



Введение
Ассоциированная со льдом фауна, к которой относят ряд представителей ракообразных, является неотъемлемой частью морских сообществ арктическиx и антарктических вод (Мельников, 1989; Gulliksen, Lønne, 1991; Garrison, 1991; Werner, Arbizu, 1999; Arndt, Swadling, 2006). Входящие в данные сообщества виды подразделяются на автохтонные, весь жизненный цикл которых связан со льдом, и аллохтонные, ассоциированные с ним лишь в определенные периоды жизненного цикла (Horner et al., 1992). К автохтонным представителям арктической подледной фауны относятся амфиподы Gammarus wilkitski Birula, 1897, Onisimus glacialis G. O. Sars, 1900, O. nanseni G. O. Sars, 1900 и Apherusa glacialis (Hansen, 1888), мизида Mysis polaris Holmquist, 1959 и копепода Jaschnovia brevis (Gulliksen, Lønne, 1991). Однако принадлежность последнего вида к ледовой фауне вызывает ряд сомнений ввиду отсутствия наблюдений всех стадий жизненного цикла вблизи ледовой поверхности.
Изначальной целью нашей работы являлось изучение особенностей распределения автохтонной ледовой фауны амфипод в районе границы распространения ледового покрова в северной части Баренцева моря в зависимости от сезонных особенностей ледовых условий. Однако, после обнаружения в отобранном материале особей J. brevis, фокус работы закономерно сместился на данный вид и описание особенностей его распределения в новом месте обитания, а также пересмотр имеющихся в литературе представлений относительно распространения вида J. brevis с целью уточнения его экологических особенностей.
Материалы
Отбор проб зоопланктона был произведен в ходе двух экспедиций ММБИ КНЦ РАН на НИС «Дальние Зеленцы» в июле и ноябре 2017 г. (рис. 1, табл. 1).
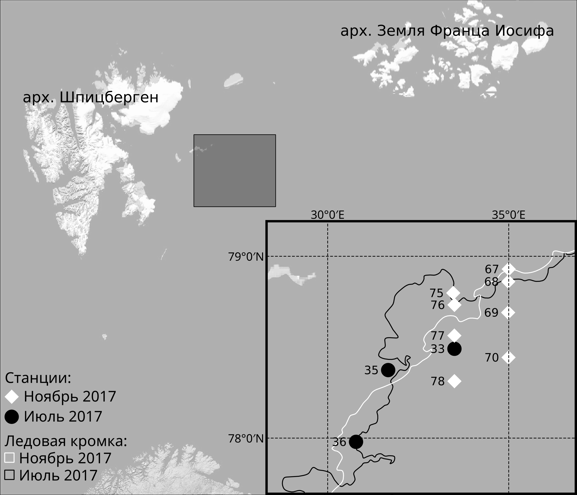
Рис. 1. Схема расположения станций и границы распространения льдов
Fig. 1. Scheme of location of stations and boundaries of ice
Зоопланктон отбирали двумя методами (путем горизонтального и вертикального облова столба воды, см. табл. 1) с использованием сетей ИКС-80 (диаметр входного отверстия 80 см, размер ячеи 500 мкм) и WP-2 (диаметр входного отверстия 50 см, размер ячеи 200 мкм). Вертикальные ловы в июле осуществлялись от дна до поверхности, в ноябре – в слое 0–50 м. Горизонтальные ловы выполнялись в течение 10 минут с момента касания входного обруча поверхности до полного его выхода из воды. В течение этого времени судно циркулировало с постоянной скоростью, равной 3 узлам. Максимальный слой облова не превышал 5–10 метров. На станциях 67 и 75 (см. рис. 1), ввиду присутствия у поверхности большого количества мелких ледовых образований, горизонтальное траление не проводилось. Всего в ходе экспедиционных работ было отобрано 20 проб на 11 станциях.
Таблица 1. Характеристики станций отбора проб
| Станция | Дата | Широта | Долгота | Орудие лова | Горизонтальный лов | Вертикальный лов |
| 33 | 16.07.2017 | 78.5020 | 33.5013 | ИКС-80 | + | + |
| 35 | 17.07.2017 | 78.3843 | 31.6710 | ИКС-80 | + | + |
| 36 | 17.07.2017 | 77.9786 | 30.7827 | ИКС-80 | + | + |
| 67 | 28.11.2017 | 78.9322 | 35.0000 | WP-2 | - | + |
| 68 | 28.11.2017 | 78.8657 | 34.9877 | ИКС-80, WP-2 | + | + |
| 69 | 28.11.2017 | 78.6998 | 34.9847 | ИКС-80 | + | - |
| 70 | 28.11.2017 | 78.4542 | 34.9953 | ИКС-80 | + | - |
| 75 | 29.11.2017 | 78.8050 | 33.4683 | ИКС-80, WP-2 | - | + |
| 76 | 29.11.2017 | 78.7410 | 33.5012 | ИКС-80, WP-2 | + | + |
| 77 | 29.11.2017 | 78.5743 | 33.4997 | ИКС-80 | + | - |
| 78 | 29.11.2017 | 78.3225 | 33.5007 | ИКС-80 | + | - |
Абиотические характеристики среды на станциях, такие как соленость и температура, измерялись при помощи CTD зонда SBE 19 plus V2 и SBE 19 plus фирмы SEA-BIRD ELECTRONICS (США).
Собранный материал фиксировали 4 %-ным раствором формалина. При дальнейшей камеральной обработке каждую пробу просматривали тотально в камере Богорова с использованием микроскопа МБС–10.
Информация о границах распространения льдов на исследованной акватории (см. рис. 1), о типе льда и его сплоченности была взята на сайте ФГБУ «ААНИИ» (www.aari.ru).
Результаты
Характеристика условий среды
Визуальные наблюдения с борта судна, а также спутниковые данные о положении границы распространения льда во время проведения экспедиционных работ свидетельствуют о том, что ледовая обстановка в районе исследований в июле значительно отличалась от таковой в ноябре. Граница льда была достаточно хорошо видна невооруженным глазом. Сплоченность льда в непосредственной близости от района исследований составляла 7–10 баллов. С другой стороны, в конце ноября ледовый покров только начал формироваться (рис. 1) и был представлен ниласом и молодым льдом низкой сплоченности. Визуально наблюдались отдельные небольшие ледовые поля.
Значения солености и температуры на станциях в пределах каждого сезона отличались незначительно. Характер их изменения на двух станциях в июле и ноябре в зависимости от глубины представлен на рис. 2. Средние значения температуры и солености в поверхностном 10-метровом слое на станциях приведены в табл. 2. Особое внимание с нашей стороны к данному слою вызвано тем, что именно в нем в июле особи J. brevis отсутствовали на всех станциях, в то время как в ноябре наблюдалась противоположная картина (за исключением ст. 78).
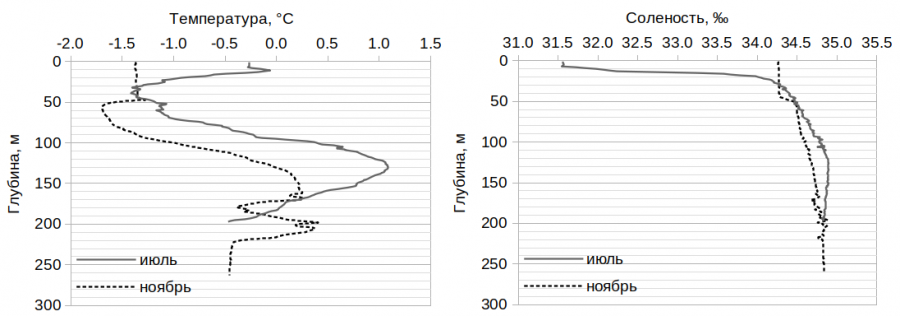
Рис. 2. Распределение показателей температуры и солености в зависимости от глубины на ст. 35 в июле и ст. 77 в ноябре
Fig. 2. Distribution of temperature and salinity indications depending on depth at st. 35 in July and st. 77 in November
Таблица 2. Значения температуры и солености в поверхностном слое 0–10 м
| Июль | |||||||
| Станция | Температура, °С | Соленость, ‰ | |||||
| Среднее | Мин. | Макс. | Среднее | Мин. | Макс. | ||
| 33 | -0.01 | -0.23 | 0.05 | 32.42 | 32.35 | 32.6 | |
| 35 | -0.24 | -0.28 | -0.11 | 31.65 | 31.55 | 31.98 | |
| 36 | -0.56 | -0.69 | -0.38 | 31.84 | 31.45 | 32.51 | |
| Ноябрь | |||||||
| Станция | Температура, °С | Соленость, ‰ | |||||
| Среднее | Мин. | Макс. | Среднее | Мин. | Макс. | ||
| 67 | -1.69 | -1.69 | -1.68 | 34.04 | 34.04 | 34.05 | |
| 68 | н/д | н/д | |||||
| 69 | -1.45 | -1.45 | -1.45 | 34.09 | 34.09 | 34.09 | |
| 70 | -0.97 | -0.97 | -0.97 | 33.96 | 33.96 | 33.96 | |
| 75 | -1.78 | -1.8 | -1.74 | 34.1 | 34.1 | 34.11 | |
| 76 | -1.39 | -1.39 | -1.38 | 34.22 | 34.21 | 34.22 | |
| 77 | -1.38 | -1.38 | -1.37 | 34.26 | 34.26 | 34.27 | |
| 78 | -1.48 | -1.48 | -1.47 | 34.07 | 34.07 | 34.07 | |
Jaschnovia brevis и представители ледовой фауны
В июле на исследованной акватории были обнаружены 3 вида амфипод (A. glacialis, O. glacialis и G. wilkitski) и 1 вид копепод (J. brevis), относящихся к представителям ледовой фауны (Gulliksen, Lønne, 1991). Все эти виды присутствовали на наиболее близких к ледовой кромке станциях, в то время как на удалении от нее (ст. 33) были обнаружены только особи A. glacialis и J. brevis. Следует отметить, что последний вид был зарегистрирован на всех станциях только в вертикальных ловах от дна до поверхности. Глубина в районе проведения работ составляла 220–235 м. Популяция J. brevis была представлена II–V копеподитными стадиями развития.
В конце ноября картина пространственного распределения J. brevis выглядела сходным образом, за исключением того, что особи вида присутствовали как в поверхностном слое (горизонтальные ловы), так и в нижележащем. Лишь на одной станции – ст. 78 (самая удаленная от границы распространения льда, наравне со ст. 70) не было обнаружено ни одной особи. Возрастная структура популяции J. brevis, в отличие от таковой в июле, характеризовалась наличием только старших копеподитных стадий V–VI, причем половозрелые особи были представлены как самками, так и самцами.
Обнаруженные в июле амфиподы практически полностью отсутствовали в конце ноября, за исключением одной особи G. wilkitski, обнаруженной в ловах на ст. 69.
Обсуждение
Несмотря на то, что особи J. brevis отмечались ранее почти во всех морях российского сектора Арктики, нахождение этого вида на исследованной нами акватории отмечается впервые. Предположение о присутствии J. brevis в Баренцевом море высказывалось ранее (Scott et al., 2002), но точного документального подтверждения этому до сих пор получено не было. Ранее на акватории Стур-фьорда были отмечены представители рода Jaschnovia (Hirche, Kosobokova, 2011), которые, однако, в равной степени могли являться как видом J. brevis, так и J. tolli (Linko, 1913). В видовых списках зоопланктона Баренцева моря упоминания J. brevis отсутствуют (Dvoretsky, Dvoretsky, 2010). Систематическое описание вида (Markhaseva, 1996) не включает вышеуказанную акваторию в географические границы его местообитания. Однако, если предположить, что J. brevis является ассоциированным со льдом видом, и учесть, что в экстремально холодные годы ледовитость Баренцева моря может достигать 86 % от его общей площади (Воды Баренцева…, 2016), то логично заключить, что J. brevis в отдельные периоды может иметь широкое распространение, которое не ограничивается лишь северной частью акватории.
Полученный нами материал, а также накопленные к настоящему времени сведения относительно распространения популяции J. brevis в различных биотопах позволяют при детальном рассмотрении дать более точную экологическую характеристику данному виду.
Несмотря на многочисленность свидетельств обитания J. brevis в непосредственной близости от нижней поверхности ледового покрова, что служит основанием для причисления ее к автохтонной ледовой фауне (Melnikov, Kulikov, 1980; Werner et al., 2002; Scott et al., 2002; Ikävalko, 2004; Kosobokova et al., 2011), можно привести столько же, если не больше, примеров обнаружения особей этого вида на акваториях, полностью свободных ото льда (Kosobokova et al., 1998; Fetzer et al., 2002; Walkusz et al., 2010, 2013; Ershova et al., 2015).
В дополнение к этому следует отметить и различия в распространении типичных представителей автохтонной ледовой фауны, таких как G. wilkitski, O. glacialis и A. glacialis в прикромочной зоне, обнаруженные нами как в период таяния льда в июле, так и в момент его образования в ноябре. Будучи достаточно жестко привязанными ко льду как субстрату, G. wilkitski и O. glacialis в июле предсказуемо были обнаружены только на близких к ледовой кромке станциях. Наиболее мобильной из трех видов ледовых амфипод является A. glacialis, чем и объясняется ее присутствие на большем расстоянии от ледовой кромки. В конце ноября, когда еще не существует непосредственного контакта только начинающего формироваться ледового покрова с многолетними льдами – центром расселения данных видов (Gulliksen, Lønne, 1991; Hop, Pavlova, 2008), полное отсутствие этих «ледовых» амфипод в нашем материале также является вполне закономерным. При этом J. brevis присутствовала на всей исследованной акватории как в июле, так и в ноябре (исключение – ст. 78).
Вместе с тем состав липидов у J. brevis сходен с таковым у O. glacialis, A. glacialis и G. wilkitski (Scott et al., 1999, 2002; Werner, Auel, 2005), что свидетельствует о сходстве их рационов. Отличительной чертой липидов всех этих видов является преобладание триацилглицеролов над восковыми эфирами, что характерно как для арктической, так и антарктической ледовой фауны ракообразных (Lee et al., 2006). Однако в работе Скотт с соавторами (Scott et al., 2002) указывается, что у J. brevis значительная доля жирных кислот с короткими цепями (16:0n-7, 16:0, 14:0) сопоставима с таковой у Metridia longa (Lubbock, 1854), являющейся типичным арктическим интерзональным всеядным видом (Кособокова, 2012).
Можно предположить, что другой важной абиотической характеристикой, возможно, ограничивающей распространение J. brevis, является соленость. Однако при внимательном рассмотрении имеющихся данных становится ясно, что данный фактор вряд ли может считаться лимитирующим ввиду очень широкого диапазона его значений, при которых данный вид был обнаружен. Так, в Арктическом бассейне J. brevis обитала в районах с соленостью 29.6–31 ‰ (Мельников, 1989), в Чукотском море – 31–33 ‰ (Ершова, 2017), в море Лаптевых – 17–33 ‰ (Kosobokova et al., 1998), в Карском море – 2–33 ‰ (Fetzer et al., 2002), в Баренцевом море – 32–34.9 ‰ (наши данные), в Гренландском море – около 30 ‰ (Werner et al., 2002), в море Бофорта – 31.6–32.7 ‰ (Walkusz et al., 2013).
Из вышесказанного следует, что принадлежность J. brevis к автохтонной ледовой фауне, а также ее приуроченность к определенным соленостным характеристикам неоднозначна. В настоящее время в литературе присутствует много аргументов как за, так и против использования этих параметров в качестве определяющих экологические предпочтения данного вида. В значительно большей степени, по нашему мнению, на данную роль подходят такие характеристики, как температура и слой обитания, с помощью которых можно достаточно логично объяснить все несоответствия в экологическом описании вида.
Так, с большой долей вероятности можно говорить, что J. brevis является эпипелагическим видом, ограниченным в своем распространении изобатой 50 м. Обнаружение особей у дна в Гренландском море (Markhaseva, 1996) также укладывается в это утверждение, поскольку глубина в районах ее находок составляла 17 и 50 м, а обнаружение всех копеподитных стадий в поверхностных слоях в глубоководной Арктике (Мельников, 1989; наши данные) исключает жесткую связь жизненного цикла с придонным слоем. J. brevis обитает на акватории большинства шельфовых арктических морей, где глубина в основном не превышает 50 м, а в более глубоководных районах, таких как Арктический бассейн, обнаруживается лишь в поверхностном слое (Мельников, 1989).
Лучшее объяснение картины пространственного распространения J. brevis можно получить, если допустить, что температурный диапазон обитания данного вида включает значения от -1° С и ниже. В пользу этого свидетельствуют и наши данные. Особи вида отсутствовали на всех трех станциях в поверхностном 10-метровом слое в июле, где температура не опускалась ниже -0.6 °С (см. табл. 2). Несмотря на небольшое количество станций, данный факт можно считать достоверным, поскольку объем профильтрованной воды при горизонтальных ловах на каждой станции составлял более 400 м3, а расстояние траления – около 900 м. Сходная картина наблюдалась в море Бофорта, где особи J. brevis присутствовали в ловах из слоя со средней температурой около -1.3 °С и отсутствовали у поверхности, где температура была существенно выше и достигала значений от 3 до 8.5 ºС ((Walkusz et al., 2013). Несмотря на то, что в большинстве рассмотренных нами работ (Мельников, 1989; Markhaseva, 1996; Kosobokova et al., 1998; Fetzer et al., 2002; Werner et al., 2002; Walkusz et al., 2013; Ершова, 2017) зоопланктонные ловы осуществлялись тотально, во всех случаях обнаружения J. brevis в водной толще отмечались на станциях, где в пределах слоя 0–50 м регистрировалась температура ниже -1 °С.
Заключение
Обобщая вышесказанное, можно предположить, что J. brevis скорее является аллохтонным видом ледовой фауны, связанным не со льдом как таковым, а со специфическими температурными условиями, характерными для периода образования/существования ледового покрова, в отсутствие которых температура поверхностного слоя повышается, и данный вид переходит в эпипелагиаль в слои с диапазоном температур от -1 °С до близких к температуре замерзания значений (-1.7 °С).
В заключение хотелось бы отметить, что наши выводы, несмотря на некоторую категоричность высказываний, носят дискуссионный характер. Возможно, по мере накопления новых данных окажется, что жизненный цикл J. brevis зависит от наличия субстрата в большей степени, чем это представляется на данном этапе. Например, для прикрепления яиц, что характерно для некоторых представителей семейства Aetideidae (Kosobokova et al., 2007).
Библиография
Воды Баренцева моря: структура, циркуляция, изменчивость / В. К. Ожигин, В. А. Ившин, А. Г. Трофимов, А. Л. Карсаков, М. Ю. Анциферов; ПИНРО. Мурманск: ПИНРО, 2016. 260 с.
Ершова Е. А. Структура и межгодовая изменчивость зоопланктонных сообществ Чукотского моря : Дис. ... канд. биол. наук. М., 2017. 128 с.
Кособокова К. Н. Зоопланктон Арктического бассейна // Структура сообществ, экология, закономерности распределения. М.: ГЕОС, 2012. 271 с.
Мельников И. А. Экосистема арктического морского льда . М.: ИО АН СССР, 1989. 191 с.
Arndt C. E., Swadling K. M. Crustacea in Arctic and Antarctic sea ice: distribution, diet and life history strategies // Advances in marine biology. 2006. Vol. 51. P. 197–315.
Dvoretsky V. G., Dvoretsky A. G. Checklist of fauna found in zooplankton samples from the Barents Sea // Polar biology. 2010. Vol. 33. № 7. P. 991–1005.
Ershova E. A., Hopcroft R. R., Kosobokova K. N. Inter-annual variability of summer mesozooplankton communities of the western Chukchi Sea: 2004–2012 // Polar Biology. 2015. Vol. 38. № 9. P. 1461–1481.
Fetzer I., Hirche H., Kolosova E. The influence of freshwater discharge on the distribution of zooplankton in the southern Kara Sea // Polar Biology. 2002. Vol. 25. № 6. P. 404–415.
Garrison D. L. Antarctic sea ice biota // American Zoologist. 1991. Vol. 31. № 1. P. 17–34.
Gulliksen B., Lønne O. J. Sea ice macrofauna in the Antarctic and the Arctic // Journal of Marine Systems. 1991. Vol. 2. № 1–2. P. 53–61.
Hirche H. J., Kosobokova K. N. Winter studies on zooplankton in Arctic seas: the Storfjord (Svalbard) and adjacent ice-covered Barents Sea // Marine biology. 2011. Vol. 158. № 10. С. 2359–2376.
Hop H., Pavlova O. Distribution and biomass transport of ice amphipods in drifting sea ice around Svalbard // Deep Sea Research Part II: Topical Studies in Oceanography. 2008. Vol. 55. № 20–21. P. 2292–2307.
Horner R., Ackley S. F., Dieckmann G. S. et al. Ecology of sea ice biota // Polar Biology. 1992. Vol. 12. № 3–4. P. 417–427.
Ikävalko J. Checklist of unicellular and invertebrate organisms within and closely associated with sea ice in the Arctic regions. Merentutkimuslaitos, 2004.
Kosobokova K. N., Hanssen H., Hirche H. J. et al. Composition and distribution of zooplankton in the Laptev Sea and adjacent Nansen Basin during summer, 1993 // Polar Biology. 1998. Vol. 19. № 1. P. 63–76.
Kosobokova K. N., Hirche H. J., Hopcroft R. R. Reproductive biology of deep-water calanoid copepods from the Arctic Ocean // Marine Biology. 2007. Vol. 151. № 3. P. 919–934.
Kosobokova K. N., Hopcroft R. R., Hirche H. J. Patterns of zooplankton diversity through the depths of the Arctic’s central basins // Marine Biodiversity. 2011. Vol. 41. № 1. С. 29–50.
Lee R. F., Hagen W., Kattner G. Lipid storage in marine zooplankton // Marine Ecology Progress Series. 2006. Vol. 307. P. 273–306.
Markhaseva E. L. Calanoid copepods of the family Aetideidae of the world ocean // Trudy Zoologicheskogo Instituta. 1996. Vol. 268.
Melnikov I. A., Kulikov A. S. The cryopelagic fauna of the central Arctic Basin // Biology of the central Arctic Basin. Moscow, 1980. P. 97–111.
Scott C. L., Falk-Petersen S., Sargent J. R. et al. Lipids and trophic interactions of ice fauna and pelagic zooplankton in the marginal ice zone of the Barents Sea // Polar Biology. 1999. Vol. 21. № 2. P. 65–70.
Scott C. L., Kwasniewski S., Falk-Petersen S. et al. Lipids and fatty acids in the copepod Jaschnovia brevis (Jaschnov) and in particulates from Arctic waters // Polar Biology. 2002. Т. 25. № 1. С. 65–71.
Walkusz W., Paulić J. E., Kwaśniewski S. et al. Distribution, diversity and biomass of summer zooplankton from the coastal Canadian Beaufort Sea // Polar Biology. 2010. Vol. 33. № 3. P. 321–335.
Walkusz W., Williams W. J., Kwasniewski S. Vertical distribution of mesozooplankton in the coastal Canadian Beaufort Sea in summer // Journal of Marine Systems. 2013. Vol. 127. P. 26–35.
Werner I., Arbizu P. M. The sub-ice fauna of the Laptev Sea and the adjacent Arctic Ocean in summer 1995 // Polar Biology. 1999. Vol. 21. № 2. P. 71–79.
Werner I., Meiners K., Schünemann H. Copepods in Arctic pack ice and the underlying water column: living conditions and exchange processes // Ice in the Environment: Proceedings of the 16th IAHR International Symposium on Ice. 2002. Vol. 3. P. 30–40.
Werner I., Auel H. Seasonal variability in abundance, respiration and lipid composition of Arctic under-ice amphipods // Marine Ecology Progress Series. 2005. Vol. 292. P. 251–262.
Благодарности
Благодарим рецензентов за ценные замечания, высказанные в процессе подготовки рукописи к публикации.
Исследование выполнено за счет гранта Российского научного фонда (проект № 17-14-01268).

 © 2011 - 2026
© 2011 - 2026