



Введение
Биологические инвазии признаны глобальной проблемой ввиду множества последствий для биоты и человека, в частности, их негативного влияния на нативные экосистемы и аборигенные виды (Simberloff et al., 2013; Olden et al., 2004). Однако инвазионный процесс является многоступенчатым, и в самом начале чужеродный вид должен натурализоваться, популяция вселенца должна быть способна к самовозобновлению (Sakai et al., 2001; Blackburn et al., 2011; Алимов, Богуцкая, 2004). На территории Русской равнины за последние десятилетия наблюдается экспансия многих чужеродных видов наземных моллюсков (Шиков, 2016). Среди них существенную долю занимают южные, в частности кавказские, виды. К этой группе вселенцев относится вид Xeropicta derbentina (Krynicki, 1836), естественный ареал которого охватывает Кавказ, Крым, Малую Азию (Шилейко, 1978). Его расселение на сегодняшний день наблюдается на территории северного Причерноморья, Приазовья, юга Среднерусской возвышенности, Донецкого кряжа, Приднепровской низменности и Приднепровской возвышенности, а также на северо-востоке Среднедунайской низменности (Гураль-Сверлова, Гураль, 2017; Адамова и др., 2019; Ostrovsky, 2023). Заметим, что на данный момент X. derbentina, населяющая в естественном ареале открытые степные биотопы, распространяется на территории с открытыми антропогенными ландшафтами. В частности, ранее нами были проведены исследования популяции X. derbentina, населяющей вместе с другими моллюсками-вселенцами этой же экологической группы открытый биотоп в окрестностях мелового карьера Белгорода (Adamova et al., 2022). В указанном исследовании плотность популяции X. derbentina за непродолжительный период (2017–2020 гг.) стала сокращаться, в то время как популяция другого вселенца, крымского вида Brephulopsis cylindrica (Menke, 1828), росла. Впрочем X. derbentina не исчезла с территории Среднерусской возвышенности: популяции моллюска обнаружены нами, помимо Белгорода, еще в двух населенных пунктах южной части Белгородской области (пгт. Волоконовка и г. Шебекино). Других моллюсков-вселенцев на территории этих населенных пунктов пока не наблюдалось. Типичным обитателем открытых биотопов, в т. ч. антропогенных, юга Среднерусской возвышенности является Caucasotachea vindobonensis (C. Pfeiffer, 1828) (Шилейко, 1978; Балашёв, 2016). Именно этот вид оказался соседом вселенца X. derbentina на исследованной нами территории. Взаимоотношения чужеродного и аборигенных видов могут носить разный характер, и этот аспект является одним из самых важных в инвазионном процессе. В нашем случае, с одной стороны, речь идет о видах с перекрывающимися экологическими нишами, и есть риск вытеснения нативного вида (Алимов, Богуцкая, 2004). С другой стороны, C. vindobonensis имеет широкий ареал, который частично совпадает с ареалом X. derbentina на Северном Кавказе и в Крыму, где виды успешно сосуществуют (Шилейко, 1978). Целью нашего исследования была оценка состояния двух популяций наземных моллюсков, населяющих один биотоп и имеющих сходные экологические требования: нативного вида C. vindobonensis и вселенца X. derbentina.
Материалы
Объектом исследования являлись популяции двух видов наземных моллюсков: X. derbentina и C. vindobonensis. Популяции совместно населяют рудеральный биотоп (пустырь) поблизости от железнодорожных путей. Исследование проводилось на территории юга Среднерусской возвышенности в пгт. Волоконовка Белгородской области (50°29'34.6920" N, 37°51'7.8660" E) в 2022 и 2023 гг. В 2022 г. учеты проводили ежемесячно с июня по октябрь, в 2023 г. – с мая по октябрь.
Методы
Состояние популяции мы оценивали исходя из данных о демографической структуре, плотности популяции, пространственного распределения, в т. ч. взаимного пространственного распределения популяций двух видов.
Для оценки плотности популяций, демографического состава и пространственного распределения использовали метод пробных площадок, расположенных в виде регулярной сетки. Для этого участок учета был разбит на 7 трансект по 10 площадок (0.25 м2). Расстояние между соседними площадками составляло 1 м.
Ежемесячно на каждой площадке проводили подсчет общего количества особей каждого вида для определения плотности популяции. Для оценки демографической структуры популяцию каждого вида разделяли на размерно-возрастные классы в зависимости от количества оборотов раковины или наличия сформированного отворота раковины (Шилейко, 1978; Adamova et al., 2022). Для X. derbentina было выделено 3 класса: неполовозрелые до двух оборотов раковины, неполовозрелые от 2 до 5 оборотов и половозрелые (5 и более оборотов). Для C. vindobonensis – 2 класса: неполовозрелые (не имеющие сформированный отворот раковины) и половозрелые (имеющие сформированный отворот раковины).
Оценка статистической значимости различий плотности популяций двух видов проводилась с использованием непараметрического критерия Вилкоксона – Манна – Уитни в среде R (R Core Team, 2023). Тип пространственного распределения определяли на основе анализа пространственной автокорреляции посредством вычисления глобального индекса Морана (Anselin, 1995) в программе ArcGIS 10.2. Кросс-корреляцию между популяциями двух видов оценивали на основе глобального индекса Морана с использованием пакета «spatialEco» в среде R (Chen, 2015; Evans, Murphy, 2023).
Результаты
На протяжении двух сезонов наблюдалось существенное различие плотности популяции X. derbentina и C. vindobonensis (табл. 1). Пик численности популяций обоих видов приходился на июнь – июль. Плотность популяции вида-вселенца достигала максимальных значений 200–215 особей/м2 в июле, что превосходило таковые значения для C. vindobonensis в 4–5 раз. Не было выявлено различий только в мае 2023 г., когда в среднем плотность обеих популяций составляла 12 особей/м2. В осенние месяцы плотность популяции снижалась как у X. derbentina, так и у C. vindobonensis. Однако при этом сохранялось существенное различие в пользу вселенца в среднем в 5–7 раз.
Таблица 1. Плотность населения (D) двух видов моллюсков, особей/м 2
| Xeropicta derbentina | Caucasotachea vindobonensis | |||||
| Dmean | SD | Dmax | Dmean | SD | Dmax | |
| 2022 | ||||||
| Июнь* | 20.66 | 30.75 | 169.23 | 5.05 | 8.97 | 30.77 |
| Июль* | 19.128 | 31.80 | 215.38 | 3.74 | 9.23 | 46.14 |
| Август* | 16.26 | 26.18 | 184.62 | 1.76 | 5.58 | 30.77 |
| Сентябрь* | 17.80 | 21.86 | 107.69 | 1.98 | 6.37 | 30.77 |
| Октябрь* | 10.11 | 17.56 | 92.31 | 0.44 | 2.58 | 15.38 |
| 2023 | ||||||
| Май | 12.09 | 24.97 | 107.69 | 12.09 | 24.97 | 107.69 |
| Июнь* | 27.69 | 30.16 | 123.08 | 8.79 | 14.00 | 46.15 |
| Июль* | 24.62 | 35.37 | 200 | 5.49 | 11.12 | 46.15 |
| Август* | 21.10 | 26.48 | 123.08 | 5.49 | 7.87 | 30.77 |
| Сентябрь* | 20.66 | 30.08 | 153.85 | 3.52 | 7.49 | 30.77 |
| Октябрь* | 13.63 | 21.68 | 92.31 | 1.91 | 5.81 | 30.77 |
Примечание. * – статистически значимые различия между плотностью популяций двух видов на основе теста Вилкоксона – Манна – Уитни, p < 0.05.
В обеих популяциях в течение сезона активности происходило изменение соотношения размерно-возрастных классов (рис. 1, 2), но оно имело несколько разный характер у исследуемых видов.
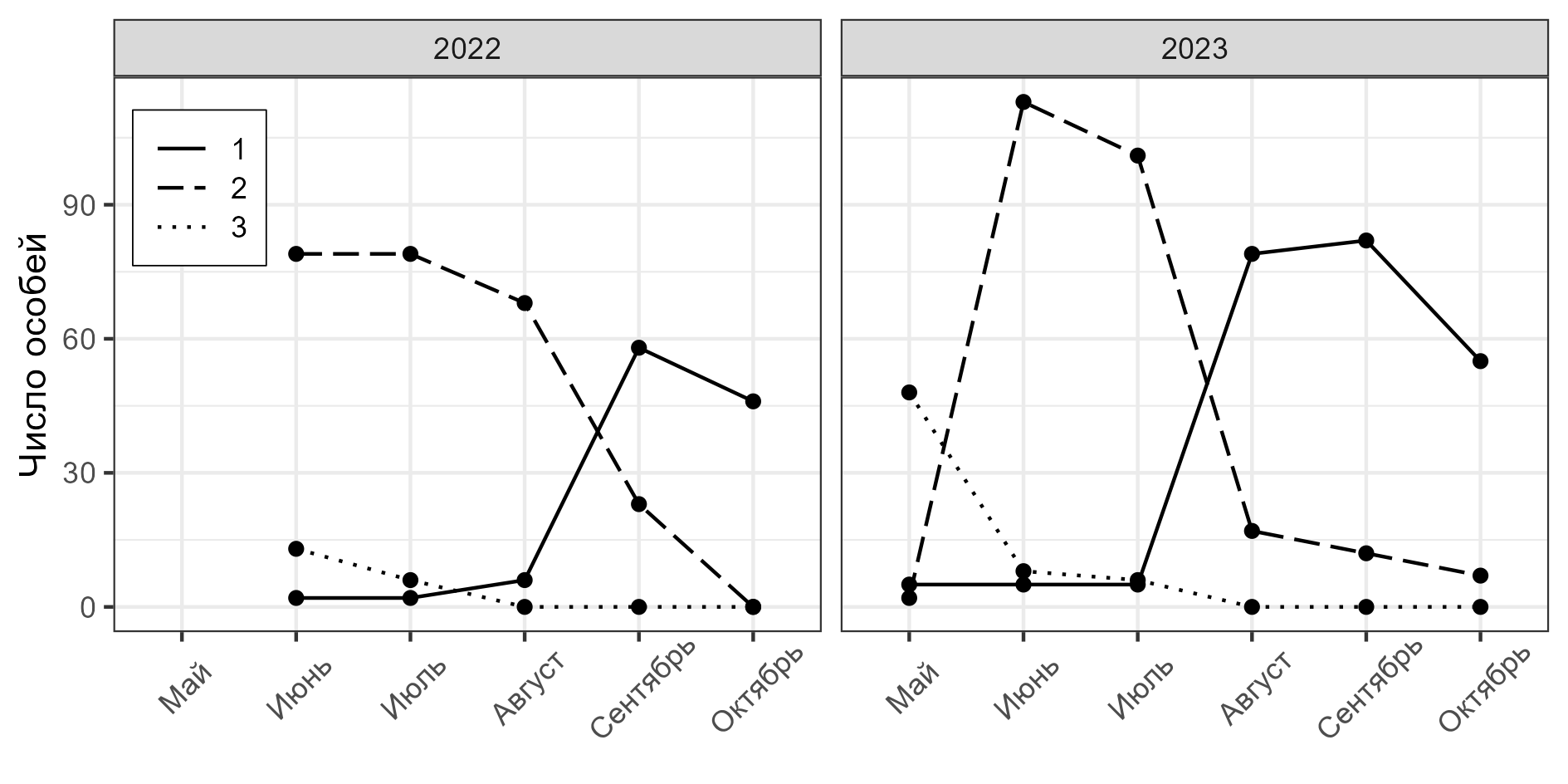
Рис. 1. Число моллюсков X. derbentina на учетных площадках: 1 – половозрелые особи, 2 – неполовозрелые особи, имеющие от 2 до 5 оборотов, 3 – неполовозрелые особи, имеющие до двух оборотов раковины
Fig. 1. Number of X. derbentina snails on the sample area: 1 – mature individuals, 2 – immature individuals, having from 2 to 5 shell whorls, 3 – immature individuals, having up to two shell whorls

Рис. 2. Число моллюсков C. vindobonensis на учетных площадках: 1 – половозрелые особи, 2 – неполовозрелые особи
Fig. 2. Number of C. vindobonensis snails on the sample areas:1 – mature individuals, 2 – immature individuals
В начале сезона активности в популяции X. derbentina появляется большое количество молодых особей, которые вылупляются из отложенных в конце прошлого сезона кладок. В течение лета они растут (увеличение доли особей 2-го класса), и осенью большую часть популяции составляют половозрелые особи. В популяции C. vindobonensis половозрелые особи составляют основную часть популяции в течение всего сезона, молодые особи присутствуют только в первой половине сезона.
Пространственное распределение особей в обеих популяциях чаще всего соответствовало случайному (табл. 2). Только в начале и конце учетов 2022 г. в популяции X. derbentina наблюдалось групповое (кластерное) распределение.
Таблица 2. Пространственное размещение моллюсков Xeropicta derbentina и Caucasotachea vindobonensis
| Вид | Год | Показатели | месяц | |||||
| май | июнь | июль | август | сентябрь | октябрь | |||
| X. derbentina | 2022 | IM | – | 0.165 | -0.016 | 0.007 | -0.006 | 0.229 |
| p-значение | – | 0.004 | 0.978 | 0.688 | 0.888 | <0.001 | ||
| Тип распределения | – | C | R | R | R | C | ||
| 2023 | IM | -0.103 | 0.042 | 0.057 | 0.0512 | 0.073 | 0.088 | |
| p-значение | 0.154 | 0.388 | 0.241 | 0.308247 | 0.165 | 0.107 | ||
| Тип распределения | R | R | R | R | R | R | ||
| C. vindobonensis | 2022 | IM | – | 0.079 | 0.021 | -0.061 | -0.007 | -0.026 |
| p-значение | – | 0.149 | 0.565 | 0.442 | 0.897 | 0.815 | ||
| Тип распределения | – | R | R | R | R | R | ||
| 2023 | IM | -0.109 | 0.030 | 0.099 | -0.040 | 0.026 | 0.083 | |
| p-значение | 0.125 | 0.496 | 0.078 | 0.694 | 0.521 | 0.108 | ||
| Тип распределения | R | R | R | R | R | R | ||
Примечание. R – случайный тип распределения, C – кластерный тип распределения, IM – глобальный индекс Морана.
Результаты анализа пространственной кросс-корреляции показали отсутствие таковой между двумя популяциями (р > 0.05). Кроме того, значения индекса Морана при оценке кросс-корреляции были невысокими: от -0.01 до 0.01.
Обсуждение
Исходя из полученных данных, можно утверждать, что популяция вида-вселенца значительно превосходит популяцию нативного вида по численности. Высокая численность X. derbentina наблюдается в других интродуцированных популяциях, например, в Провансе (Aubry et al., 2005). В ранее исследованной нами белгородской популяции X. derbentina мы наблюдали сопоставимые значения плотности на участках, где отсутствовал другой вид-вселенец (Adamova et al., 2022). Означает ли это, что X. derbentina вытесняет C. vindobonensis? Такое утверждение было бы преждевременным. У исследуемых видов отличается продолжительность жизни и жизненный цикл. X. derbentina обычно имеет однолетний жизненный цикл, хотя и способна переключаться на двулетний при определенных условиях (Kiss et al., 2005). В представленном исследовании мы наблюдали небольшое количество половозрелых особей X. derbentina в мае, так же как и в 2020 г. в другой популяции на юге Среднерусской возвышенности (Adamova et al., 2022). По всей видимости, они успешно перезимовали и потенциально могли оставить потомство в течение двух сезонов. Однако это наблюдается у небольшого количества особей в популяции. Напротив, C. vindobonensis достигает репродуктивного возраста лишь к концу второго года жизни при продолжительности жизни до семи лет (Staikou, 1998). Плотность популяции этого вида на исследуемой нами территории в сравнении с данными других работ даже можно назвать высокой: так, на севере Греции средняя плотность популяции этого вида составляла 2.80 ± 0.67 особей/м2 (Staikou, 1998). Средние значения плотности волоконовской популяции C. vindobonensis также сопоставимы с плотностью популяций Cepaea nemoralis L. (от 0.9 до 5 особей/м2) (Williamson et al., 1977). Количество ювенильных особей в волоконовской популяции C. vindobonensis также довольно большое, особенно в начале сезона активности.
Пространственная структура обеих популяций преимущественно соответствовала случайному типу распределения. В предыдущем исследовании в белгородской популяции X. derbentina при высокой плотности чаще наблюдалось агрегированное распределение особей (Adamova et al., 2022), случайное распределение было отмечено при сокращении плотности. В приведенном ранее исследовании на территории северной Греции популяция C. vindobonensis имела равномерную пространственную структуру, при этом автор указывает, что для других видов моллюсков, обитающих в этих же условиях, характерен случайный или групповой тип распределения (Staikou, 1998). Пространственное распределение наземных моллюсков зависит от многих факторов среды. На ряде видов показано влияние распределения растительности и физико-химических свойств почвы на распределение особей наземных моллюсков (Крамаренко и др., 2014). В этой же работе выявлено, для ксерофильных моллюсков характерна пространственная агрегированность. Однако в нашем исследовании только в июне и октябре 2022 г. в популяции X. derbentina наблюдалось групповое распределение особей на участке. Это может быть обусловлено изменением условий среды (например, увеличением влажности) и концентрацией моллюсков на некоторых участках. Взаимодействие особей, как внутри-, так и межвидовое, также оказывает влияние на распределение улиток (Cameron, Carter, 1979). Между исследованными нами популяциями не было выявлено взаимной пространственной корреляции, что указывает на отсутствие прямого взаимодействия между видами. Возможно, это обстоятельство, а также однородность условий среды на исследованном участке могут обуславливать случайное размещение моллюсков.
X. derbentina известна как чужеродный вид в некоторых регионах Европы, но отрицательного влияния на аборигенную малакофауну к настоящему моменту не выявлено (De Mattia, 2007; Gojšina et al., 2022; Wagner, Bertrand, 2021). В частности, в западном Средиземноморье показано даже увеличение разнообразия сообществ моллюсков в региональном масштабе, хотя отмечено, что локально вид-вселенец способен подавлять некоторые нативные виды ввиду высокой численности (Aubry et al., 2005). C. vindobonensis также успешно осваивает новые территории. Известны чужеродные популяции этого вида в центральной части Русской равнины (Egorov, 2014) и в Северной Америке (Hausdorf et al., 2021). Как и X. derbentina, C. vindobonensis проявляет экологическую пластичность и населяет разнообразные открытые биотопы (Mierzwa, 2009). Кроме того, приуроченность к антропогенным биотопам, ряд биологических особенностей (в частности, способность прикрепляться к различным предметам) обуславливают распространение этих видов человеком, что вообще характерно для многих наземных моллюсков (Cowie, Robinson, 2003). Так, распространение особей X. derbentina за счет их прикрепления к транспорту показано в северо-западном Средиземноморье (Aubry et al., 2006). Антропохорное распространение C. vindobonensis на территории Европы подтверждается молекулярно-генетическим анализом популяций (Kajtoch et al., 2017). Исследуемые нами популяции населяют прилегающую к железнодорожным путям территорию, что позволяет предположить занос вселенца с транспортом и в этом случае. Обобщая эти данные, можно предположить, что оба вида могут не только существовать на исследованной нами территории, не конкурируя друг с другом, но и расселяться на новые для них территории.
Заключение
Состояние популяций X. derbentina и C. vindobonensis на исследуемой нами территории характеризуется высокой плотностью, преобладанием ювенильных особей в начале сезона активности моллюсков, случайным размещением особей. При этом аборигенный вид C. vindobonensis не испытывает негативного воздействия со стороны вселенца. Оба вида адаптированы к обитанию в антропогенных биотопах, что позволяет предположить их дальнейшее сосуществование на территории юга Среднерусской возвышенности.
Библиография
Адамова В. В., Снегин Э. А., Украинский П. А. Морфометрическая и генетическая изменчивость популяций моллюска-вселенца Xeropicta derbentina (Gastropoda, Pulmonata, Hygromiidae) // Ruthenica. 2019. Т. 29, № 3. C. 149−160.
Алимов А. Ф., Богуцкая Н. Г. Биологические инвазии в водных и наземных экосистемах . М.: Товарищество научных изданий КМК, 2004. 436 с.
Балашёв И. А. Фауна Украины. Моллюски. Стебельчатоглазые (Stylommatophora) . Киев: Наукова Думка, 2016. Т. 29, № 5. 591 с.
Гураль-Сверлова Н. В., Гураль Р. И. Расширение ареалов наземных моллюсков рода Xeropicta (Gastropoda, Hygromiidae) на территории Украины // Российский журнал биологических инвазий. 2017. Т. 10, № 2. C. 20−27.
Крамаренко С. С., Кунах О. Н., Жуков А. В., Андрусевич Е. В. Анализ паттернов пространственной организации популяций наземных моллюсков: подход с использованием методов геостатистики // Бюллетень Дальневосточного малакологического общества. 2014. Т. 18. C. 5−40.
Шиков Е. В. Адвентивные виды наземной малакофауны центра Русской равнины // Ruthenica. 2016. Т. 26, № 3-4. C. 153−164.
Шилейко А. А. Наземные моллюски надсемейства Helicoidea. Фауна СССР. Моллюски . Л.: Наука, 1978. Т. 3, № 6. 384 с.
Adamova V. V., Ukrainskiy P. A., Krymskaya O. V. Demographic and spatial structure at the stage of expansion in the populations of some alien land snails in Belgorod city (CentralRussian Upland) // Ruthenica. Russian Malacological Journal. 2022. Vol. 32, № 1. P. 21−39.
Anselin L. Local indicators of spatial association – LISA // Geographical analysis. 1995. Vol. 27, № 2. P. 93−115.
Aubry S., Labaune C., Magnin F., Kiss L. Habitat and integration within indigenous communities of Xeropicta Derbentina (Gastropoda: Hygromiidae) a recently introduced land snail in south-eastern France // Diversity and Distributions. 2005. Vol. 11. P. 539−547.
Aubry S., Labaune C., Magnin F., Roche P., Kiss L. Active and passive dispersal of an invading land snail in Mediterranean France // Journal of Animal Ecology. 2006. Vol. 75, № 3. P. 802−813.
Blackburn T. M., Pyšek P., Bacher S., Carlton J. T., Duncan R. P., Jarošík V., Wilson J. R. U., Richardson D. M. A proposed unified framework for biological invasions // Trends in Ecology & Evolution. 2011. Vol. 26, № 7. P. 333−339.
Cameron R. A. D., Carter M. A. Intra- and Interspecific Effects of population density on growth and activity in some Helicid land snails (Gastropoda: Pulmonata) // The Journal of Animal Ecology. 1979. P. 48−237. DOI: 10.2307/4111.
Chen Y. A new methodology of spatial cross-correlation analysis // PLOS ONE. 2015. Vol. 10, № 5. P. 1−20.
Cowie R. H., Robinson D. G. Pathways of introduction of nonindigenous land and freshwater snails and slugs // Invasive species: vectors and management strategies. 2003. P. 93─122.
De Mattia W. Xeropicta derbentina (Krynicky, 1836) (Gastropoda, Hygromiidae) in Italy and along the Croatian coast, with notes on its systematics and nomenclature // Basteria. 2007. Vol. 71, № 1-3. P. 1−12.
Egorov R. The first record of Cepaea vindobonensis (Pfeiffer, 1828) (Stylommatophora: Helicidae) in the central part of European Russia // Malacologica Bohemoslovaca. 2014. Vol. 13. P. 110−113.
Evans J. S., Murphy M. A. R package version 2.0-2 // spatialEco. URL: https://github.com/jeffreyevans/spatialEco (дата обращения: 22.12.2023).
Gojšina V., Páll-Gergely B., Vujić M., Dedov I. First record of the genus Xeropicta Monterosato, 1892 (Gastropoda: Eupulmonata: Geomitridae) in Serbia // Folia Malacologica. First record of the genus Xeropicta Monterosato, 1892 (Gastropoda). 2022. Vol. 30, № 1. P. 47−53.
Hausdorf B., Parr M., Shappell L. J., Oldeland J., Robinson D. G. The introduction of the European Caucasotachea vindobonensis (Gastropoda: Helicidae) in North America, its origin and its potential range // Biological Invasions. The introduction of the European Caucasotachea vindobonensis (Gastropoda). 2021. Vol. 23, № 11. P. 3281−3289.
Kajtoch Ł., Davison A., Grindon A., Deli T., Sramkó G., Gwardjan M., Kramarenko S., Mierzwa-Szymkowiak D., Ruta R., Ścibior R., Tóth J. P., Wade C., Kolasa M., Egorov R. V., Fehér Z. Reconstructed historical distribution and phylogeography unravels non-steppic origin of Caucasotachea vindobonensis (Gastropoda: Helicidae) // Organisms Diversity & Evolution. Reconstructed historical distribution and phylogeography unravels non-steppic origin of Caucasotachea vindobonensis (Gastropoda). 2017. Vol. 17, № 3. P. 679−692.
Kiss L., Labaune C., Magnin F., Aubry S. Plasticity of the life cycle of Xeropicta derbentina (Krynicki, 1836), a recently introduced snail in mediterranean France // Journal of Molluscan Studies. 2005. Vol. 71, № 3. P. 221−231.
Mierzwa D. Cepaea vindobonensis (Férussac, 1821) (Gastropoda: Pulmonata: Helicidae) in central, northwestern and western Poland // Folia Malacologica. 2009. Vol. 17, № 4. P. 185−198.
Olden J. D., LeRoy Poff N., Douglas M. R., Douglas M. E., Fausch K. D. Ecological and evolutionary consequences of biotic homogenization // Trends in Ecology & Evolution. 2004. Vol. 19, № 1. P. 18−24.
Ostrovsky A. M. Xeropicta derbentina (Krynicki, 1836) (Gastropoda: Eupulmonata: Geomitridae) in Belarus – new data // Folia Malacologica. 2023. Vol. 31, № 1. P. 43−47. DOI: 10.12657/folmal.031.006.
R Core Team (2023). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL: https://www.R-project.org/ (дата обращения: 22.12.2023).
Sakai A. K., Allendorf F. W., Holt J. S., Lodge D. M., Molofsky J., With K. A., Baughman S., Cabin R. J., Cohen J. E., Ellstrand N. C., McCauley D. E., O’Neil P., Parker I. M., Thompson J. N., Weller S. G. The population biology of invasive species // Annual Review of Ecology and Systematics. 2001. Vol. 32, № 1. P. 305−332.
Simberloff D., Martin J. L., Genovesi P., Maris V., Wardle D. A., Aronson J., Courchamp F., Galil B., García-Berthou E., Pascal M., Pyšek P., Sousa R., Tabacchi E., Vilà M. Impacts of biological invasions: what’s what and the way forward // Trends in Ecology & Evolution. 2013. Vol. 28, № 1. P. 58−66.
Staikou A. E. Aspects of life cycle, population dynamics, growth and secondary production of the pulmonate snail Cepaea vindobonensis (Férussac, 1821) in northern Greece // Journal of Molluscan Studies. 1998. Vol. 64, № 3. P. 297−308.
Wagner A., Bertrand A. Première observation de l’Hélicelle des Balkans, Xeropicta derbentina (Krynicki, 1836) (Mollusca, Gastropoda, Geomitridae) en Alsace (France, Grand Est, Bas-Rhin) // Folia Conchyliologica. 2021. Vol. 66. P. 73−76.
Williamson P., Cameron R., Carter M. Population dynamics of the landsnail Cepaea nemoralis L.: А six-year study // The Journal of Animal Ecology. 1977. P. 181−194.

 © 2011 - 2024
© 2011 - 2024