



Введение
В связи с активным развитием рыбоводства в Республике Карелия и увеличением объемов производства рыбной продукции перед учеными все чаще встает проблема сохранения водных экосистем, на которых располагаются рыбоводные хозяйства. Одним из таких примеров является оз. Тарасмозеро. На его берегу расположен Кедрозерский рыбоводный завод, который уже на протяжении 22 лет (с 1992 года) сбрасывает в водоем загрязненные воды с форелевых выростных канав, оказывая тем самым значительную биогенную нагрузку. Проектная мощность форелевого комплекса рассчитана на выращивание 200 т товарной форели и 50 т молоди в год.
Как известно, дополнительное поступление биогенных элементов в водоем (главным образом органических соединений N, P, C) влечет за собой снижение прозрачности воды, увеличение биомассы и первичной продукции фитопланктона, а также других начальных звеньев трофической цепи. Ухудшается кислородный режим, возникают новые ассоциации перифитона, отмечается ежегодное «цветение» воды, усиленное образование детрита и заиление грунтов (Решетников, 1982).
Изменение условий существования отражается на видовом составе, соотношении таксономических групп, структуре популяций и количественных показателях зоопланктона, бентоса и ихтиофауны. Наиболее четко изменения в структуре сообщества прослеживаются на примере зоопланктона.
Целью работы было изучение динамики видового разнообразия, количественных и структурных показателей сообщества зоопланктона оз. Тарасмозера в условиях длительного антропогенного загрязнения.
Материалы
Озеро Тарасмозеро принадлежит к бассейну р. Лижмы, который расположен в средней части Карельского гидрографического района к северо-западу от Заонежского полуострова. Тарасмозеро – небольшое по площади мелководное озеро, расположено между южной оконечностью Кедрозера и Малой Лижемской губой Онежского озера. Через него протекает р. Нижняя Лижма, вытекающая из Кедрозера и впадающая в Малую Лижемскую губу. Озеро имеет удлиненную форму, вытянуто с СЗ на ЮВ. В отличие от других озер бассейна Тарасмозеро характеризуется исключительно высоким показателем условного водообмена (табл. 1).
Таблица 1. Основные морфометрические и гидрологические показатели Тарасмозера (по: Озера Карелии, 2013)
| Показатель | оз. Тарасмозеро |
| Географические координаты | 62о 23’ с. ш. ; 34о 27’ в. д. |
| Площадь зеркала, км2 | 1.1 |
| Длина, км | 2.3 |
| Ширина ср./макс., км | 0.5/0.9 |
| Глубина ср./макс., м | 3.7/5.8 |
| Показатель условного водообмена | 59.6 |
| Площадь водосбора, км2 | 703 |
| Объем водных масс, млн м3 | 4.1 |
Гидрологические показатели водоема в связи со строительством рыбоводного хозяйства практически не изменились. Только на участке р. Лижмы протяженностью 0.9 км от Кедрозера до Тарасмозера уменьшился расход воды в зависимости от рыбоводного цикла на 0.52–1.0 м3/с (проектные данные), что составляет 6.0–11.6 %, или в среднем около 8 % от среднемноголетнего расхода воды р. Лижмы.
Пробы зоопланктона отбирались с интервалом в 4 года с 1989 по 2012 год трижды за вегетационный период с 15 июня до 12 сентября на 4 постоянных станциях. Даты отбора проб по годам отличались на 10–12 дней. Станции располагались по главной оси водоема в направлении с СЗ на ЮВ, диапазон глубин составлял от 2.5 до 4.5 м. Температура воды во время исследований колебалась в пределах 15–23 оС.
Методы
Для отбора проб применялся планктобатометр объемом 2 л, при этом облавливались все слои воды с интервалом в 1 м с двукратной повторностью. Интегрированные пробы процеживались через планктонную сеть (диаметр ячеи 100 мкм), концентрировались до 100 мм3 и фиксировались 4 % формалином. Данные по численности и биомассе зоопланктона за вегетационный период каждого года усреднялись.
Обработка проб проводилась непосредственно авторами по общепринятым методикам гидробиологического мониторинга (Руководство…, 1992). Биомасса зоопланктона определялась расчетным методом (Ruttner-Kolisko, 1977; Балушкина, Винберг, 1979). Для анализа структуры зоопланктоценоза использовались следующие индикационные показатели: Bcrus/Brot, Nclad/Ncop, Bcycl/Bcal. Индекс видового разнообразия рассчитывался по формуле Шеннона – Уивера (Shannon, Weaver, 1963). Для оценки трофического статуса на основе изучения изменений в качественном составе зоопланктона нами использовался индекс трофности Е, предложенный А. Х. Мяэметсом (1979) с учетом дополнительного списка видов-индикаторов трофности, содержащегося в работе L. Hakkari (1972). Также трофический статус водоема оценивался по шкале трофности для зоопланктона по методике С. П. Китаева (1984). Систематика низших ракообразных и коловраток приводится согласно современным представлениям о таксономии планктонных беспозвоночных (Определитель…, 2010).
Результаты
За период исследований с 1989 по 2012 год cписок планктонных организмов, отмеченных нами в озере Тарасмозеро, насчитывает 60 таксонов (табл. 2). Из них коловраток (Rotifera) – 18, ветвистоусых ракообразных (Cladocera) – 29, веслоногих ракообразных Copepoda –13 (Calaniformes – 3 и Cyclopiformes – 10).
Таблица 2. Видовой состав зоопланктона Тарасмозера
| Таксон | Годы исследований | ||||||
| 1989 | 1992 | 1996 | 2000 | 2004 | 2008 | 2012 | |
| Тип Rotifera
КлассEurotatoria |
|||||||
| Сем. Synchaetidae | |||||||
| Synchaeta stylata Wierz. | − | − | + | + | − | + | + |
| S. pectinata Ehrenberg | − | − | + | − | + | + | + |
| Polyarthra minor Voight | − | − | − | − | − | + | − |
| P. luminosa Kutikova | − | − | + | + | − | + | + |
| Ploesoma truncatum (Lev.) | + | − | − | − | − | − | + |
| Bipalpus hudsoni (Imhof) | + | + | + | + | + | + | + |
| Сем. Asplanchnidae | |||||||
| Asplanchna priodonta Gosse | + | + | + | + | + | + | + |
| Сем. Euchlanidae | |||||||
| Euchlanis dilatata Ehrenberg | − | − | − | + | + | + | + |
| E. triquetra Ehrenberg | + | + | − | + | + | − | + |
| E. deflexa Gosse | − | − | − | − | − | − | + |
| Сем. Brachionidae | |||||||
| Keratella cochlearis Gosse | + | + | + | + | − | + | − |
| K. serrulata (Ehrenberg) | − | − | + | − | − | − | − |
| K. quadrata (Muller) | − | + | + | − | − | − | − |
| Kellicottia longispina (Kel.) | + | + | + | + | + | + | + |
| Notholca caudata Carlin | + | + | - | − | − | − | − |
| Сем. Conochilidae | |||||||
| Conochilus unicornis Rousselet | − | + | + | − | + | + | + |
| Сем. Testudinellidae | |||||||
| Testudinella patina (Hermann) | − | − | + | − | + | − | + |
| Сем. Filinidae | |||||||
| Filinia longiseta (Ehrenberg) | − | − | − | + | − | + | + |
| Тип ArthropodaПодтип Branchiata
Класс Crustacea Подкласс Branchiopoda Отряд Cladocera |
|||||||
| Сем. Sididae | |||||||
| Sida crystallina (O.F. Muller) | + | + | − | − | + | + | + |
| Limnosida frontosa Sars | − | + | + | + | − | + | + |
| Latona setifera O.F. Muller | + | − | − | − | + | − | − |
| Diaphanosoma brachyurum (Lievin) | − | − | + | − | + | − | + |
| Сем. Holopedidae | |||||||
| Holopedium gibberum Zaddach | + | + | + | + | − | + | − |
| Сем. Daphniidae | |||||||
| Daphnia longispina O.F. Muller | + | + | + | + | + | + | + |
| D. cucullata Sars | + | − | − | − | + | − | − |
| D. cristata Sars | + | − | − | − | + | + | + |
| Ceriodaphnia quadrangula (O.F.Muller) | + | + | + | + | − | − | + |
| Сем. Macrothricidae | |||||||
| Ophryoxus gracilis Sars | + | + | + | + | + | + | − |
| Сем. Chydoridae | |||||||
| Eurycercus lamellatus (O.F. Muller) | + | + | + | + | + | + | − |
| P. truncatus (O.F. Muller) | + | − | − | + | − | − | + |
| P. denticulatus (Birge) | + | + | − | − | − | + | − |
| Chydorus sphaericus (O.F. Muller) | + | + | + | + | + | + | + |
| Pseudochydorus globosus Baird | + | − | − | − | − | − | − |
| Alona quadrangularis (O.F. Muller) | + | + | − | + | + | − | + |
| Biapertura affinis Leydig | − | − | − | − | + | + | − |
| Acroperus harpae (Baird) | + | + | + | + | + | + | + |
| A. elongatus elongatus (Sars) | − | − | + | + | - | + | + |
| Graptoleberis testudinaria (Ficher) | + | − | − | − | − | − | + |
| Rhynchotalona falcata (Sars) | − | − | − | − | + | − | + |
| Camptocercus restirostris Schoedler | + | − | − | − | − | − | − |
| Сем. Bosminidae | |||||||
| Bosmina (Bosmina) longirostris (O.F. Muller) | − | + | + | + | + | + | + |
| B. longispina Leydig | + | + | + | − | − | − | − |
| B. (Eubosmina) cf. coregoni Baird | + | + | + | + | − | − | − |
| B. (Eubosmina) cf. kessleri Uljanin | + | + | − | + | − | − | − |
| Сем. Polyphemidae | |||||||
| Polyphemus pediculus (Linne) | + | + | − | − | + | + | + |
| Bythotrephes brevimanus Leydig | − | − | − | + | − | − | − |
| Сем. Leptodoridae | |||||||
| Leptodora kindtii (Focke) | − | − | − | − | − | + | + |
| Подкласс Maxillopoda
Отряд Copepoda Подотряд Calanoida |
|||||||
| Сем. Diaptomidae | |||||||
| Eudiaptomus gracilis Sars | + | + | + | + | + | + | + |
| Сем. Temoridae | |||||||
| Eurytemora lacustris (Poppe) | + | − | + | − | − | − | − |
| Heterocope appendiculata Sars | + | − | + | − | − | + | - |
| Подотряд Cyclopoida | |||||||
| Сем. Cyclopidae
Подсемейство Eucyclopinae |
|||||||
| Macrocyclops albidus (Jurine) | + | − | − | − | − | − | + |
| Eucyclops serrulatus Lilljeborg | − | − | − | − | + | + | + |
| E. serrulatus var. proximus (Lilljeborg) | - | − | − | − | + | + | + |
| Paracyclops fimbriatus (Fisher) | − | − | − | - | − | + | + |
| Подсемейство Cyclopinae | |||||||
| Cyclops strenuus Fisher | + | + | + | + | + | + | + |
| C. vicinus Uljanin | − | − | − | + | − | − | − |
| Megacyclops viridis (Jurine) | + | + | − | − | + | − | + |
| M. gigas (Claus) | + | − | − | − | − | + | + |
| Mesocyclops (s. str.) leuckarti Claus | + | + | + | + | + | + | + |
| Thermocyclops oithonoides Sars | + | + | + | + | + | + | + |
Анализ динамики видового богатства зоопланктона Тарасмозера показывает, что если в 1989 году доля коловраток от общего числа видов составляла 18 %, ветвистоусых – 51 %, веслоногих – 31 %, то в настоящее время – 33 , 43 и 25 % соответственно (табл. 3).
Таблица 3. Количество таксономических единиц зоопланктона Тарасмозера по группам
| Группы | 1989 | 1992 | 1996 | 2000 | 2004 | 2008 | 2012 |
| Rotifera | 7 | 8 | 9 | 8 | 7 | 11 | 13 |
| Cladocera | 20 | 17 | 14 | 16 | 16 | 16 | 17 |
| Calanoida | 3 | 1 | 3 | 1 | 2 | 2 | 1 |
| Cyclopoida | 9 | 4 | 3 | 5 | 6 | 7 | 9 |
| Всего | 40 | 30 | 31 | 29 | 31 | 36 | 40 |
Также нами были выявлены изменения в соотношении количественных показателей основных групп (табл. 4, 5). Так, в конце 1980-х - начале 1990-х годов по численности преобладали ветвистоусые и веслоногие ракообразные (35 и 55 % соответственно), а доля коловраток колебалась около 10 %. В то же время преобладание крупных форм среди ветвистоусых определяло их доминирование по биомассе в зоопланктоне озера – 75 %, на долю веслоногих приходилось 22 %, на долю коловраток – 3 %.
Таблица 4. Динамика численности зоопланктона Тарасмозера в летний период в среднем за 1989–2012 годы (тыс. экз/м3)
| Год | Показатель | Rotifera | Cladocera | Copepoda | Всего |
| 1989 | N | 1.6 | 4.6 | 4.8 | 11.0 |
| % | 14.6 | 41.8 | 43.6 | 100 | |
| 1992 | N | 0.96 | 4.8 | 11.54 | 17.3 |
| % | 5.6 | 27.7 | 66.7 | 100 | |
| 1996 | N | 3.7 | 4.3 | 2.3 | 10.3 |
| % | 35.2 | 40.9 | 23.9 | 100 | |
| 2000 | N | 6.9 | 5.8 | 1.0 | 13.7 |
| % | 50.4 | 42.3 | 7.3 | 100 | |
| 2004 | N | 1.8 | 5.2 | 3.1 | 10.1 |
| % | 17.8 | 51.5 | 30.7 | 100 | |
| 2008 | N | 2.0 | 6.8 | 5.2 | 14.0 |
| % | 14.3 | 48.6 | 37.1 | 100 | |
| 2012 | N | 3.4 | 5.8 | 4.2 | 13.4 |
| % | 25.2 | 43.4 | 32.4 | 100 |
Относительно низкий вклад копепод в образовании общих показателей биомассы объясняется преобладанием в этой группе мелких видов р. Mesocyclops – M. oithonoides и M. leuckarti. Коловратки были представлены главным образом K. longispina и A. priodonta, которые встречались повсеместно и существенной роли в образовании биомассы зоопланктона не играли.
Таблица 5. Динамика биомассы зоопланктона Тарасмозера в летний период в среднем за 1989–2012 гг. (мг/м3)
| Год | Показатель | Rotifera | Cladocera | Copepoda | Всего |
| 1989 | B | 11.5 | 495.9 | 40.3 | 547.7 |
| % | 2.1 | 90.5 | 7.4 | 100 | |
| 1992 | B | 31.0 | 385.0 | 232.0 | 648.0 |
| % | 4.8 | 59.4 | 35.8 | 100 | |
| 1996 | B | 51.0 | 512.2 | 18.6 | 581.8 |
| % | 8.8 | 88.0 | 3.2 | 100 | |
| 2000 | B | 111.4 | 708.9 | 29.8 | 850.1 |
| % | 13.1 | 83.4 | 3.5 | 100 | |
| 2004 | B | 50.0 | 672.0 | 52.0 | 774.0 |
| % | 6.5 | 86.8 | 6.7 | 100 | |
| 2008 | B | 90.0 | 840.0 | 80.0 | 1010.0 |
| % | 9.0 | 83.0 | 8.0 | 100 | |
| 2012 | B | 58.7 | 542.2 | 310.3 | 911.2 |
| % | 6.4 | 59.5 | 34.1 | 100 |
Обсуждение
Анализ многолетней динамики видового состава зоопланктона в Тарасмозере показал, что летний комплекс руководящих компонентов планктонной фауны сравнительно невелик и более или менее однороден по всей акватории. Он слагается из небольшого числа видов, имеющих в Карелии широкое распространение и отличающихся высокой численностью в озерах различных лимнологических типов. В озере четкого разграничения экологических комплексов зоопланктона на пелагический, прибрежно-зарослевый и придонный не наблюдается. Даже в центральной зоне озера обычными компонентами зоопланктона становятся обитатели зарослевого прибрежья.
Многие придонные виды довольно часто встречаются по всей толще воды, что характерно и для большинства мелких озер Северо-Запада России (Салазкин, 1976). Основной фаунистический комплекс зоопланктона составляют эвритопные организмы, имеющие широкую амплитуду приспособленности к меняющимся условиям обитания. Летом, по сравнению с весенним периодом, в составе зоопланктона появляется значительно больше видов ветвистоусых (Sida crystallina, Graptoleberis testudinaria, Ceriodaphnia quadrangula, до 4 видов и форм р. Bosmina и др.).
До начала работы форелевого комплекса в 1992 году, Тарасмозеро относилось к классу олиготрофных водоемов со среднелетней биомассой зоопланктона до 500 мг/м3 (Кучко, 2004; Куликова, 2007). Основа биомассы (от 50 до 90 %) создавалась за счет ветвистоусых ракообразных, главным образом видов р. Bosmina. На долю циклопид (Mesocyclops leuckarti, Thermocyclpos oithonoides, Cyclops strenuus) приходилось в среднем от 10 до 40 %. Зоопланктон литорали отличался от пелагического повышенным видовым разнообразием коловраток (до 7 видов). В центральной части озера на глубинах 3–4.5 м зоопланктон по числу встречающихся видов и количественным показателям мало отличался от литорального (1.5–2 м), однако биомасса в целом складывалась за счет более крупных объектов, поскольку здесь значительно ниже численность коловраток и мелких циклопид, а удельный вес крупных калянид (Eudiaptomus gracilis, Heterocope appendiculata) увеличивался до 10 %.
После начала работы форелевого комплекса (1992 год) наиболее заметные изменения видового состава зоопланктона в Тарасмозере произошли в группе коловраток. За период с 1989 по 2012 год число их видов увеличилось с 7 до 13 (рисунок). Видовое разнообразие и обилие коловраток в водоеме определяется напряженностью конкурентных отношений за пищевые ресурсы, а также воздействием различных факторов среды. В благоприятных условиях, размножаясь партеногенетически, они быстро достигают высокой численности, однако ввиду мелких размеров и веса (большинство имеют индивидуальный вес менее 1 мкг) значения общей биомассы коловраток редко превышают несколько десятков миллиграмм в 1 м3. Общими за все время исследований являлись такие виды, как Kellicottia longispina, Keratella cochlearis, Asplanchna priodonta, Bipalpus hudsoni, которые входят в состав руководящих видов зоопланктона большинства карельских водоемов и распространены повсеместно. Начиная с 1996 года в пробах стали отмечаться коловратки, относимые к группе индикаторов повышенной трофности, – Synchaeta stylata, S. pectinata, Polyarthra luminosa.
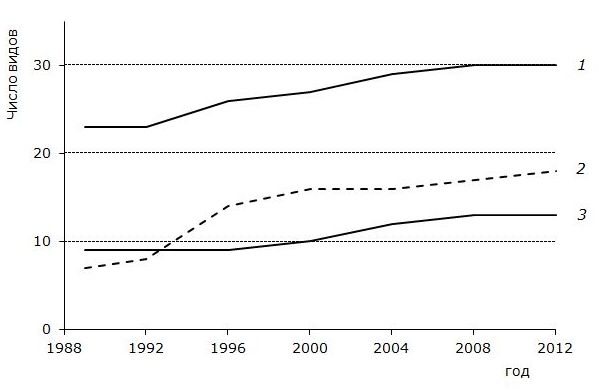
Многолетняя динамика видового богатства зоопланктона оз. Тарасмозеро. 1 – Cladocera, 2 – Rotatoria, 3 – Copepoda
Long-term dynamics of species diversity of zooplankton in Lake Tamasozero . 1 – Cladocera, 2 – Rotatoria, 3 – Copepoda
Появление этих видов в планктоне часто указывает на увеличение биогенной нагрузки на водоем или наличие источников органического загрязнения (Андроникова, 1996; Думнич, 2000). Численность некоторых холодноводных олигосапробных видов коловраток заметно снизилась и к концу 1990-х годов в Тарасмозере фактически перестали встречаться такие виды, как Notholca caudata, Keratella quadrata. Это свидетельствует о более выраженной реакции коловраток, по сравнению с другими группами, на изменение условий в озере за истекший период.
Так, до ввода в действие форелевого хозяйства, в 1989–1992 годах заметную роль в сообществе (до 20 % по численности) играла коловратка Conochilus unicornis (олиго-мезотрофный вид), которая к 1997 году выпала из субдоминантов, а в настоящее время этот вид отличается незначительной численностью. Постепенное усиление органической нагрузки на водоем привело к увеличению относительной численности K. cochlearis наряду с K. longispina до 18 % к 2004 году, а также заметному развитию видов из сем. Synchaetidae. Повышение роли этого комплекса наблюдается в настоящее время и сказывается на величине показателя сапробности воды Тарасмозера.
Менее заметные изменения видового богатства произошли в группе ветвистоусых ракообразных. В 1989 году было отмечено максимальное количество кладоцер – 20 видов. Это объясняется тем, что в видовой список, в отличие от последующих лет, также вошли организмы, отмеченные в питании молоди плотвы, окуня и уклеи в литорали северной части водоема (в основном бентические зарослевые виды сем. Chydoridae – Graptoleberis testudinaria, Camptocercus restirostris), которые малочисленны и не всегда улавливаются стандартными орудиями лова. В дальнейшем количество видов ветвистоусых по годам колебалось в пределах 14–17. После ввода в эксплуатацию форелевого хозяйства в 1992 году отмечается снижение численности некоторых пелагических и эвритопных олиго-мезотрофных видов сем. Bosminidae – Bosmina longispina, B. coregoni, B. kessleri. С другой стороны, возрастает обилие видов зарослевого зоопланктоценоза – главным образом за счет крупных Sida crystallina, Eurycercus lamellatus, Latona setifera, Alona quadrangularis, Polyphemus pediculus, видов рода Acroperus.
Среди веслоногих ракообразных в последние годы исследований (2008, 2012) в пробах стали встречаться некоторые бентические зарослевые виды, не отмеченные ранее – Eucyclops serrulatus и Paracyclops fimbriatus. В то же время из списка пелагического копеподного планктона начиная с 2000 года выпала Eurytemora lacustris. В остальном существенных изменений в группе зарегистрировано не было и веслоногие сохранили относительную стабильность видового состава на протяжении всего периода исследований.
По величине индекса трофности Мяэметса (табл. 6) Тарасмозеро постепенно переходит в разряд мезотрофных водоемов (0.5–1.0). Этому способствуют малые размеры водоема и постоянное присутствие в планктоне коловраток семейства Synchaetidae и зарослевых форм Cladocera (главным образом представителей семейства Chydoridae), являющихся в массе своей индикаторами мезотрофии. В составе зоопланктона также встречаются виды, которые, по свидетельству ряда авторов, в условиях умеренных широт служат индикаторами повышенной трофности, – Polyarthra luminosa, Filinia longiseta, Trichocerca insignis, Daphnia longispina, Cyclops kolensis (Рылов, 1948; Мануйлова, 1964; Кутикова, 1970; Pejler, 1965). Полученные нами данные хорошо согласуются с результатами продолжительных исследований на двух озерах южной Карелии – Сямозере и Пертозере. На Сямозере показатель Е за 20 лет (с 1955 по 1979 год) увеличился в 2.9 раза и составил 0.52, за эти годы трофический статус Сямозера изменился, и по уровню количественного развития зоопланктона озеро перешло из олиготрофного в мезотрофное состояние (Куликова, 2004). Пертозеро, в результате воздействия схожих эвтрофирующих факторов (сельскохозяйственные стоки с водосбора, воздушный перенос, строительство дачных кооперативов на побережье, усиление рекреационной нагрузки), также стало относиться к разряду мезотрофных водоемов и к 1989 году индекс трофности составил 0.86 (Тимакова, 1997). Результаты оценки качества воды по методу Пантле и Букка (Макрушин, 1974; Куликова, 1983), основанной на индикаторной значимости организмов и частоте их встречаемости, сопоставимы с данными по уровню трофности. После начала работы форелевого хозяйства значения индекса стали постепенно возрастать, отражая изменения в видовом составе и структуре зоопланктоценоза.
После начала работы форелевого хозяйства было отмечено некоторое увеличение численности коловраток, в 2000 году их доля составляла около 50 % от общего количества зоопланктона. Доминировали в основном некрупные коловратки сем. Synchaetidae и Brachionidae – S. pectinata, K. longispina, K. cochlearis. Однако в дальнейшем ожидаемого роста численности коловраток не произошло, к 2008 году их доля в общем количестве зоопланктона практически осталась на уровне 1989–1992 годов и составила всего 14.3 %. Это можно объяснить изменением трофической структуры группы коловраток. Начиная с 2004 года отмечается тенденция к увеличению количественных показателей хищного крупноразмерного вида A. priodonta (до 20 % численности и 70 % от общей биомассы коловраток). Удельный вес коловраток в общей биомассе зоопланктона также возрос и в 2008 году составил 9 %. Вместе с тем следует отметить, что в некоторых мезотрофных водоемах доля A. priodonta в общей биомассе коловраток значительно выше и достигает 88–95 %, а ее вклад в тотальную биомассу зоопланктона составляет от 25 до 40 % (Макарцева, 1988; Думнич, 2000).
В группе ветвистоусых ракообразных были отмечены следующие изменения. При общем сохранении доминирования кладоцер по биомассе за весь период исследования в результате перестройки внутренней структуры произошли изменения в комплексе ценозообразующих видов. Начиная с 1996–1997 годов в группу доминант входят в основном зарослевые виды – S. crystallina, E. lamellatus, A. quadrangularis, Ch. sphaericus, а их вклад в создание общей биомассы зоопланктона увеличился с 22 до 50 %. Подобное явление характерно для удобряемых малых водоемов (площадью до 300 га) Северо-Запада России, на которых развито интенсивное озерное рыбоводство (Моисеева, Алексеев, 1986). Это говорит о том, что в малых водоемах литоральный зоопланктон в большей мере реагирует на увеличение притока биогенов, чем пелагический. Интенсивное развитие крупных зарослевых видов отразилось на показателе средней индивидуальной массы зоопланктеров, который за период исследования возрос и в 2012 году составил 0,068 мг/особь (табл. 6).
Таблица 6. Динамика структурных показателей зоопланктона оз. Тарасмозера
| Год | 1989 | 1992 | 1996 | 2000 | 2004 | 2008 | 2012 |
| Wср. (мг/особь) | 0.049 | 0.037 | 0.056 | 0.062 | 0.076 | 0.071 | 0.068 |
| Bcrus/Brot | 46.6 | 19.9 | 10.4 | 6.6 | 14.5 | 10.2 | 14.5 |
| Bcycl/Bcal | 2.82 | 2.11 | 3.24 | 4.74 | 6.65 | 5.58 | 9.36 |
| Nclad/Ncop | 0.71 | 3.3 | 1.9 | 5.8 | 1.7 | 1.3 | 1.4 |
| Индекс трофности | 0.44 | 0.43 | 0.42 | 0.50 | 0.58 | 0.75 | 0.71 |
| Индекс сапробности | 0.95 | 1.10 | 1.15 | 1.12 | 1.26 | 1.37 | 1.42 |
| Индекс Шеннона, бит/экз. | 1.68 | 1.70 | 1.70 | 1.71 | 1.73 | 1.65 | 1.72 |
В группе веслоногих ракообразных снижается доля диаптомид и возрастает доля циклопид преимущественно за счет видов M. leuckarti и Th. oithonoides, величина индекса Bcycl/Bcal за период исследований увеличилась с 2.82 до 9.36. Анализ динамики других структурных показателей позволяет расценивать происходящие изменения в сообществе зоопланктона Тарасмозера как результат длительного воздействия сточных вод форелевого хозяйства. Также с начала 2000-х годов на территории частного водосбора Тарасмозера развивается дачное строительство, что может приводить к дополнительному поступлению в водоем биогенных веществ. Показатель Bcrus/Brot после ввода в действие форелевого хозяйства заметно снизился, что указывает на возрастание роли коловраток в образовании общей биомассы зоопланктона и функционировании сообщества в целом. Несмотря на это отмечается увеличение абсолютных показателей биомассы в среднем за вегетационный период до 1000 мг/м3 и среднего веса зоопланктеров до 0,068 мг/особь к 2012 году. Показатель Nclad/Ncop по годам отличается незначительно (за исключением 2000 года) и отражает определенную стабильность численности основных групп зоопланктона на протяжении всего периода исследований.
Заключение
1. В Тарасмозере после ввода в эксплуатацию форелевого хозяйства (1992 год) отмечено незначительное обогащение видового состава зоопланктона за счет представителей Rotatoria и Cladocera. Было зафиксировано появление 5 видов зоопланктона, не встречавшихся ранее, большинство из которых являются индикаторами мезо- и эвтрофных условий по системе сапробности Пантле – Букка.
2. Показано увеличение индекса трофности Мяэметса, отражающего изменения в соотношении как таксономических групп, так и индикаторных организмов. По величине данного показателя Тарасмозеро перешло в разряд мезотрофных водоемов.
3. По уровню количественного развития зоопланктона Тарасмозеро постепенно переходит от олиготрофного к мезотрофному типу со среднелетней биомассой около 1 г/м3.
4. За время исследований отмечены структурные перестройки сообщества зоопланктона, также свидетельствующие о повышении трофического статуса водоема. В Тарасмозере, которое характеризуется малой площадью, незначительными глубинами, высоким коэффициентом водообмена и развитой высшей водной растительностью, происходит интенсивное развитие зарослевого зоопланктоценоза (в основном крупные формы Cladocera и некоторые виды Rotifera), что отражается на увеличении средней индивидуальной массы зоопланктеров. Для уточнения причин отмеченных явлений целесообразно рекомендовать проведение альгологических и гидрохимических исследований на водоеме в течение вегетационного периода.
Библиография
Андроникова И. Н. Структурно-функциональная организация зоопланктона озерных экосистем разных тропических типов . СПб.: Наука, 1996. 189 с.
Думнич Н. В. Ракообразные (Crustacea) и коловратки (Rotatoria) крупных озер Вологодской области : автореф. дисс. ... канд. биол. наук. Петрозаводск, 2000. 25 с.
Китаев С. П. Экологические основы биопродуктивности озер разных природных зон . М.: Наука, 1984. 207 с.
Куликова Т. П. Зоопланктон водоемов бассейна реки Шуи (Карелия) . Петрозаводск: КарНЦ РАН, 2004. 124 с.
Куликова Т. П. Зоопланктон водных объектов бассейна Онежского озера . Петрозаводск: Карельский научный центр РАН, 2007. 223 с.
Кутикова Л. А. Коловратки фауны СССР (Rotatoria) . Л.: Наука, 1970. 744 с.
Кучко Я. А. Влияние форелевого хозяйства на сообщество зоопланктона озерно-речной экосистемы : автореф. дисс. … канд. биол. наук. Петрозаводск, 2004. 26 с.
Лазарева Н. Б. Протозойный планктон и качество воды в Петрозаводском Онего // Петрозаводское Онего и его лимнологические особенности. Петрозаводск: Изд-во КарНЦ РАН, 1994. С. 108–122.
Мануйлова Е. Ф. Ветвистоусые рачки фауны СССР . М.: Наука, 1964. 327 с.
Макарцева Е. С. Изменения структуры и качественных показателей зоопланктона при повышении уровня трофии озер // Изменения структуры экосистем озер в условиях возрастающей антропогенной нагрузки. Л.: Наука, 1988. С. 221–241.
Макрушин А. В. Биологический анализ качества вод . Л.: ЗИН АН СССР, 1974. 59 с.
Моисеева И. Г., Алексеев В. Р. Фауна ветвистоусых и веслоногих ракообразных литоральной зоны малых озер Северо- Запада СССР // Исследования пресноводных и морских беспозвоночных : Труды ЗИН АН СССР. 1986. Т. 152. С. 116–128.
Мяэметс А. Х. Качественный состав пелагического зоопланктона как показатель трофности озера // Изучение и освоение водоемов Прибалтики и Белоруссии: тезисы докладов 20-й науч. конф. Рига: Изд-во Тарту, 1979. Т. 1. С. 12–15.
Озера Карелии: Справочник / под ред. Н. Нcopepods . Филатова, В. И. Кухарева. Петрозаводск: КарНЦРАН, 2013. 464 с.
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 1. Зоопланктон / под ред. В. Р. Алексеева, С. Я. Цалолихина. М.: Товарищество научных изданий КМК, 2010. 495 с.
Пидгайко М. Л. Зоопланктон водоемов Европейской части СССР . Л.: Наука, 1984. 207 с.
Решетников Ю. С. Изменение озерных экосистем // Изменение структуры рыбного населения эвтрофируемого водоема. М.: Наука, 1982. С. 5–12.
Руководство по гидробиологическому мониторингу пресноводных экосистем / под ред. В. А. Абакумова. СПб.: Наука, 1992. 318 с.
Рылов В.М. Cyclopoida пресных вод . М.; Л.: Изд-во АН СССР, 1948. Т. 3. Вып. 3. 319 с.
Салазкин А. А. Основные типы озер гумидной зоны и их биолого-продукционная характеристика . М.: Изв. ГосНИОРХ, 1976. Т. 108. 194 с.
Тимакова М. В. Коловратки и ракообразные оз. Пертозеро (структура, динамика, продуктивность) : автореф. дисс. … канд. биол. наук. Петрозаводск, 1997. 24 с.
Hakkari L. Zooplankton species as indicators of environment // Aqua Fennica. Helsinki. 1972. P. 46–54.
Pejler B. On long – term stability of zooplankton composition // Scr. Limnol. Uppsala. 1975. Bd. II. P. 107–117.
Ruttner-Kolisko A. Suggestion for biomass calculation of planktonic rotifers // Arch. Hydrobiol. Ergebn. Limnol. Struttgart, 1977. H. 8. S. 71–78.
Shannon C. E., Weaver W. The mathematical theory of communication. Urbana: Univ. Illinois Press, 1963. 117 p.
Благодарности
Работа выполнена при финансовой поддержке программ ОБН РАН «Биологические ресурсы России: динамика в условиях глобальных климатических и антропогенных воздействий», Президиума РАН «Живая природа: современное состояние и проблемы развития» и Минобрнауки РФ (НШ-1410.2014.4; Соглашение 8101), гранта РФФИ №12-04-00022а.

 © 2011 - 2024
© 2011 - 2024