



Введение
Жуки семейства Carabidae широко распространены по всему миру. Они хорошо изучены в биологическом и экологическом отношениях, считаются прекрасными индикаторами среды обитания (Кисилев, 2005; Lovei, 2008; Koivula, 2011; Dangale, 2012). Публикаций на тему изменчивости размеров карабид значительно меньше. Большинство таких работ выполнены на межвидовом уровне, когда оценивается не межпопуляционная изменчивость морфометрических признаков, а доля видов, различающихся по размерам, в сообществе (Sustek, 1987; Jelaska et el., 2010 и др.). Несмотря на обширный список работ, посвященных изучению закономерностей изменения размеров тела у насекомых, очень в немногих раскрыты непосредственные причины, определяющие изменчивость этого признака. Большинство обзорных публикаций является компиляцией работ, выполненных в разное время, разными авторами и по разной методике (Chown, Gaston, 2009; Stillwell et al., 2010; Gouws et al., 2011). В таких работах, как правило, не представляются данные по условиям среды, при которых собирали объекты. В то же время известно, что изменение отдельных факторов среды с неизбежностью влечет за собой изменение тех органов и признаков, которые по своей функции связаны с данными факторами среды (Шварц, 1980). Реакции организма, возникающие в связи с воздействием факторов внешней среды, являются первоначально своего рода раздражителями, вызывающими, в свою очередь, формообразовательную реакцию. А. Н. Северцов также отмечал, что биологически важные изменения строения и функций живых организмов происходят под прямым или косвенным влиянием внешней среды и в соответствии с изменениями этой среды. Биологически важная перемена в условиях существования (среде) данного вида животных является стимулом к изменению его организации; характер же изменения среды и количественное и качественное соотношение между изменением среды и функциями изменяющегося организма определяют направление, в котором пойдет эволюция изменяющегося вида в данную эпоху (Северцов, 1934). При этом важно не только гораздо более полно изучить факторы, связывающие индивидуальное формообразование организма (идущие под знаком прогрессивной дифференциации) в один целостный процесс. Нужно изучать факторы, определяющие координацию частей при филогенетических преобразованиях организма в одно единое целое (Шмальгаузен, 1968).
Проблема целостности организма, имеющая длительную историю, не только не утратила актаульности для современной биологии, но, наоборот, приобрела важнейшее гносеологическое значение с возникновением новых биологических дисциплин. Так, появилась макроэкология – поддисциплина экологии, которая имеет дело с изучением взаимоотношений организмов и окружающей среды в большом пространственном аспекте для объяснения статистических паттернов обилия распределения и разнообразия (Brown, Maurer, 1989). Эта область знаний традиционно была сосредоточена на межвидовом сравнительном анализе, но недавние работы начали анализировать макроэкологические паттерны этого явления с позиций географической перспективы (Gaston, Blackburn, 2000). Такой подход более ярко выявляет связи между экологическими признаками и средой обитания и обеспечивает понимание ряда широкомасштабных экологических явлений, в то время как большинство экологических исследований узконаправленные или экспериментальные, выражаются в детализированной информации, которая не может быть использована для распространения от одного сообщества к другому (Brown, 1995).
Изменчивость размеров тела жужелиц изучали на примере Poecilus cupreus. Проанализирован материал, собранный в разных точках ареала жужелицы Poecilus cupreus. Особенности статистической обработки позволили выделить факторы, влияющие на изменчивость размеров у этого вида. В наших работах по другим видам жужелиц было показано, что при воздействии одного и того же фактора среды разные отделы насекомого могут меняться разнонаправленно, и, наоборот, одинаковый характер изменчивости размеров конкретных органов насекомого может определяться разными факторами среды (Суходольская, Савельев, 2012; Sukhodolskaya, 2014). Цель настоящего исследования – определить вклад факторов среды в изменчивость размеров жужелицы Poecilus cupreus L., 1758.
Материалы
Объектом исследования была жужелица P. cupreus – широко распространенный палеаркт. Это лугово-полевой вид с высокой численностью в агроценозах. Имеет большое значение как энтомофаг. Находит благоприятные условия для обитания в городской черте и имеет широкий диапазон требований к среде обитания (Крыжановский, 1983).
В пределах Республики Татарстан жуков отлавливали стандартным методом почвенных ловушек. Ловушки экпонировались дважды в каждый из вегетационных сезонов 2006–2008 гг. Особи из других регионов в виде заспиртованных или высушенных на ватниках экземпляров были любезно предоставлены нам в рамках договоров о научном сотрудничестве Кемеровским государственным университетом, Институтом систематики и экологии животных СО РАН и Ставропольским государственным аграрным университетом. При этом авторы просили исследователей представить такой материал для морфометрических промеров, который наиболее полно подходил бы к схеме нашего исследования, то есть были бы задействованы города, агроценозы и естественные ценозы (табл. 1).
Таблица 1. Описание мест отлова жуков, задействованных в морфометрических промерах
Table 1. Location of sampling sites of beetles for morphometric measurements
| Регион | Географические координаты | Место отлова | Самки | Самцы | |
| Республика Татарстан | 55ос.ш. 49ов.д. | г. Казань | газон, сосняк | 63 | 63 |
| агроценозы | сельхозкультуры | 427 | 440 | ||
| естественные ценозы | сосняки, луга | 164 | 135 | ||
| Кемеровская область | 54ос.ш. 87ов.д. | г. Кемерово | сосняки, газоны, луга | 109 | 141 |
| агроценозы | сельхозкультуры | 145 | 111 | ||
| естественные ценозы | сосняк, луг | 39 | 47 | ||
| Новосибирская область | 55ос.ш. 27ов.д. | г. Новосибирск | сосняк, газон | 23 | 56 |
| естественные ценозы | сосняк, луга | 54 | 32 | ||
| Ставропольский край | 45ос.ш. 41ов.д. | г. Ставрополь | газоны | 26 | 18 |
| агроценозы | сельхозкультуры | 112 | 169 | ||
| Всего | 1286 | 1253 |
Жуков измеряли под бинокулярным микроскопом МБС-9 с мерной линейкой (цена деления 0.1 мм). Обмеры проводились индивидуально при дифференциации по полу по шести мерным признакам: длина надкрылий – расстояние по шву от середины бортика до вершины надкрылий (рис. 1, точки 1, 2); ширина надкрылий – расстояние между плечевыми углами надкрылий (рис. 1, точки 3, 4); длина переднеспинки – расстояние по средней линии от основания до вершины (рис. 1, точки 5, 6); ширина переднеспинки – ширина основания (рис. 1, точки 7, 8); длина головы – расстояние от шеи до верхней губы (рис. 1, точки 9, 10); расстояние между глазами (рис. 1, точки 11, 12). Всего промерено 2539 особей.

Рис. 1. Иллюстрация промеров жуков: 1–2 – длина надкрылий, 3–4 – ширина надкрылий, 5–6 – длина переднеспинки, 7–8 – ширина переднеспинки, 9–10 – длина головы, 11–12 – расстояние между глазами (ширина головы)
Fig. 1. Illustration of beetles measurements: 1–2 – elytra length, 3–4 – elytra width, 5–6 – pronotum length, 7–8 – pronotum width, 9–10 – head length, 11–12 – distance between eyes (head width)
Методы
Статистическую обработку результатов проводили в программе R (R development…, 2011) с использованием линейных моделей. В качестве независимых переменных рассматриваются регион и антропогенная нагрузка (обитание в городе, агроценозе, естественном биотопе). Влияние всех остальных переменных (факторов) считается случайным и суммарно оценивается в виде члена ошибки в модели. Используемые переменные рассматриваются как категориальные, в качестве базы сравнения для них используются: для региона – Республика Татарстан (как центр ареала P. cupreus), для антропогенной нагрузки – город. В иллюстрациях вклад факторов определенного типа кодирован соответствующими символами: @ – вклад региона, % – антропогенный вклад.
Вклады региона и антропогенной нагрузки в морфометрическую изменчивость популяций исследуемого вида считаются аддитивными и независимыми. Влияние перечисленных факторов считается различным для пола жука, кроме того, учитывается и собственно влияние пола. Другими словами, в модели участвует пол жука и парные взаимодействия пола с каждым из перечисленных факторов. Контрасты для кодов в моделях выбраны так, что базовыми (соответствующими нулевым значениям в матрице модели) для каждого из них являются перечисленные выше параметры. Для оценки значимости факторов использовался дисперсионный анализ построенной модели. Для каждого признака оценивался вклад всех переменных и взаимодействий с указанием доверительных интервалов и значимости (по критерию Стьюдента) и статистики остатков (ошибок). Полученные оценки и их доверительные интервалы использовались для представления результатов в графическом и табличном видах: взаимодействия сравнивались с базой сравнения (использовался 95 % доверительный интервал для нормальной аппроксимации). Кроме того, выводились доверительные интервалы для суммы вклада пола и отдельных переменных.
Использовали также метод главных компонент (PCA). Общепринятым считается, что первая главная компонента отражает изменение «масштаба» жужелицы (синхронное изменение признаков с учетом аллометрии). Если PC1 объясняет около половины дисперсии, то все методы анализа, основанные на расстояниях, будут отражать в основном именно эту «масштабную» изменчивость размеров.
Результаты
Размеры жуков отличаются высокой изменчивостью, что можно видеть по величине стандартного отклонения (табл. 2) и размаху доверительных интервалов (рис. 2, 3).
Таблица 2. Описательная статистика морфометрических промеров P. cupreus (мм)
Table 2. Descriptive statistics of morphometric measurements in P. cupreus (mm)
| Среднее | Минимум | Максимум | Станд.отклонение | Ошибка средней | ||||||
| самки | самцы | самки | самцы | самки | самцы | самки | самцы | самки | самцы | |
| Длина надкрылий | 6.88 | 6.73 | 4.20 | 4.90 | 8.50 | 8.50 | 0.49 | 0.49 | 0.01 | 0.01 |
| Ширина надкрылий | 3.05 | 3.01 | 1.60 | 1.40 | 5.70 | 6.30 | 0.72 | 0.90 | 0.02 | 0.03 |
| Длина переднеспинки | 2.41 | 2.35 | 1.50 | 1.20 | 3.50 | 3.40 | 0.24 | 0.21 | 0.01 | 0.01 |
| Ширина переднеспинки | 3.11 | 3.05 | 1.90 | 1.90 | 4.00 | 4.00 | 0.30 | 0.28 | 0.01 | 0.01 |
| Длина головы | 1.38 | 1.34 | 0.80 | 0.70 | 2.60 | 2.50 | 0.22 | 0.23 | 0.01 | 0.01 |
| Расстояние между глазами | 1.42 | 1.41 | 0.90 | 0.80 | 2.40 | 3.00 | 0.18 | 0.20 | 0.01 | 0.01 |
На рис. 2 и 3 представлены величины сдвигов в размерах надкрылий жуков. Это основной отдел тела жужелиц, по которому судят об общих размерах. Видно, что характер изменчивости длины и ширины надкрылий различается при действии одного и того же фактора среды. Условия обитания в Ставропольской области уменьшают длину надкрылий у самок, но увеличивают – у самцов. Ширина надкрылий в этих условиях увеличивается у особей обоих полов. Условия обитания в естественных ценозах никак не сказываются на ширине надкрылий жуков обоих полов, но увеличивают длину надкрылий у самок.
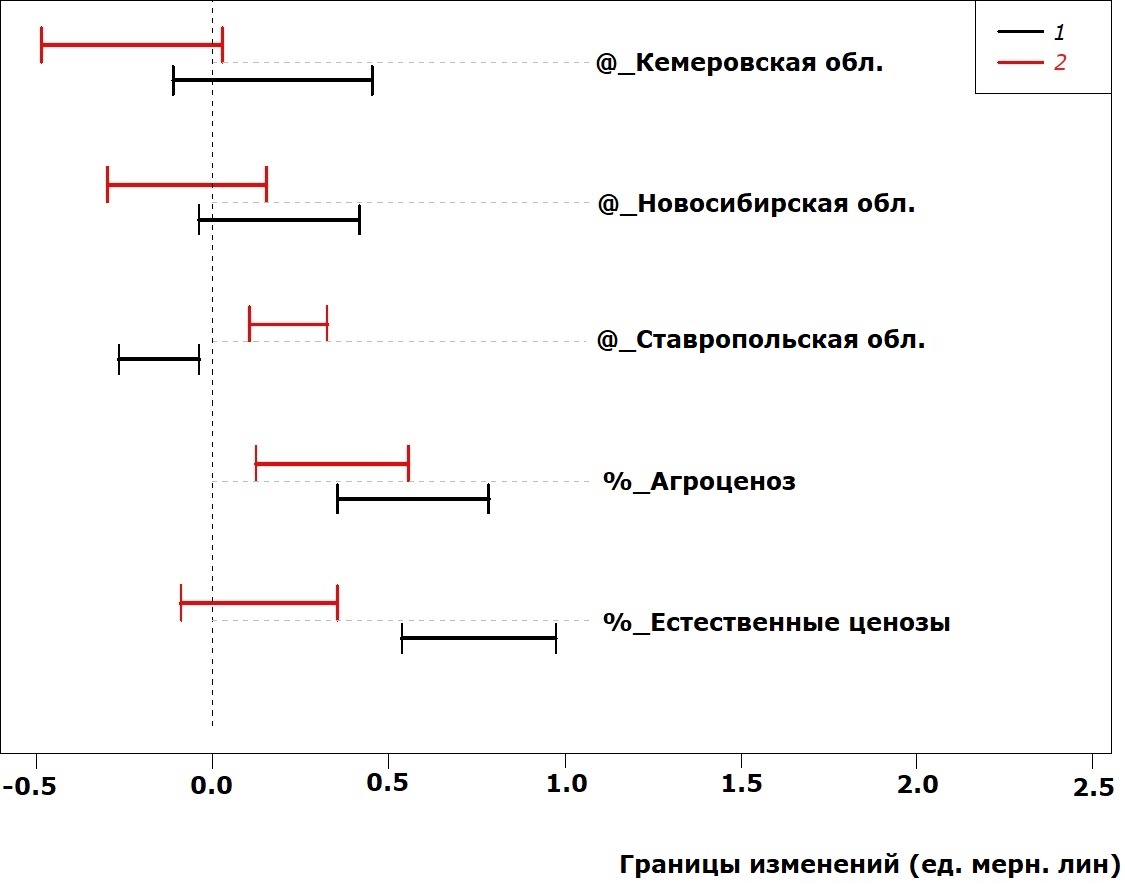
Рис. 2. Сдвиг в значениях длины надкрылий у P. cupreus за счет действия факторов среды (здесь и далее 1 – самки, 2 – самцы)
Fig. 2. Elytra length deviation in P. cupreus due to the impact of environmental factors (here and elsewhere 1 – females, 2 – males)
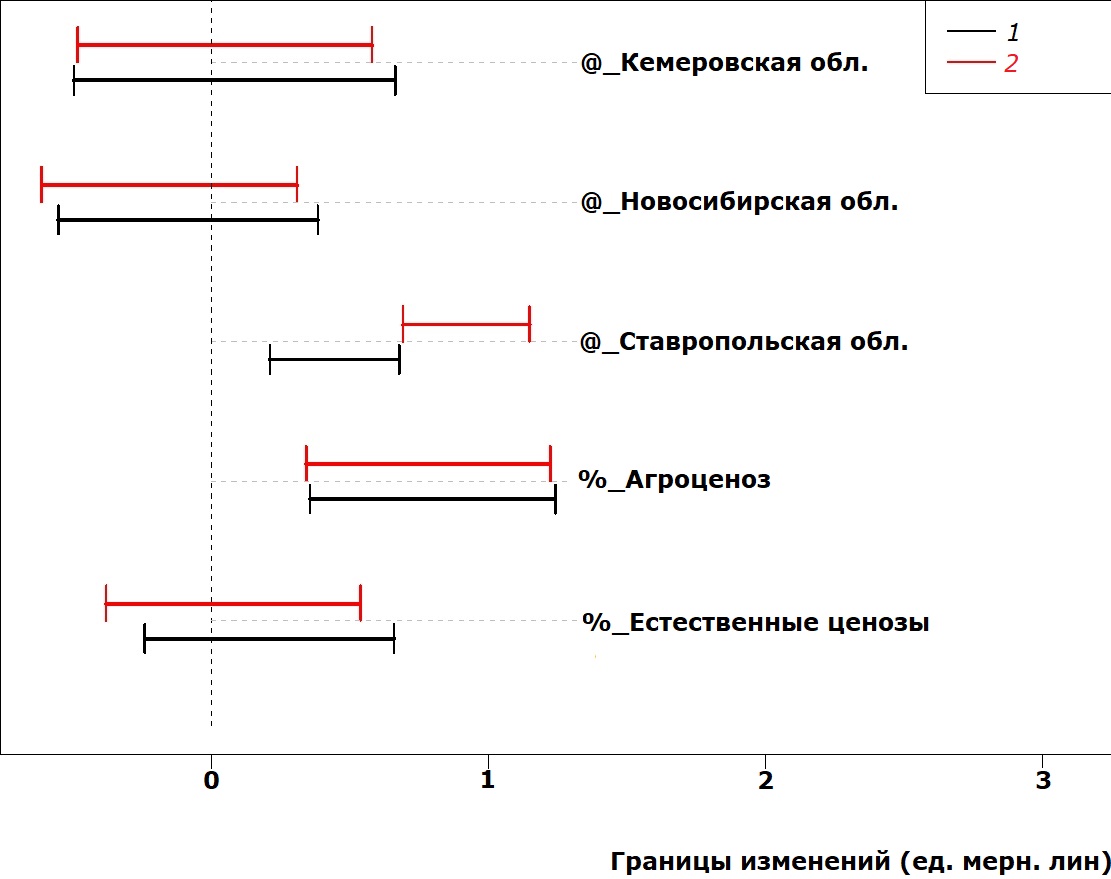
Рис. 3. Сдвиг в значениях ширины надкрылий у P. cupreus за счет действия факторов среды
Fig. 3. Elytra width deviation in P. cupreus due to the impact of environmental factors
Параметры переднеспинки под влиянием факторов среды меняются несколько иначе (рис. 4–5). Так, значимый вклад в длину переднеспинки помимо условий Ставрополя вносят условия обитания в Кемеровской области (в сторону увеличения), а ширина переднеспинки становится значимо меньше при обитании во всех исследованных регионах по сравнению с Республикой Татарстан (которая расположена севернее). Антропогенное влияние на параметры переднеспинки также разнонаправленно: если в условиях агроценоза и естественных ценозов длина переднеспинки увеличивается, то в этих же условиях ширина переднеспинки уменьшается. Причем этот феномен иногда не в равной степени выражен у самок и самцов.
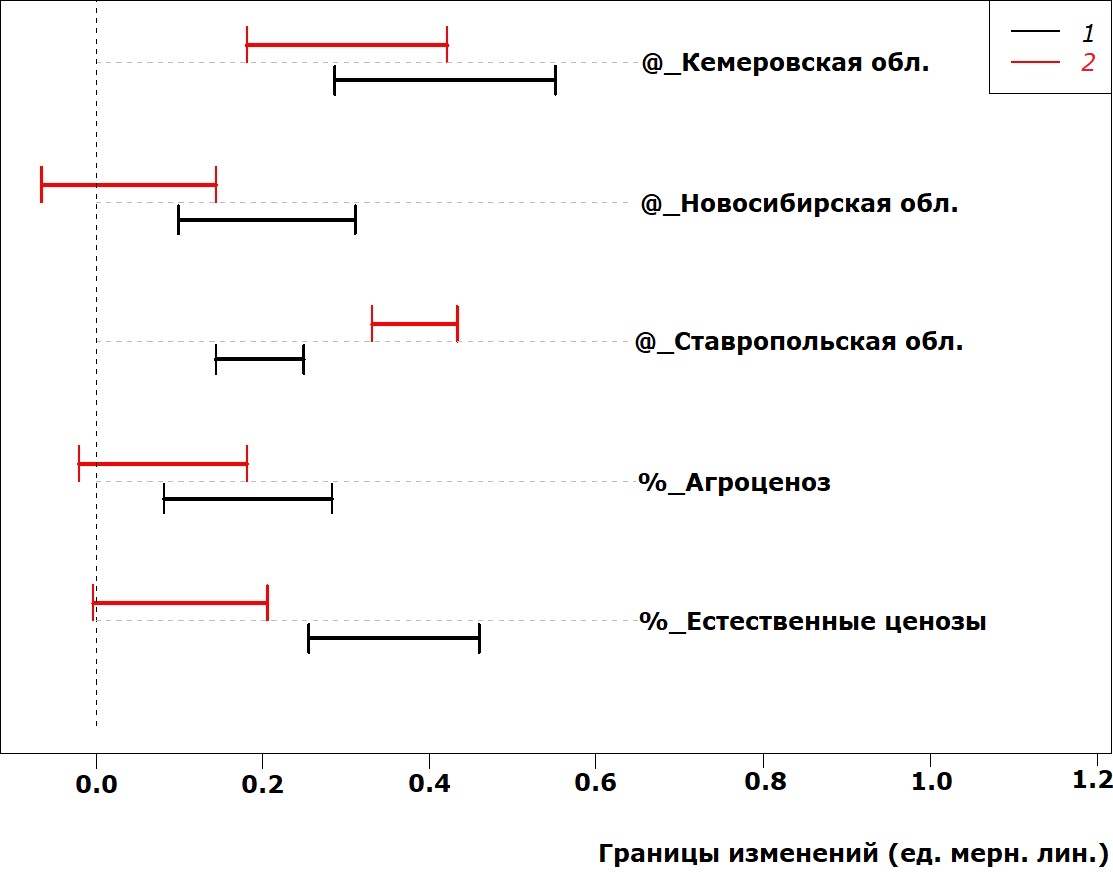
Рис. 4. Сдвиг в значениях длины переднеспинки у P. cupreus за счет действия факторов среды
Fig. 4. Pronotum length deviation in P. cupreus due to the impact of environmental factors
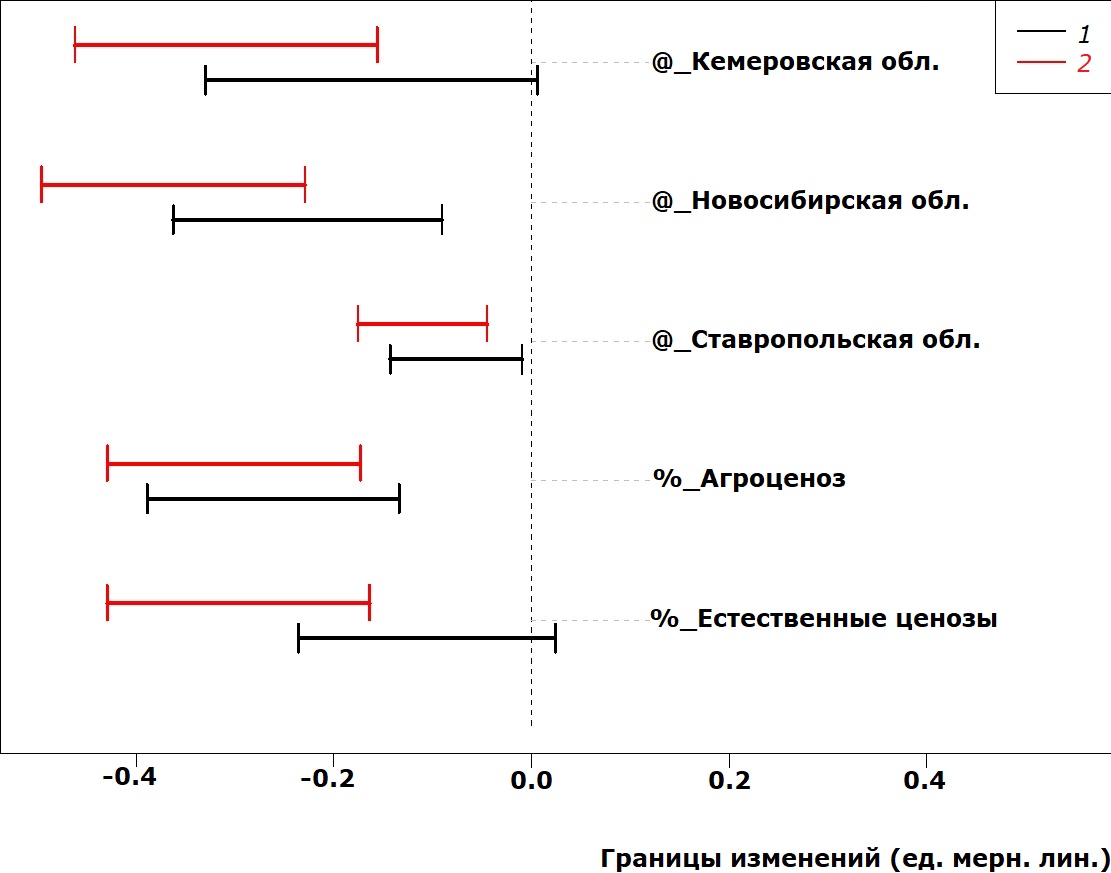
Рис. 5. Сдвиг в значениях ширины переднеспинки у P. cupreus за счет действия факторов среды
Fig. 5. Pronotum length deviation in P. cupreus due to the impact of environmental factors
Изменчивость параметров головы исследованных жуков представлена на рис. 6 и 7: под влиянием всех исследованных факторов значения длины и ширины головы увеличиваются (за исключением самцов в естественных ценозах).
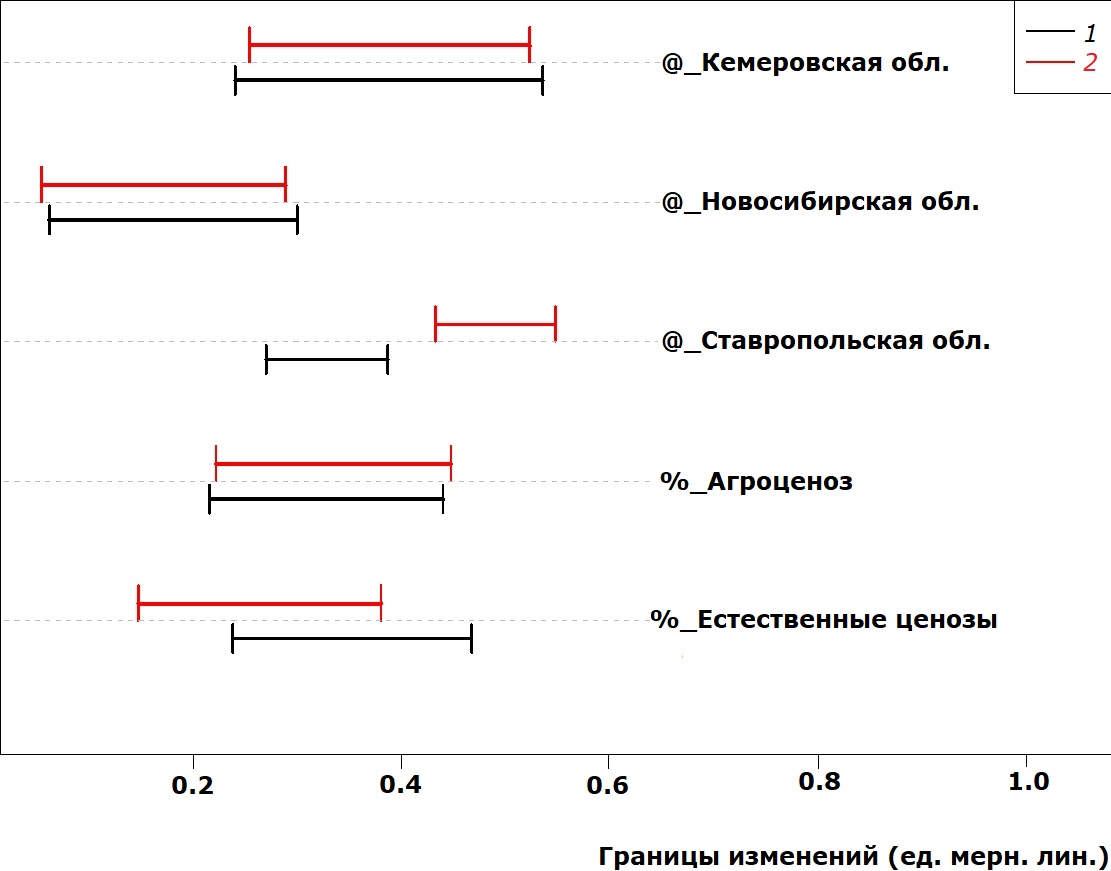
Рис. 6. Сдвиг в значениях длины головы у P. cupreus за счет действия факторов среды (на данном рисунке вертикальная штриховая линия, обозначающая базовые значения, совпадает с левой границей рисунка).
Fig. 6. Head length deviation in P. cupreus due to the impact of environmental factors (vertical dashed line denoting basic values coincides with the left border of the figure)
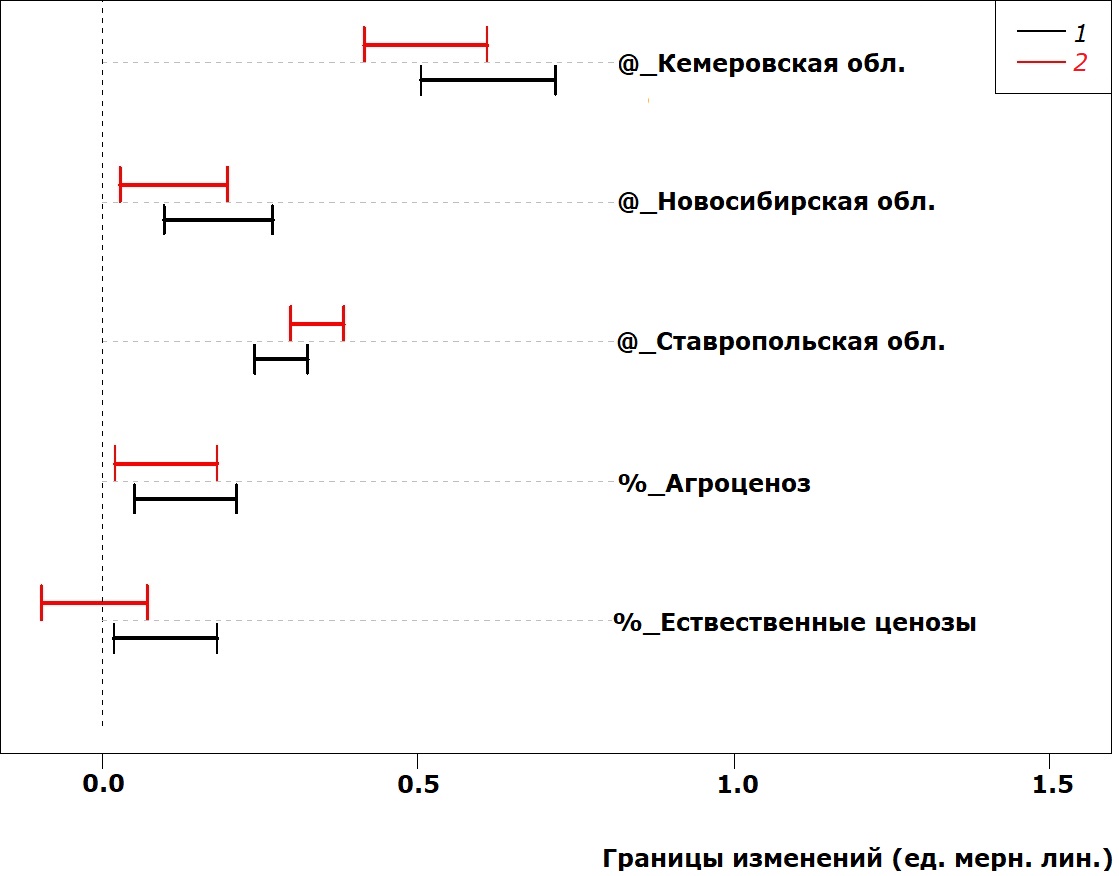
Рис. 7. Сдвиг в значениях ширины головы (расстояние между глазами) у P. cupreus за счет действия факторов среды
Fig. 7. Head width (distance between eyes) deviation in P. cupreus due to the impact of environmental factors
Обобщенный анализ результатов по изменчивости всех признаков свидетельствует о том, что в широтном аспекте изменчивость разных отделов тела P. cupreus различна: размеры надкрылий не показывают направленной изменчивости, длина переднеспинки больше в южных регионах по сравнению с северными, а ширина переднеспинки, наборот, в южных регионах меньше. Размеры головы уменьшаются по направлению к северу.
Результаты дисперсионного анализа модели, оценивающей вклад факторов в изменчивость, к примеру, длины надкрылий, представлены в табл. 3.
Таблица 3. Вклад экологических факторов в изменчивость длины надкрылий у P. cupreus
(F = 32.93, dF = 1286, R2 = 0.109, p < 0,001)
| Число степеней свободы | Сумма квадратов | Средние квадраты | Значение F | Уровень значимости | |
| Пол | 1 | 12.50 | 12.50 | 43.67 | *** |
| Пол:Регион | 6 | 65.42 | 10.90 | 37.21 | *** |
| Пол:Антропоген | 4 | 28.24 | 7.06 | 24.09 | *** |
| Остаточная | 2949 | 864.24 | 0.29 |
*** – уровень значимости p < 0,001.
Проанализировав материал с применением метода главных компонент (PCA), мы пришли к выводу, что разные отделы тела у жужелицы P. cupreus несут различающиеся факторные нагрузки (табл. 4, 5). Анализ этих таблиц показывает, что как у самок, так и самцов исследуемого вида регистрируется аллометрия, наиболее выраженная для ширины надкрылий. Это позволяет предположить существование изменчивости формы жуков в различной экологической обстановке.
Таблица 4. Факторные нагрузки признаков P. cupreus при анализе методом главных компонент (самки)
Table 4. Factors loading in P. cupreus traits in PCA (females)
| Признаки | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 |
| Длина надкрылий | 0.44 | 0.09 | –0.46 | 0.45 | 0.61 | 0.12 |
| Ширина надкрылий | 0.11 | 0.96 | –0.08 | –0.2 | –0.12 | –0.09 |
| Длина переднеспинки | 0.49 | –0.02 | 0.17 | –0.02 | –0.36 | 0.78 |
| Ширина переднеспинки | 0.41 | –0.24 | –0.61 | –0.25 | –0.47 | –0.35 |
| Длина головы | 0.43 | 0.03 | 0.51 | 0.52 | –0.22 | –0.47 |
| Расстояние между глазами | 0.44 | –0.11 | 0.35 | –0.65 | 0.47 | –0.16 |
| Доля объясненной дисперсии (%) | 47 | 16.9 | 13.1 | 9.2 | 7.1 | 6.7 |
Таблица 5. Факторные нагрузки признаков P. cupreus при анализе методом главных компонент (самцы)
Table 5. Factors loading in P. cupreus traits in PCA (males)
| Признаки | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 |
| Длина надкрылий | 0.46 | 0 | 0.49 | 0.22 | –0.24 | –0.66 |
| Ширина надкрылий | 0.11 | –0.92 | 0.22 | –0.26 | 0.01 | 0.15 |
| Длина переднеспинки | 0.49 | 0.04 | –0.03 | 0.09 | 0.86 | 0.04 |
| Ширина переднеспинки | 0.43 | 0.34 | 0.42 | –0.35 | –0.24 | 0.58 |
| Длина головы | 0.43 | –0.18 | –0.41 | 0.63 | –0.33 | 0.33 |
| Расстояние между глазами | 0.4 | 0.02 | –0.6 | –0.6 | –0.17 | –0.31 |
| Доля объясненной дисперсии (%) | 42 | 17.2 | 15 | 10.2 | 8.4 | 7.2 |
Н
Обсуждение
Изменчивость размеров жужелиц изучается, как правило, на уровне сообществ. В любом биотопе сообщество включает примерно 10–50 видов жужелиц. Просчитывается число особей видов мелких, средних и крупных размеров и строятся кривые распределения размеров всех видов. По полученным данным судят о степени нарушенности биотопа, его лесистости и т. д. (Sustek, 1987; Lelaska, Durbes, 2009; Hanson et al., 2016). Такой же межвидовой анализ примянется в других целях. В частности, в широтном градиенте было показано, что кривая изменчивости размеров карабид горбообразна: размеры увеличиваются от Северной Африки до Центральной Европы, а затем вновь уменьшаются по направлению к высоким широтам (Homburg et al., 2012). При мониторинговых исследованиях влияния возмущающих факторов на размеры жужелиц был даже рассчитан индекс – «средняя индивидуальная биомасса», который считается индикатором сукцессионного состояния окружающей среды и применяется достаточно широко (Szyszko et al., 2000). Данных по внутривидовой широтной изменчивости жужелиц несравненно меньше, и получены они в локальных масштабах (Филиппов, 2008; Evans, 1977). Некоторым исключением можно признать публикацию, где авторы с привлечением собственных и литературных данных приводят кривые изменчивости длины надкрылий жужелиц в широтном градиенте (Sukhodolskaya, Saveliev, 2016), в которой отмечается родоспецифичность проявления широтных клинов. Полученные нами данные говорят о том, что изменчивость разных отделов тела у P. cupreus в широтном градиенте может быть разнонаправленной. Такие выводы согласуются с мнением M. Shelomi (2012, p. 516), который, проанализировав большой массив данных по широтной изменчивости артропод, отмечает, что «в зависимости от того, меряете ли вы крыло или ногу, отбираете выборки в пределах страны или континента – выводы по соответствию изменчивости в данном организме правилу Бергманна могут быть совершенно противоположными».
Выводы по изменчивости размеров жужелиц под влиянием обитания на антропогенно нарушенных территориях достаточно противоречивы. Так, отмечалось, что размеры Carabus nemoralis уменьшаются по направлению от периферии к центру города, что, по-видимому, связано с фрагментацией мест обитания (Weller, Ganzhorn, 2004). Однако у этого же вида карабид размер жуков, обитающих в пригородной зоне, меньше, чем у особей в городе и естественных ценозах (Howe, Enggaard, 2006). Авторы связывают уменьшение размеров жужелицы Carabus nemoralis в пригородах с тем, что антропогенный пресс в пригородах сильнее по сравнению с городскими или естественными местообитаниями. Причем в большей степени он сказывается на размерах самок. Авторы считают, что самки жужелиц более вариабельны в размерах в связи с большими затратами из-за продукции яиц. В подобной работе С. Венна (Venn, 2007) показано, что в нарушенных местообитаниях не самки становятся меньше, а увеличивают свой размер самцы. И в том, и в другом случаях стирается половой диморфизм по размерам, поскольку у жужелиц, как правило, самки больше самцов. Насколько уменьшение размеров жуков определяется загрязнением среды, остается невыясненным, поскольку, по данным одних авторов, морфометрические признаки жужелиц не изменяются даже в условиях повышенной поллютантной нагрузки (Гонгальский, Бутовский, 1999), а по другим – выявлено, что кормление жуков куколками мух, которые питались пищей с тяжелыми металлами, приводит к уменьшению длины элитр, и, по-видимому, на загрязненных территориях у жужелиц снижается сопротивляемость пестицидам и потребление пищи (Maryanski et al., 2002).
Такие противоположные результаты разных авторов, оценивающих изменчивость размеров жужелиц в градиенте антропогенного воздействия, объясняются, на наш взгляд, несколькими причинами. Во-первых, реакция на антропогенный пресс видоспецифична. Как было показано, даже экологически близкие виды жужелиц по-разному реагируют на степень антропогенного воздействия. К примеру, Carabus granulatus уменьшается в размерах при обитании в пригороде, а Carabus cancellatus – в городе (Суходольская, Савельев, 2012, 2014). Во-вторых, карабидологи при оценке изменчивости размеров берут в анализ, как правило, один признак – или длину элитр, или общую длину тела насекомого, не учитывая факт, что изменчивость отдельных органов жужелиц может быть разнонаправленной. Это было показано как в нашей работе для P. cupreus, так и ранее для других видов насекомых (Stillwell et al., 2010; Sukhodolskaya, 2014). В-третьих, не учитывается половая дифференциация фенотипической пластичности мерных признаков: реакция самок и самцов на действие какого-либо фактора среды у жужелиц одного и того же вида может быть различной (Суходольская, Еремеева, 2013), что приводит к прямо противоположным выводам при интерпретации данных о размерной изменчивости при наличии явных сдвигов в половой структуре популяций. И, наконец, в-четвертых: несмотря на широкое использование (в зарубежных исследованиях) многомерного анализа (PCA, MDS, MANOVA, GLM), большинство исследований по карабидам узконаправленны и экспериментальны. Они дают детальную информацию, которая не может быть использована для формулирования обобщающих выводов, применимых к разным видам или родам жужелиц.
Заключение
Полученные данные по изменчивости размеров жужелицы P. cupreus свидетельствуют о том, что размеры надкрылий у этого вида увеличиваются по направлению к высоким широтам, а размеры переднеспинки уменьшаются. Вклад урбанизации в изменчивость размеров выражается в статистически значимом уменьшении размеров практически всех отделов жука. При этом изменчивость размеров самок и самцов может быть разнонаправленной.
Клинальная изменчивость морфологических признаков у видов, распространенных вдоль климатических градиентов, часто представляет трудности для таксономии, поскольку выбираемые морфологические признаки меняются под действием факторов среды. При проведении биоиндикационных и мониторинговых работ в анализ следует брать несколько признаков, поскольку изменчивость разных признаков при действии одного и того же фактора среды может быть разнонаправленной, а также следует учитывать половую структуру популяций в силу наличия различий в пластичности самок и самцов. Такие процессы ведут к тому, что в различной экологической обстановке меняется и форма жуков, что, по всей видимости, имеет приспособительное значение.
Библиография
Гонгальский К. Б., Бутовский Р. О. Жужелицы (Coleoptera, Carabidae) в окрестностях Косогорского металлургического комплекса // Проблемы почвенной зоологии: биоразнообразие и жизнь почвенной системы: Материалы V Всерос. совещания по почвенной зоологии / Под ред. Б. Р. Стригановой. М.: Т-во научных изданий КМК, 1999. С. 258–259.
Кисилев С. В. Экологические аспекты энтомофауны промышленных зон г. Тулы: Дис. … канд. биол. наук . Саранск, 2005. 178 с.
Крыжановский О. Л. Фауна СССР. Жесткокрылые . М.: Наука, 1983. 341 с.
Северцов А. Н. Главные направления эволюционного процесса . М.; Л.: Биомедгиз, 1934. 150 с.
Суходольская Р. А., Еремеева Н. И. Закономерности изменчивости размеров и формы жужелицы Carabus aeruginosus Fischer von Waldheim, 1822 (Coleoptera, Carabidae) // Сибирский экологический журнал. 2013. № 6. С. 803–812.
Суходольская Р. А., Савельев А. А. Влияние экологических факторов на морфометрическую изменчивость и половой диморфизм жужелиц (на примере Carabus cancellatus Ill.) // Прикладная энтомология. 2012. Т. 3. № 2 (8). С. 28–38.
Суходольская Р. А., Савельев А. А. Влияние экологических факторов на размерные признаки жужелицы Carabus granulatus L. (Coleoptera, Carabidae) // Экология. 2014. Т. 5. С. 369–375.
Филиппов Б. Ю. Пути адаптации и экологические закономерности освоения жужелицами (Coleoptera, Carabidae) севера Русской равнины: Автореф. дис. … д-ра биол. наук . М., 2008. 40 с.
Шварц С. С. Экологические закономерности эволюции . М.: Наука, 1980. 278 с.
Шмальгаузен И. И. Факторы эволюции . М.: Наука, 1968. 451 с.
Brown J. H., Maurer B. A. Macroecology: the division of food and space among species of continents // Science. 1989. Vol. 243. P. 1145–1150.
Brown J. H. Macroecology. Chicago: University of Chicago Press, 1995. 270 p.
Chown S. L., Gaston K. J. Body size variation in insects: a macroecological perspective // Biological Reviws. 2009. Vol. 85. Issue 1. P. 139–169.
Dangale C. D. Tiger Beetles as the Appropriate Bioindicators of Environmental Change and Pollution // Environmental Toxicants and their Effects on Species and Ecosystems: 32nd Annual Sessions of the Institute of Biology Sri Lanka. 2012. P. 55–63.
Ernsting G., Isaaks J. A. Effects of temperature and season on egg size, hatchling size and adult size in Notiophilus biguttatus // Ecological Entomology. 1977. Vol. 22. Issue 1. P. 32–40.
Evans V. G. Geographic variation, distribution and taxonomic status of the intertidal insect Thalassotrehus barbarae (Horn.) (Coleoptera, Carabidae) // Quaestiones Entomologicae. 1977. Vol. 13. P. 83–90.
Gaston K. J., Blackburn T. M. Process and Pattern in Macroecology. London: Blackwell, 2000. 214 p.
Gouws E. J., Gaston K. J., Chown S. L. Intraspecific Body Size Distributions of Insects // PLoS One. 2011. Vol. 6 (3). P. e16606. Doi: 10.1371/journal.pone.0016606 PMCID: MC3068144.
Hanson H. I., Palmu E., Birkhofer F., Smith H. G., Hedlund K. Agricultural Land Use Determines the Traits Composition og Ground Beetles Communities // PLoS ONE. 2016. Vol. 11 (1). P. eo146329.
Homburg K., Schuldt A., Drees C., Assmann T. Broad-scale geographic patterns in body size and hind wing development of western Palaearctic carabid beetles (Coleoptera: Carabidae) // Ecography. 2012. Vol. 35. P. 001–012.
Howe A., Enggaard M. Ground Beetles and urbanization // Environmental Biology – module II. Roskilde University, 2006. 88 p.
Jelaska L. S., Jesovnik A., Jelaska S. D., Pirnat A., Kucinic M., Durbesic P. Variation of carabid beetle and ant assemblages, and their morpho-ecological traits within natural temperate forests in Medvednica Natural Park // Izvorni i zvanstveni clanci. 2010. Vol. 134. P. 475–486.
Koivula M. J. Useful model organisms, indicators, or both? Ground beetles (Coleoptera, Carabidae) reflecting environmental conditions // Zookeys. 2011. Vol. 100. P. 287–317.
Jelaska L. S., Durbesic P. Comparison of the body size and wing form of carabid species (Coleoptera, Carabidae) between isolated and continuous forest habitats // Annales de la Societe Entomologique de France. 2009. Vol. 45. № 3. P. 327–338.
Lovei G. Ecology and conservation biology of Ground Beetles (Coleoptera, Carabidae) in an age of increasing human dominance. 2008. 145 p. URL: http://real-d.mtak.hu/121/1/Lovei.pdf (дата обращения 05.12.2010).
Maryanski M., Kramarz P., Laskowski R., Niklinska M. Decreased energetic reserves, morphological changes and accumulation of metals in Carabid Beetles (Poecilus cupreus L.) exposed to zinc- or cadmium-contaminated food // Ecotoxicology. 2002. Vol. 11. P. 127–139.
McGeoch M. A. The selection, testing and application of terrestrial insects as bioindicators // Biological Reviews. 1998. Vol. 73. P. 181–201.
R Development Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria, 2011. URL: http://www.R-project.org/ (дата обращения 12.02.2016).
Shelomi M. Where are we now? Bergmanns rule sensu lato in insects // Amer. Naturalist. 2012. Vol. 180 (1). № 4. P. 511–519.
Stillwell R. C., Blanckenhorn W. U., Teder T., Davodowitz G., Fox C. W. Sex Differences in Phenotypic Plasticity Affect Variation in Sexual Size Dimorphism in Insects: From Physiology to Evolution // Annual Review Entomology. 2010. Vol. 55. P. 227–245.
Sukhodolskaya R. Variation in Body Size and Body Shape in Ground Beetle Pterostichus melanarius Ill. (Coleoptera, Carabidae) // Journal of Agri-Food and Applied Sciences. 2014. Vol. 2 (7). P. 196–205.
Sukhodolskaya R., Saveliev A. Body Size Variation of Ground Beetles (Coleoptera, Carabidae) in Latitudinal Gradient // Periodicum Biologorum. 2016. Vol. 118. № 3. P. 273–280.
Sustek Z. Changes in body size structure of body size communities (Coleoptera, Carabidae) in urbanization gradient // Biologia (Bratislava). 1987. Vol. 43. Issue 2. P. 145–156.
Szyszko J., Vermuelen H. J. W., Klimaszewski M., Schwerk A. Mean Individual Biomass (MIB) of ground beetles (Carabidae) as an indicator of the state of the environment // Natural history and applied ecology of carabid beetles. Brandmayr P., Lovei G., Brandmayr T. Z., Casale A., Vigna Taglianti A. (eds.). Sofia; Moscow: Pensoft publishers, 2000. P. 289–294.
Venn S. Morphological responses to disturbance in wing-polymorphic carabid species (Coleoptera: Carabidae) of managed urban grasslands // Baltic J. Coleopterology. 2007. Vol. 7 (1). P. 51–59.
Weller B., Ganzhorn J. U. Carabid beetle community composition, body size, and fluctuating asymmetry along an urban-rural gradient // Basic and Applied Ecology. 2004. Vol. 5. P. 193–201.
Благодарности
Авторы приносят искреннюю благодарность специалистам-карабидологам – проф. Кемеровского государственного университета, д. б. н. Н. И. Еремеевой, проф. Ставропольского аграрного университета д. б. н. Е. В. Ченикаловой, в. н. с. Зоологического музея ИСиЭЖ СОРАН В. Г. Мордковичу, к. б. н. А. К. Жеребцову за предоставление материала для морфометрического анализа.

 © 2011 - 2026
© 2011 - 2026