P. 149.
Musacchia X. J. The viability of Chrysemys picta submerged at various temperatures, Physiological Zoology. 1959. No 32. P. 47–50.
Nelson D. O., Heath J. E., Prosser C. L. Evolution of temperature regulatory mechanisms, Amer. Zool. 1984. No 24. P. 791–807.
Norris K. S. The ecology of the desert iguana Dipsosaurus dorsalis, Ecology. 1953. No 34. P. 256–287.
Parker G. H. The time of submergence necessary to drown alligators and turtles, Occas. Papers Boston Soc. Nat. Hist. 1925. Vol. 5. P. 157–159.
Parker H. W. A new melanic lizard from Transjordania, and some speculations concerning melanism, Proc. Zool. Soc. London, 1935. P. 137–142.
Parker H. W. The lizard genus Aprasia; its taxonomic and temperature correlated variation, Bulletin of the British Museum of Natural History. 1956. No 3. P. 363–385
Pearse A. S. Animal ecology. New York: McGraw-Hill, 1931. 642 p.
Pearse A. S., Hall F. G. Homoiothermism: The origins of warm-blooded vertebrates. New York: J. Wiley and Sons, 1928. 119 p.
Pearson O. P. Habits of the lizard, Liolaemus multiformis multiformis at high altitudes in southern Peru, Copeia. 1954. No 2. P. 111–116.
Pearson O. P., Brandford D. F. Thermoregulation of lizards and toad at high altitudes in Peru, Copeia. 1976. No 1. P. 155–170.
Reese A. M. Some reactions of Alligator mississippiensis, Jour. Comp. Psychol. 1923. Vol. 3. No 1. P. 51–59.
Reichling H. Transpiration und Vorzugstemperatur mitteleuropiiischer Reptilien und Amphibien, Zool. Jb. (Syst.). 1957. Vol. 67. P. 1–64.
Richet Ch. La chaleur animale. (F. Alcan, Ed.). Paris, 1889. 307 p.
Saint-Girons H., Saint-Girons M. C. Cycle d'activite et thermoregulation chez les reptiles (lezards et serpents), Vie Milieu. 1956. No 7. P. 133–226.
Schmidt-Nielsen K. Animal physiology. New York: Cambridge University Press, 1997. 607 p.
Schmidt-Nielsen K. Desert animals: Physiological problems of heat and water. Oxford: Clarendon Press, 1964. 277 p.
Soetbeer F. Ueber die Korperwarme der poikilothermen Wirbelthiere, Arch. Exp. Pathol. Pharmakol. 1898. Vol. 40. P. 53–80.
Sutherlanad A. The temperature of reptiles, monotremes and marsupials, Proceedings of the Royal Society of Victoria. 1896. No 9. P. 57–67.
Templeton J. R. Reptiles, The Comparative Physiology of Thermoregulation. (G. C. Whittow, Ed.). New York; London: Academic Press, 1970. Vol. 1. P. 167–221.
Valensiennes A. Obsrevations faites pendant l’incubation d’une femelle du python a deux raies (Pythin bivittatus, Kuhl.) pendant les mois de mai et de juin 1841, Comp. Rend., Acad. Sci. Paris, 1841. Vol. 13. P. 126–133.
Warden C. J., Jenkins T. N., Warner L. H. Comparative psychology. Vertebrates. New York: The Ronald Press, 1936. 560 p.
Weese A. O. An experimental study of the reactions of the horned lizard, Phrynosoma modestum Gir., a reptile of the semi-arid desert, Biol. Bull. 1917. No 32. P. 98–116.
Weese A. O. Environmental reactions of Phrynosoma, Amer. Naturalist. 1919. No 53. P. 33–54.
Weigmann R. Die wirkung starker abkuhlung auf amphibien und reptilian, Zeitschrift fiir Wissenschaffliche Zoologie. 1929. P. 1–692.
Werner F. Beiträge zur Kenntnis der Reptilien und Amphibien von Istrien und Dalmatien, Verh. zool, bot. Ges. Wien. 1891. No 41. S. 751–768.
Woodbury A. M., Hardy R. Studies of the desert tortoise Gopherus agassizii, Ecol. Monogr. 1948. Vol. 18. P. 145–200.

 © 2011 - 2026
© 2011 - 2026