



Введение
Вопрос, формируются ли экологические сообщества в соответствии с определенными правилами или же под действием случайных причин, является одним из основных в современной экологии. Считается, что сообщества птиц являются рыхлыми собраниями видов (loose assemblages), которые в состоянии сосуществовать с разными видами (Sybertz, Reich, 2015). Вместе с тем для объяснения неслучайных группировок видов был сформулирован ряд правил. Наиболее спорной оказалась идея межвидовой конкуренции. Diamond (1975) предполагал, что межвидовая конкуренция может порождать запрещенные комбинации видов, которые никогда не встречаются в природе, и распределения, соответствующие шахматной доске, при которых конкурирующие виды избегают друг друга. Кроме того, теория межвидовой конкуренции предполагает, что сходные виды для совместного сосуществования должны различаться размерами тела, что позволяет уменьшить конкуренцию за потребляемый ресурс (Wang et al., 2011). Fox в 1987 г. (цит. по: Wang et al., 2011) в рамках теории межвидовой конкуренции высказал идею о том, что природные сообщества формируются за счет включения в их состав видов из разных функциональных групп. В отличие от предыдущих моделей, вложенные структуры (nestedness) не связаны с межвидовой конкуренцией и возникают в случае, когда бедные видами участки являются подмножествами богатых видами участков (Patterson, Atmar, 1986). Кроме того, возможны и индивидуалистические ответы видов на местные градиенты факторов среды, которые также могут приводить к формированию определенных пространственных структур.
Для объяснения наблюдаемой территориальной неоднородности видового состава, структуры и динамики сообществ, а также для проверки ранее высказанных гипотез и правил сформулирована синтетическая теория метасообщества (Metacommunity), позволяющая увязать градиенты среды, межвидовые взаимоотношения и случайные процессы. В рамках концепции метасообщества (Metacommunity) к настоящему времени сложились два основных направления изучения пространственных изменений в видовом составе (Presley et al., 2010): первый предполагает выявление механизмов и процессов, вызывающих эти изменения; второй сосредоточен на идентификации образцов распределения видов в пространстве (относительно градиентов среды) и их сравнении с идеализированными образцами.
В последние годы были разработаны аналитические методы, позволяющие выявить многочисленные аспекты структуры метасообщества (Presley et al., 2009). Одним из таких подходов является анализ элементов структуры метасообщества (Analysis of elements of metacommunity structure – EMS), позволяющий выявить когерентность, сменяемость видового состава вдоль экологического градиента и характер группировки границ видовых диапазонов (ranges). Этот подход позволяет одновременно сравнить наблюдаемое распределение видов с 6 идеализированными образцами, такими как случайное распределение, шахматные доски, вложенные подмножества, равномерный градиент, клементсовский градиент, глисоновский градиент (Leibold, Mikkelson, 2002). Каждый из этих образцов представляет собой участки в многомерном континууме возможных пространственных распределений видов. К настоящему времени выделены также дополнительные образцы – квази-структуры, и модели группировки границ видовых диапазонов (boundary clumping) для вложенных подмножеств, позволяющих еще более детализировать континуум возможных пространственных распределений видов вдоль основных градиентов среды (Presley et al., 2009; Presley et al., 2010). Кроме того, анализ элементов структуры метасообщества (EMS) может быть использован как первый шаг для изучения структуры метасообщества, позволяющий сформулировать ряд гипотез для организации более детальных исследований и выявления процессов, обеспечивающих наблюдаемое распределение видов по местообитаниям. В контексте EMS метасообщество понимается как совокупность экологических сообществ на разных участках, которые потенциально, но не обязательно, могут быть связаны за счет расселения видов, в то время как сообщество – это группа видов в данном месте (Leibold, Mikkelson, 2002).
К настоящему времени сформулированы четыре парадигмы для объяснения структуры метасообщества: 1) нейтральная теория предполагает, что различные виды эквивалентны, и уделяет большое внимание случайным процессам и расселению видов; 2) концепция динамики патчей (Patch dynamics) предполагает, что все участки идентичны, и делает акцент на конкурентной колонизации патчей-участков видами; 3) в рамках концепции сортировки видов (Species sorting) считается, что распределение видов и видовой состав сообществ определяются местными градиентами окружающей среды, а хорошо расселяющиеся виды способны достигать всех подходящих участков; 4) идея массовых эффектов предполагает, что важную роль в формировании сообществ играют экологические градиенты и расселение видов (Meynard, Quinn, 2008). Winegardner с соавторами (Vilmi et al., 2016) предположили, что массовые эффекты и динамика патчей являются частными случаями сортировки видов. С учетом имеющихся публикаций можно предполагать, что соотношение и значимость для структуры сообществ нейтральных процессов и процессов, связанных с экологическими нишами видов, могут различаться в разных физико-географических регионах. Так, например, для сообществ птиц Франции характерно совместное влияние факторов окружающей среды и процессов расселения, тогда как для сообществ Северной Америки решающее значение имеют местные экологические факторы при умеренном влиянии дисперсии (Ozkan et al., 2012). Для лесов на юго-западе Черного моря (Турция) показана доминирующая роль экологической сортировки видов за счет нишевых процессов, тогда как неэкологическим пространственным процессам отводится второстепенная роль (Ozkan et al., 2012).
В рамках данного исследования оценены закономерности распределения видов вдоль важнейших экологических градиентов и возможная роль ряда правил и процессов в формировании сообществ птиц. За счет тестирования нескольких альтернативных моделей сборки сообществ можно выявить ведущие процессы, определяющие наблюдаемую пространственную структуру.
Материалы
Изучение сообществ птиц проводилось на границе южной тайги и северной лесостепи Зауралья (рис. 1). Большая часть сведений получена во время исследований на территории Каменского района Свердловской и Кунашакского района Челябинской областей. Физико-географическое районирование региона принято согласно представлениям Горчаковского (1968), который выделял переходную подзону предлесостепных сосновых и березовых лесов, относящуюся к тайге. Рельеф местности – это слабоволнистая равнина (пенеплен), наклоненная к востоку. Долины местных рек не велики. Отличительная особенность региона – обилие озер со значительными тростниковыми зарослями по берегам. Вегетационный период начинается с конца апреля и продолжается до начала октября. Зима длится 5–5.5 месяца (Орлова, 1962). На зиму все реки и озера замерзают. В западной части района исследований произрастают сосновые леса (Pinus sylvestris). К юго-востоку от Двуреченска сосновые леса сохраняются только по долинам рек, а междуречья заняты мозаичными осиново-березовыми лесами. По поймам рек распространены ивовые и ольховые заросли.
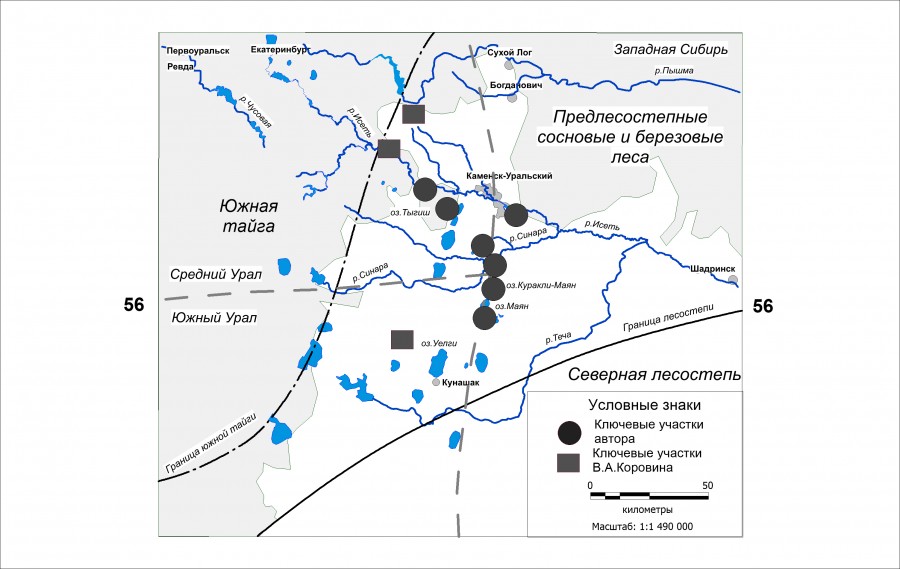
Рис. 1. Расположение ключевых участков на границе тайги и лесостепи Зауралья
Fig. 1. The location of key sites on the border of taiga and forest steppe of Trans-Urals
За последние 300 лет территория сильно преобразована деятельностью человека: луга распаханы и заняты полями, леса и перелески вырублены. В некоторых местах сосновые леса замещены вторичными березняками порослевого или смешанного происхождения. Помимо физического уничтожения коренных ландшафтов, велико промышленное загрязнение территории. На участке Каменск-Уральский – Челябинск располагается головная часть Восточно-Уральского радиоактивного следа. Ключевые участки для полевых исследований были заложены в пределах наименее измененных ландшафтов.
Расположение ключевых участков, маршрутов, площадок. Изучение птиц проводилось в период с 2000 по 2011 г. Суммарная трудоемкость составляет более 330 дней, проведенных «в поле». Для изучения птиц заложена сеть маршрутов, точечных учетов и площадок. Общая протяженность маршрутов (помимо учетов на площадках) более 1000 км. За время исследований найдено около 1000 гнезд птиц.
Первоначально (2000–2005 гг.) в районе исследований была заложена сеть маршрутов, охватывающая основные ландшафты. Маршрутные исследования проводились преимущественно в гнездовой период в долине р. Исеть от д. Шилова до устья р. Каменка, а также от г. Каменск-Уральский до д. Черноскутова. Протяженность исследованного участка долины р. Исеть составляет 53 км. Обследовано 6.5 км долины р. Камышенка (Каменский район Свердловской области) от с. Мал. Белоносов до ее устья и долина р. Каменка в черте г. Каменск-Уральский. В долине р. Синара исследования проводились на участке с. Усть-Багаряк – с. Новый Быт. Протяженность обследованного участка долины р. Синара составляет 9 км. В гнездовой период и зимой проводились исследования в окрестностях оз. Тыгиш, где изучались сообщества птиц, гнездящихся в тростниково-осоковых зарослях и в окрестных осиново-березовых перелесках среди лугов-покосов и полей. Дополнительно в гнездовой период были обследованы оз. Большой Сунгуль, Малый Сунгуль и Червяное. Маршрутные исследования проводились в районе оз. Куракли-Маян. Краткосрочные маршрутные исследования в гнездовой период проводились в районе оз. Березовское, Сосновское, Черное, Уелги, Сайгерлы, Айдыкуль, Комкуль, Сугояк, Мамынкуль, Алакуль, Актюбинское, Тимренкуль. Также непродолжительные исследования проводились в районе оз. Шувакиш к северо-западу от г. Екатеринбурга. В черте г. Каменск-Уральский в весеннее и летнее время исследования проводились на территории рекультивированных отвалов алюминиевого завода. В пределах жилой застройки изучение птиц проводилось только в зимнее время (г. Каменск-Уральский, с. Рыбниково). В агроценозах заложен 5-километровый маршрут к востоку от д. Черноусова. Лесополоса вдоль железной дороги изучалась на участке от «Станция 35 км» до оз. Комкуль (железнодорожная ветка Каменск-Уральский – Челябинск).
В 2006–2011 гг. использовались точечные учеты на большой территории на участке к востоку от железной дороги Каменск-Уральский – Челябинск от долины р. Синара (п. Новый Быт) на севере до юго-западного берега оз. Куракли-Маян на юге, в террасных сосняках долины Исети в окрестностях с. Маминское - д. Шилова - с. Исетское, в террасных березняках к востоку от Каменска-Уральского, в мозаичных осиново-березовых лесах междуречий.
Дополнительно в 2005 г., в первой декаде мая, а затем с третьей декады мая по конец июля, проводились стационарные исследования птиц в районе оз. Маян на участке оз. Маян – оз. Комкуль – оз. Алакуль.
В 2006, 2010 и в 2011 гг. в пределах ранее указанного участка к востоку от железной дороги Каменск-Уральский – Челябинск проводилось тотальное картирование гнезд Соколообразных и Совообразных. Ширина обследованного участка от 8 км на севере до 3 км на юге, площадь 40 км2. На междуречье исследования проводились на участке шириной 5 км, к западу от железной дороги Каменск-Уральский – Челябинск от широты с. Сипавское на севере до долины р. Синара и Багаряк на юге. В пределах данного участка дополнительно была заложена учетная площадка для картирования гнезд Соколообразных и Совообразных размером 10 км2.
В 2010–2011 гг. в окрестностях оз. Куракли-Маян проводилось тотальное картирование гнездящихся птиц на слабо лесистой площадке размером 1.7 км2. В 2011 г. дополнительно проводилось тотальное картирование птиц на площадке размером 9 км2 на участке между с. Окулово и оз. Куракли-Маян.
В пределах изученной территории было выбрано 17 участков-сайтов, по которым имеется наибольшее количество данных о гнездящихся видах птиц. Каждый из выделенных сайтов включает несколько биоценозов. Таким образом, выделение участков для описания проводилось по территориальному принципу, а не с учетом однообразия. В этом случае гораздо удобнее использовать внемасштабный термин «сайт», чтобы не путать с учетной площадкой или ключевым участком.
О границах сайтов. Для сайтов, расположенных в пределах обширных и однообразных территорий, в качестве границ были выбраны условные границы обследованной территории. В этом случае сайт – это обследованная площадка в пределах обширного ландшафта. Для сайтов небольшой площади в качестве границ выбраны естественные границы, хорошо видимые на местности. Например, озеро Тыгиш (Каменский район Свердловской области) имеет площадь чуть более 6 км2. Озерная котловина данного озера была выбрана в качестве сайта, включающего акваторию и узкий бордюр из тростниково-осоковых зарослей. Оз. Тыгищ расположено изолированно от других водоемов, его можно обследовать целиком и обойти вокруг. Озерная котловина оз. Куракли-Маян (Кунашакский район Челябинской области) в 4 раза больше, чем котловина Тыгина. Кроме того, по берегам Куракли-Маян произрастают обширные тростниковые, рогозовые и осоковые заросли. Максимальная ширина этих зарослей более 2 км (расстояние от края тростников до открытой воды – непосредственно акватории оз. Куракли-Маян). В пределах обширных тростниковых зарослей оз. Куракли-Маян был выбран участок площадью около 5 км2, в пределах которого и проводились исследования птиц. Обследовать же все оз. Куракли-Маян практически невозможно.
В связи с ограниченностью возможностей исследование каждого выделенного сайта проводилось не ежегодно. В отдельные годы больше внимания уделялось то одному, то другому сайту. Кроме того, богатым видами птиц сайтам, где велика возможность пропуска видов, уделялось в целом больше внимания, а полевые исследования были более продолжительными по сравнению с сайтами, населенными небольшим числом видов птиц.
На междуречьях исследуемые сайты имели форму многоугольников, квадратную или прямоугольную форму. В долинах рек сайты – многоугольники – ориентированы вдоль русла реки. Списки видов для каждого выделенного на местности сайта составлялись путем обобщения всех имеющихся данных, полученных в период с 2000 по 2011 г. При статистической обработке использовались только сайты, по которым имеются данные не менее чем за 3 гнездовых периода. Пересчет на площадь и расчет плотности населения не производились.
Полевые исследования проводились во время гнездового периода (май – июнь) и зимой (преимущественно в 2000–2002 гг.). Кроме того, для выявления полного видового состава птиц в районе исследований и особенностей фенологии местных видов полевые исследования проводились во второй половине апреля – мае и в июле – августе. При статистической обработке данных сведения о пролетных видах исключены из анализа. Для каждого выделенного на местности сайта составлялись списки гнездящихся и возможно гнездящихся видов, пролетных видов, использующих данный сайт только во время сезонных миграций, а также зимующих видов. К гнездящимся относились виды, для которых были получены прямые свидетельства гнездования: находки гнезд с кладкой или птенцами, встречи плохо летающих нераспавшихся выводков, встречи взрослых птиц, таскающих корм птенцам в гнезде или вблизи гнезда, встречи взрослых птиц, сидящих в гнезде в характерной позе (некоторые хищные, чайки и крачки, гнезда которых, расположенные в труднодоступных местах, не были осмотрены). К возможно гнездящимся видам относились виды, для которых не удалось получить прямые доказательства гнездования. В этом случае критериями возможного гнездования являлись встречи беспокоящихся взрослых особей, а также территориальных самцов, поющих в течение долгого времени в пределах небольшого подходящего для гнездования участка. Ряд видов (погоныши, водяной пастушок) считались безусловно гнездящимися, хотя прямых и косвенных признаков гнездования не было найдено.
Учеты птиц проводились как в светлое время суток, так и ночью (за исключением сомкнутых лесов).
Методы
По результатам учетов птиц строились матрицы присутствия-отсутствия, где в строках располагались виды птиц, а в столбцах – заселяемые этими видами сайты. Принадлежность вида к тому или иному сайту оценивалась только по факту гнездования в его пределах. При построении матрицы присутствия-отсутствия данные, полученные при помощи разных методов учетов, но в пределах одного сайта, объединялись. Перед анализом структуры метасообщества все встречи птиц за разные годы в одном сайте также объединялись для того, чтобы минимизировать влияние случайных причин на состав населения птиц (Bert, 2001; Royan et al., 2016). Вид считался присутствующим в пределах какого-либо сайта, если он был обнаружен гнездящимся хотя бы один раз за все время исследований. В данном исследовании сайт – участок земной поверхности размером от 2–3 до 10 км2, поддерживающий существование локального сообщества птиц. Участки меньшего размера не рассматривались, учеты в их пределах не проводились. В пределах одного сайта могут располагаться несколько местообитаний или биоценозов. Термин «сайт» используется вместо понятий «местообитание», «биотоп» или «биоценоз», т. к. сайт является атрибутом местности, а не характеристикой конкретных видов. Таким образом, «сайт» – это удобный внемасштабный термин.
Для описания особенностей распределения видов по сайтам использовались методы, разработанные в рамках концепции метасообщества (Leibold, Mikkelson, 2002). Данный пространственно неявный подход предполагает тестирование распределения видов по сайтам для определения соответствия ряду идеализированных образцов. К настоящему времени (табл. 1, рис. 2) идентифицировано 6 таких идеализированных образцов: 1) вложенные подмножества (Nestedness), 2) шахматные доски (Checkerboard), 3) клементсовский градиент (Clementsian gradient), 4) глисоновский градиент (Gleasonian gradient), 5) равномерное распределение, 6) случайное распределение (Leibold, Mikkelson, 2002), а также ряд квази-структур. Каждый из этих идеализированных образцов представляет собой область в многомерном континууме структуры метасообщества (Presley et al., 2010). Анализ проводится в три этапа (рис. 3). На первом этапе оценивается последовательность видов (coherence), на втором – сменяемость (turnover), на третьем – характер группировки границ в распределении видов в ряду сайтов (boundary clumping). Каждый из этих трех этапов затрагивает отдельный аспект структуры метасообщества (Leibold, Mikkelson, 2002).
Для проведения EMS в данном исследовании использовались два подхода к ординации матриц присутствия-отсутствия. Первый подход предполагает проведение ординации за счет взаимного усреднения (reciprocal averaging), при котором сайты и виды в матрице присутствия-отсутствия сортируются таким образом, что сходные по видовому составу сайты и виды, населяющие одни и те же сайты, оказываются как можно ближе друг к другу. В этом случае метасообщество выстраивается в соответствии с результирующей всех местных градиентов среды (Leibold, Mikkelson, 2002). Второй подход предполагает ординацию матрицы присутствия-отсутствия в соответствии с осями, полученными с помощью метода главных компонент (Presley et al., 2009). Ординация в соответствии с важнейшими осями позволяет оценить различные ответы метасообщества на существующие градиенты факторов среды в районе исследований.
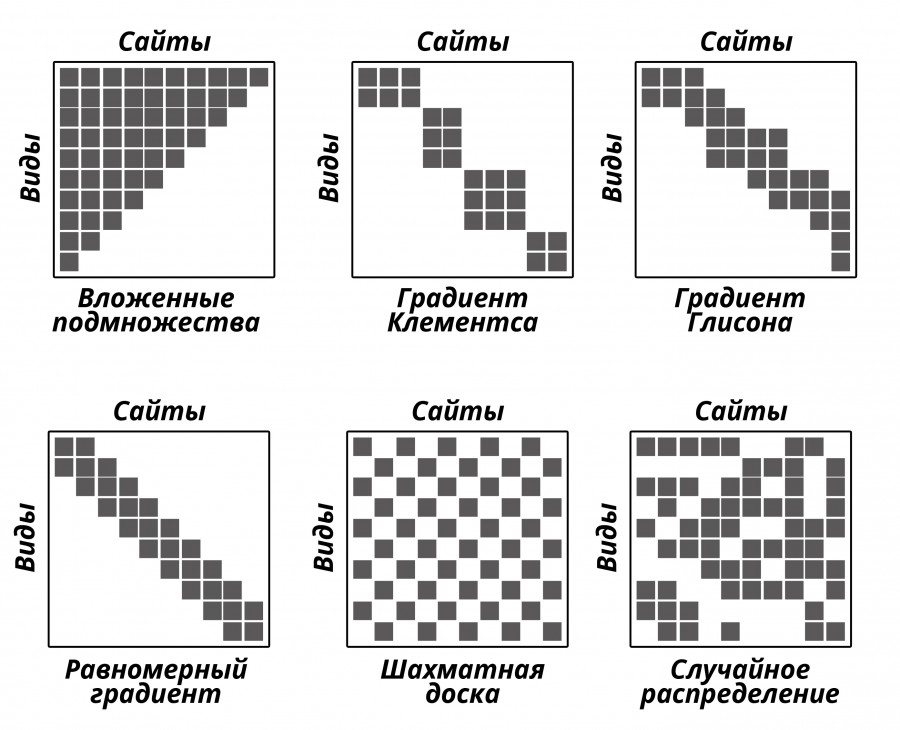
Рис. 2. Основные идеализированные структуры метасообщества. По оси «Х» располагаются сайты, по оси «У» – виды. Серые квадратики – присутствие вида в пределах сайта. Белые участки – отсутствие вида в пределах сайта
Fig. 2. The main idealized structures of the meta-communities. On the “X” axis are sites, on the “Y” axis – species. Gray squares denote the presence of species within the site. White areas – the lack of species within the site
Когерентность (coherence) определяется путем подсчета встроенных в распределение видов отсутствий (Embedded absences). Согласно современным представлениям, численность (или плотность населения) вида соответствует гауссовскому распределению. В этом случае, если вид с определенной численностью встречается в двух точках в пределах какого-либо экологического градиента, то он должен заселять и участки, располагающиеся между этими точками, т. е. имеющие промежуточные значения экологического градиента. Таким образом, не должно быть никаких отсутствий вида в n-мерном гиперобъеме, определяющем фундаментальные ниши видов (Presley et al., 2010). Характер ответов видов на экологический градиент может значительно различаться. В случае, если виды, как группа, не демонстрируют одинаковый ответ на один и тот же градиент, то распределение видов не формирует когерентную структуру. Отсутствие когерентности, однако, еще не означает случайного распределения видов по местообитаниям.
Таблица 1. Идеализированные паттерны метасообщества и используемые индексы для их обнаружения
| Паттерн | Описание | Индекс | Когерентность | Оборот видов | Группировка границ |
| Шахматная доска (Checkerboards) | Для пар видов характерны взаимоисключающие распределения (запрещенные комбинации) | C-Score V-Ratio | Отрицательная | Случайный | Случайная |
| Вложенные подмножества (Nested subsets) | Видовой состав местообитаний соответствует вложенным подмножествам | NODF | Положительная | Отрицательный | Случайная |
| Градиент Клементса (Clementsian) | Сообщества представляют собой дискретные группы, замещающие друг друга как группа видов | Morisita | Положительная | Положительный | Положительная Индекс Мориситы больше 1 |
| Градиент Глисона (Gleasonian) | Случайное распределение диапазонов видов вдоль градиентов среды | Morisita | Положительная | Положительный | Не отличается от случайного |
| Равномерный градиент (Evenly spaced gradients) | Диапазоны видов распределены более равномерно, чем ожидается при случайном распределении | Morisita | Положительная | Положительный | Отрицательная, Индекс Мориситы меньше 1 |
| Случайное распределение (Random) | Не наблюдается никаких градиентов в распределении видов по местообитаниям | Не отличается от случайной | Не отличается от случайного | Не отличается от случайной |
Когерентность может быть положительной, отрицательной и случайной. Низкое число встроенных отсутствий (embedded absences) по сравнению с распределением нулевых моделей свидетельствует о положительной когерентности (positive coherence) в распределении видов по сайтам. Лишь после доказательства когерентности в распределении видов можно переходить к следующим этапам. В случае отрицательной когерентности (negative coherence) тестируемое распределение видов будет больше соответствовать шахматной доске (Checkerboard), а на выбор видами местообитаний будет большое влияние оказывать межвидовая конкуренция и взаимное исключение (Leibold, Mikkelson, 2002; Presley et al., 2010). Если же индекс встроенных отсутствий (Embedded absences) не отличается от ожидаемого для нулевой модели, то распределение видов в пределах метасообщества является случайным, а существующий экологический градиент не оказывает влияния на распределение видов по сайтам (Presley et al., 2010). Индекс (EmbAbs) реализован в программе Turnover (Ulrich, 2012)

Рис. 3. Схема анализа элементов структуры метасообщества с дополнениями на основе имеющихся публикаций. Ссылки на источники литературы в тексте
Fig. 3. Scheme of analysis of the elements of the meta-community structure with additions based on existing publications. References to sources are given in the text
Оборот видов (Turnover). Оборот видов (turnover) оценивается путем подсчета количества раз, когда один вид заменяет другой в двух сайтах. В матрице присутствия-отсутствия каждая такая замена называется шахматной доской и имеет вид:
| 1 | 0 |
| 0 | 1 |
Общее число таких замен является мерой оборота видов (turnover) в исследуемой матрице. В этом случае нужно учитывать тот факт, что число таких замен в матрице чувствительно к положению диапазонов (ranges) и сообществ относительно друг друга, а также и к отсутствиям, включенным в эти диапазоны и сообщества. В данном случае эти замены не связаны с основным направлением изменчивости видового состава в пределах какого-либо градиента среды. Перед началом тестирования матрицы присутствия-отсутствия сортируются определенным образом для того, чтобы минимизировать число включенных отсутствий (embedded absences). Только после сортировки матрицы проводится подсчет числа замен и сравнение этого показателя с аналогичным для нулевых моделей. Оборот видов (turnover) может быть положительным или отрицательным (Leibold, Mikkelson, 2002). В случае отрицательного оборота (negative turnover) структура метасообщества будет больше соответствовать вложенным подмножествам (nested subset). Вложенность (nestedness), в свою очередь, также проверяется с помощью расчета специальных статистических показателей.
Во вложенной модели (Nestedness) видовой состав бедных видами сообществ является подмножеством богатых видами сообществ (Ulrich et al., 2009). Одна из целей такого анализа – это выявление отклоняющихся видов, распределение которых не соответствует направлению изменчивости экологических градиентов (Ulrich et al., 2009). Анализ вложенных матриц показывает, что виды-специалисты, населяющие небольшое число сайтов, взаимодействуют с разными группами видов (как правило, с неродственными видами), тогда как генералисты взаимодействуют преимущественно только друг с другом (т. е. с видами-генералистами).
Вложенность (Nestedness) сообществ была показана для многих таксонов, за исключением водных беспозвоночных. Предполагается, что вложенность сообществ (Nestedness) является сложным феноменом, на который оказывает влияние множество факторов (Boecklen, 1997). Также выявлена функциональная вложенность сообществ (Matthews et al., 2015).
При Nestedness-анализе проводится специальная сортировка матрицы. В результате чего все виды и сайты оказываются отсортированными определенным образом: виды, населяющие максимальное количество сайтов, оказываются в верхних строчках матрицы, а наиболее богатые видами сайты располагаются в левой части матрицы. После такого преобразования и вычисления индексов, характеризующих распределение видов по сайтам, проводится сравнение эмпирической матрицы и эмпирических индексов с аналогичными для случайных матриц и индексов, построенных на основе нулевых моделей. В данном случае так же использовалась наиболее консервативная фиксированная (FF) для строк и столбцов нулевая модель.
В данной публикации в качестве индексов при Nestedness-анализе используется NODF и C-Score. NODF (nestedness metric based on overlap and decreasing fill) – это одна из самых популярных мер, используемых в Nestedness-анализе (Podani, Schmera, 2012). Индекс NODF измеряется в диапазоне от 0 до 100. При значении индекса NODF, равном 100, вложенность и упорядоченность матрицы максимальна. Если значение равно нулю, то вложенности (nestedness) нет. Расчет индекса NODF может быть произведен отдельно как для строк и столбцов, так и для всей матрицы в целом (Morrison, 2013).
Для расчета индексов использовалось специализированное программное обеспечение «NODF. Version 2.0, от 03.09.2012» и Ecosim 7. Расчеты индексов, характеризующих распределение видов по местообитаниям, проводились раздельно для мало трансформированных местообитаний (собственные данные) и для агроландшафта. В последнем случае использовались данные о составе гнездящихся видов птиц В. А. Коровина (2004).
Индекс для идиосинкразических видов и сайтов также основывается на NODF. Кроме того, идиосинкразические виды проявляют широкую экологическую валентность и хорошую способность к расселению по сравнению с видами, формирующими вложенные сообщества (Sebastian-Gonzalez et al., 2010).
Группировка границ видовых диапазонов (Clumping), в свою очередь, тестируется только в случае положительного оборота видов. Группировка границ (clumping) может быть положительной, отрицательной или случайной. В случае положительной группировки границ распределение видов соответствует клементсовскому градиенту (Clementsian gradient), в случае отрицательной группировки границ – равномерному градиенту (Evenly spaced gradient), в случае случайной группировки – глисоновскому градиенту (Gleasonian gradient). В качестве меры, характеризующей распределение границ диапазонов (ranges), используется индекс Мориситы (Leibold, Mikkelson, 2002).
При анализе градиентов факторов среды использовались следующие переменные: площадь сайта; лесистость сайта; проективное покрытие сосны (1 ярус) в пределах сайта; проективное покрытие березы (1 ярус) в пределах сайта; проективное покрытие осины (1 ярус) в пределах сайта; число древесных пород первого яруса; рельеф местности; сельскохозяйственная нагрузка; рекреационная нагрузка; обводненность. Все выделенные переменные ранжировались от «0» – фактор отсутствует до «5» – фактор максимально выражен. При определении важнейших экологических градиентов использовался метод главных компонент. Для выделения главных компонент использовалась программа «Past».
Для оценки влияния расстояния между сайтами на видовой состав птиц вычислялся коэффициент корреляции между сходством видового состава птиц (индекс Чекановского – Съеренсена) двух сайтов и расстоянием между этими сайтами. Значения индексов сходства получены в результате попарного сравнения видового состава данного сайта со всеми остальными исследованными сайтами. Аналогичным образом проводилось попарное вычисление расстояния между данным сайтом и всеми остальными сайтами. Для расчета расстояний между сайтами использовались Яндекс.Карты.
Для проверки гипотезы о том, что отношения размеров тела видов, населяющих данный сайт, образуют определенный постоянный интервал, который позволяет снизить межвидовую конкуренцию, вычислялась разница длины тела. На первоначальном этапе проводится сортировка таблицы, содержащей сведения о видовом составе сайта и длине тела каждого вида (Gotelli, Ellison, 2002). Виды ранжируются по длине тела – от наименьшей до наибольшей в данном сайте. Затем вычисляется разница в длине тела двух соседних по размеру видов. Разница в длине тела двух видов для простоты называется авторами метода сегментом. Расчет длины сегмента повторяется для каждой пары видов. В дальнейшем вычислялась дисперсия длины сегмента как показатель постоянства в соотношениях размеров тела. Затем наблюдаемая дисперсия длины сегмента для каждого исследованного сайта сравнивалась с дисперсией, рассчитанной для 1000 сгенерированных случайных сообществ с помощью нулевой модели. Для расчетов использовалась программа Ecosim 7. Для матрицы, состоящей из n видов, Ecosim 7 использует наименьшие и наибольшие размеры тела в качестве границы интервала. Затем программа имитирует размещение остальных n-2 видов внутри этого интервала для создания случайного распределения (Gotelli, Entsminger, 2001).
Минимальная разница в длине сегментов соответствует гипотезе Хатчинсона о том, что для сосуществования двух видов в одном сообществе необходима хотя бы минимальная разница между этими видами. В случае если виды разделены за счет такого критического минимума, то минимальная разница между размерами тела двух видов будет больше, чем предсказывает нулевая модель. Если же наблюдается сходство в размерах тела двух видов, вызванное экологическими ограничениями, то минимальная разница двух видов будет меньше случайной, полученной с помощью нулевых моделей.
Минимальная разница в размерах проверяет гипотезу о том, что существует критическое минимальное различие между видами, необходимое для сосуществования в пределах одного сайта. Дисперсия минимального различия размеров между видами (дисперсия длины сегмента) позволяет проверить гипотезу о том, что размеры видов равномерно распределены, даже если нет никакого минимального различия между размерами тела. Если же дисперсия близка к нулю, то наблюдается одинаковое различие между размерами видов в сообществе. Если же дисперсия велика, то часть видов очень сходна по размеру, а другая часть резко отличается от первой. Для сообществ, структурированных под действием межвидовой конкуренции, наблюдаемые значения дисперсии должны быть меньше, чем для рассчитанных с помощью нулевой модели, и статистически значимо отличаться от случайных (Wang et al., 2011).
Перечень видов приводится в соответствии с новейшей сводкой (Коблик и др., 2006). Сведения о средних размерах тела птиц (длина тела) взяты из сводок «Птицы Советского Союза» (1951–1954) с учетом подвидов. Базовые статистические расчеты (кластерный анализ, одновыборочный t-критерий, корреляция) там, где это не оговорено отдельно, проводились с помощью SPSS Statistics.
.
Результаты
Экологические сообщества являются результатом взаимодействия между местными и региональными процессами (Leibold et al., 2004). Относительная роль этих процессов – рассредоточение видов из регионального пула видов в соответствии с их экологическими нишами – зависит от местной обстановки и особенностей самих видов, составляющих метасообщество.
В пределах рассматриваемой территории выявлены два важнейших градиента, объясняющих 86.7 % дисперсии экологических условий. Первый экологический градиент связан с лесистостью и обводненностью территории (68.6 % дисперсии), второй (18.2 %) – с открытостью и рекреационной нагрузкой. Первая ось представляет собой переход от заболоченных сайтов к лесным через открытые и слабо залесенные сайты (рис. 4).
В период с 2000 по 2011 г. в пределах постоянных маршрутов и площадок автором зарегистрировано 140 гнездящихся и возможно гнездящихся видов птиц. В агроландшафте гнездится 30 видов (Коровин, 2004). Общее число видов, участвующих в анализе, 142. Наибольшее число видов зарегистрировано в лесолуговых сайтах (от 45 до 96 видов), несколько меньше в лесных сайтах (от 31 до 55) и в тростниково-осоковых зарослях по берегам озер (от 36 до 49 видов). Меньше всего видов населяет обширные луга, лишенные кустарниковой и древесной растительности (16), и рекультивированные отвалы в промзоне (17). Сообщества агроландшафта бедны видами (Коровин, 2004). По полям многолетних трав и пастбищам гнездится от 13 до 26 видов. На полях зерновых и пропашных культур, молодых залежах и пашнях число гнездящихся видов очень мало (от 2 до 11).
При рассмотрении обобщенных данных обо всех видах и сайтах, включая агроландшафт, и ординации матрицы за счет взаимного усреднения индекс EmbAbs не отличается от ожидаемого для нулевой модели (FF), что свидетельствует об отсутствии положительной когерентности в распределении видов по сайтам. В этом случае структура метасообщества соответствует случайному типу. При исключении агроландшафта из анализа распределение видов соответствует глисоновскому градиенту (табл. 2), для которого характерна плавная сменяемость видового состава вдоль градиента при беспорядочно собранных границах диапазонов (Species ranges). Для видов, гнездящихся только в агроландшафте, наиболее подходящим шаблоном будет «шахматная доска» (Checkerboard), т. к. эмпирический индекс EmbAbs статистически значимо превышает рассчитанный для нулевой модели. В зимнее время структура метасообщества (без агроландшафта) также соответствует случайному типу (Random), но это не означает, что сообщества собраны наугад. В подобных случаях случайная структура свидетельствует о том, что виды распределены по сайтам независимо друг от друга. При ординации матрицы за счет взаимного усреднения распределение видов отряда Passeriformes соответствует глисоновскому градиенту (Gleasonian gradient). Распределение видов остальных отрядов соответствует случайному.

Рис. 4. Основные экологические градиенты в районе исследований. Сокращения: ТОЗ – тростниково-осоковые заросли по берегам озер; ЛТ – луг с тростником; СР и МР – пойменные заросли и пойменные луга по средним и малым рекам; ЛИК – луга с ивняками и мелкими колками; ОБКЛ – осиново-березовые перелески среди лугов; ОБЛ – мозаичные осиново-березовые леса междуречий; ТерС и ТерБ – террасные сосняки и террасные березняки по долинам средних рек. Ось «Х» – Component 1; Ось «Y» – Component 2
Fig. 4. The main environmental gradients in the study area. Abbreviations: ТОЗ – reed-sedge thickets along the shores of lakes; ЛТ – reed meadow; СР and МР – floodplain thickets and floodplain meadows along medium and small rivers; ЛИК - meadows with willow and small pegs; ОБКЛ – aspen-birch groves among the meadows; ОБЛ – mosaic base-birch woods of interfluves; ТерС and ТерБ – terraced pine forests and terraced birch forests in the valleys of middle rivers. Axis "X" – Component 1; Axis "Y" – Component 2
При ординации матрицы присутствия-отсутствия в соответствии с первой компонентой структура метасообщества хорошо соответствует клементсовскому градиенту (Clementsian gradient). К краям первой компоненты тяготеют относительно изолированные группы видов, населяющие, с одной стороны, заболоченные сайты, с другой – залесенные. На уровне отрядов распределение видов соответствует клементсовскому градиенту для Passeriformes. Глисоновский градиент (Gleasonian gradient) характерен для отряда Charadriiformes. Отряды Falconiformes и Piciformes демонстрируют квази-глиссоновское распределение (Quasi-Gleasonian gradient), а Anseriformes, Galliformes и Strigiformes – случайное. Вторая ось менее информативна. При ординации матрицы в соответствии со второй компонентой структура метасообщества также соответствует клементсовскому градиенту (Clementsian gradient).
Клементсовская структура метасообщества подчеркивает дискретность сообществ вдоль экологического градиента и вытеснение группой видов другие группы в пространстве. Кроме того, можно предполагать, что виды, входящие в дискретные сообщества, аналогичным образом реагируют на вариации экологических переменных или проявляют сходные исторические эффекты (Heino et al., 2016). Для изучаемого метасообщества с использованием байесовского подхода было выявлено три положительных статистически значимых ассоциации видов (Попов, 2016): 1) виды, связанные преимущественно с заболоченными участками (23.9 % от общего числа гнездящихся видов); 2) виды, связанные с древесной и кустарниковой растительностью (39.4 %); 3) виды, связанные с луговой растительность (4.9 %). Кроме того, 26.2 % видов распределены по сайтам независимо от других видов птиц. Еще 5.6 % образуют статистически значимые ассоциации только с одним видом.
Структура метасообщества зависит от доминирующего механизма, формирующего распределение видов вдоль экологического градиента, таким образом, специфичность среды обитания и отклики на абиотические компоненты очень важны (Willig, Presley, 2015) для определения видовых диапазонов (range). В этом случае структура метасообщества может быть связана с шириной экологической ниши местных видов, с положением видовых диапазонов относительно важнейшего экологического градиента и с совокупной шириной ниши всего таксона. Ранее отмечалось, что наличие в составе метасообщества большого числа видов с незначительной шириной ниши, относительно экологического градиента, за счет индивидуальных ответов видов может приводить к формированию положительного оборота видов и структур, соответствующих глисоновскому градиенту. Тогда как большое число видов с большой шириной ниши способствует незначительному обороту и формированию квази-глисоновских структур (Presley et al., 2010). Применительно к изучаемому метасообществу связь между шириной ниши и положением относительно градиента несколько сложнее. Так, например, отряд Passeriformes демонстрирует хорошо выраженный клеменсовский градиент. Для видов этого отряда ширина ниши составляет от 0.05 до 0.76 (доля заселяемых сайтов от общего числа исследованных сайтов). Медиана ширины ниши видов отряда Passeriformes вдвое больше, чем медиана для всех видов метасообщества. Совокупная ширина ниши Passeriformes охватывает весь градиент (первая компонента) – от заболоченных сайтов до сомкнутых лесов. Отряды Falconiformes и Piciformes демонстрируют квази-глисоновский градиент. Медиана ширины ниши для этих отрядов различается (критерий Краскала-Уолиса при p = 0.05): Piciformes, как правило, заселяют от 35 до 52 % сайтов (кроме Dendrocopos minor), тогда как Falconiformes заселяют от 6 до 35 % сайтов. Лишь Buteo buteo является эвритопным видом, заселяющим 47 % сайтов. Для Falconiformes совокупная ширина ниши охватывает весь градиент, тогда как Piciformes заселяют только сайты, имеющие древесную растительность.
Случайный тип структуры метасообщества отрядов Galliformes и Strigiformes, возможно, связан с узкой шириной ниши видов и небольшим перекрытием видовых диапазонов. Для отряда Anseriformes, также демонстрирующего случайную структуру, ожидалось, что распределение видов будет соответствовать вложенным подмножествам, но из-за большого количества встроенных отсутствий этого не наблюдается.
Сменяемость видового состава гнездящихся и возможно гнездящихся птиц, выявленная за счет взаимного усреднения, в целом соответствует основному экологическому градиенту в районе исследований. Корреляция между первой компонентой и осью, полученной за счет взаимного усреднения, составляет 0.912 (Спирмен) и 0.780 (Кендалл) при p < 0.001. Корреляция между второй компонентой и осью, полученной за счет взаимного усреднения, статистически не значима. Географическая широта также не оказывает влияние на сменяемость видового состава птиц в пределах метасообщества.
Положительная связь с лесистостью отмечена для 28 видов. Для 25 видов связь с лесистостью отрицательная. 62.2 % видов в своем распределении по сайтам статистически не связаны с лесистостью. Эти виды не предпочитают и не избегают лесистость. Положительная связь с лесистостью характерна для Turdus iliacus, Streptopelia orienthalis, Parus major, Parus montanus, Dendrocopos major, Corvus corax, Tetrao urogallus, Cuculus saturatus, Strix uralensis, Oriolus oriolus и некоторых других видов. Для 31 вида характерна положительная связь с числом видов деревьев в первом ярусе.
Обводненность – второе плечо первой компоненты – предпочитает 21 вид и избегают 14 видов. Рекреационная нагрузка сказывается на распределении 16.4 % видов, входящих в метасообщество. 30 видов птиц, гнездящихся в наименее измененных сайтах (без учета агроландшафта), демонстрируют положительную связь с небольшой сельскохозяйственной нагрузкой в пределах сайта.
Кроме реакции на градиенты среды сообщества могут быть структурированы за счет вложенности (Nestedness), связанной с селективной колонизацией или вымиранием, а также с вложенностью самих сайтов. При анализе оборота видов (Species turnover) как вдоль первой компоненты, так и при ординации матрицы за счет взаимного усреднения вложенные структуры, описанные для многих природных сообществ, не были отмечены. Специальный анализ (Nestedness analysis), при котором проводится сортировка матрицы для наилучшего выявления вложенных структур, также не выявил статистической значимости индекса NODF. Вложенность не была выявлена и на уровне отрядов и семейств. Возможно, отсутствие вложенных структур связано с отсутствием коротких и сильных градиентов, высокой мозаичностью, а также с отсутствием четко выраженных границ между сайтами.
Помимо экологических градиентов на распределение видов птиц могут оказывать влияние и пространственные процессы – расстояние между сайтами. В пределах исследуемого метасообщества и при данном масштабе исследований корреляция между близостью сайтов и сходством видового состава птиц статистически не значима. В условиях высоко мозаичных ландшафтов соседние сайты могут быть принципиально различными, что увеличивает несходство населения птиц. Можно предполагать, что роль неэкологических пространственных процессов будет значительна в однообразной местности, например, в условиях обширного сплошного лесного массива или степи. В пределах же исследуемого региона, находящегося на границе тайги и лесостепи, мозаичность ландшафтов очень велика, и ведущее значение имеют процессы сортировки видов на основе экологической ниши и избирательности видов относительно факторов среды.
Распределение птиц по сайтам может быть связано и с межвидовыми отношениями. Для тестирования гипотезы о роли межвидовой конкуренции использовалось также и «таксономическое просеивание». В этом случае расчет индексов проводился только в пределах какого-либо отряда в данном сайте, а не всех видов вместе. Кроме того, проводился расчет дисперсии различий в размерах тела для всего регионального пула видов и раздельно для отрядов.
Полученные данные свидетельствуют о том, что для большинства сайтов дисперсия различий размеров тела птиц не отличается от случайной, рассчитанной с помощью нулевых моделей (табл. 3). Для всех изученных сайтов не подтверждается гипотеза о том, что сообщества могут быть структурированы за счет межвидовой конкуренции, приводящей к формированию определенных интервалов между размерами тела. Наличие определенных и постоянных интервалов между размерами тела были выявлены только в нескольких сайтах у видов отрядов Anseriformes, Galliformes, Charadriiformes и Passeriformes. Статистически значимые различия наблюдаемой и ожидаемой дисперсии для этих отрядов были выявлены только в пределах 1–2 сайтов. При исследовании размеров тела у видов, входящих в статистически значимые положительные ассоциации, различия наблюдаемой и ожидаемой дисперсии оказались также не значимыми.
Для 6 из 29 сайтов наблюдаемая дисперсия была выше случайной, рассчитанной с помощью нулевой модели. Для положительных ассоциаций видов, связанных с луговой и древесной растительностью, также выявлено превышение наблюдаемой дисперсии над случайной (см. табл. 3). В ассоциации видов, связанных с заболоченными стациями, дисперсия различий размеров тела видов не отличается от случайной. Агрегированность в размерах тела выявлена в отрядах Charadriiformes и Passeriformes. Можно предполагать, что среда действует как экологический фильтр, препятствующий вселению видов с иными размерами тела. На уровне регионального пула видов дисперсия различий размеров тела не отличается от случайной. Лишь для отрядов Passeriformes и Falconiformes выявлена агрегированность размеров тела.
В целом полученные результаты не позволяют говорить о том, что население гнездящихся и возможно гнездящихся видов птиц формируется под влиянием межвидовой конкуренции, для снижения которой экологически и генетически родственные виды имеют какие-либо минимальные различия в размерах тела.
Таблица 2. Результаты исследований элементов структуры метасообщества птиц на границе тайги и лесостепи Зауралья. PZ(H0) – вероятность принятия нулевой гипотезы. Уровень значимости p ≤ 0.05. Жирным шрифтом выделены статистически значимые индексы
| Species | Когерентность (Coherence) | Оборот видов (Species turnover) | Группировка границ
(Boundary clumping) |
Идеализированный образец (шаблон) метасообщества | |||||||
| Emb
Abs |
PZ
(H0) |
M | SD | C-Score | PZ
(H0) |
M | SD | Mori
sita |
PZ
(H0) |
||
| Ординация за счет взаимного усреднения | |||||||||||
| Все гнездящиеся виды района исследований | 0.869 | 0.07 | 1.365 | 0.28 | 0.031 | 0.00 | 0.03 | 0.00 | 2.44 | 0.11 | Случайное распределение |
| Все гнездящиеся виды без опубликованных данных по агроландшафту | 0.409 | 0.00 | 0.841 | 0.05 | 0.064 | 0.00 | 0.06 | 0.00 | 1.56 | 0.34 | Градиент Глисона |
| Виды, гнездящиеся только в агроландшафте (данные В. А. Коровина) | 0.472 | 0.02 | 0.345 | 0.05 | 0.016 | 0.56 | 0.02 | 0.00 | 2.34 | 0.02 | Шахматная доска |
| Зима (без агроландшафта, только собственные данные) | 0.298 | 0.29 | 0.543 | 0.23 | 0.021 | 0.004 | 0.02 | 0.00 | 1.96 | 0.17 | Случайное распределение |
| Ординация в соответствии с первой компонентой | |||||||||||
| Все гнездящиеся виды (только собственные данные) | 0.532 | 0.00 | 0.916 | 0.03 | 0.064 | 0.00 | 0.06 | 0.00 | 1.89 | 0.00 | Градиент Клементса |
| Anseriformes (только собственные данные) |
0.625 | 0.14 | 0.724 | 0.07 | 0.006 | 0.97 | 0.01 | 0.00 | 5.73 | 0.00 | Случайное распределение |
| Falconiformes (только собственные данные) |
0.429 | 0.00 | 1.083 | 0.14 | 0.027 | 0.83 | 0.03 | 0.001 | 1.34 | 0.89 | Квази-клементсовский градиент |
| Galliformes (только собственные данные) |
1.105 | 0.22 | 1.335 | 0.19 | 0.050 | 0.34 | 0.05 | 0.002 | 0.75 | 0.39 | Случайное распределение |
| Charadriiformes (только собственные данные) |
0.798 | 0.00 | 1.117 | 0.11 | 0.036 | 0.00 | 0.03 | 0.00 | 2.68 | 0.51 | Градиент Глисона |
| Strigiformes (только собственные данные) |
1.000 | 0.22 | 1.298 | 0.24 | 0.051 | 0.81 | 0.05 | 0.004 | 0.61 | 0.83 | Случайное распределение |
| Falconiformes + Strigiformes (только собственные данные) |
0.600 | 0.00 | 1.143 | 0.12 | 0.029 | 0.97 | 0.03 | 0.001 | 1.51 | 0.50 | Квази-глисоновский градиент |
| Piciformes (только собственные данные) |
0.324 | 0.01 | 0.400 | 0.03 | 0.021 | 0.72 | 0.02 | 0.001 | 1.89 | 0.23 | Квази-глисоновский градиент |
| Passeriformes (только собственные данные) |
0.484 | 0.00 | 0.721 | 0.03 | 0.060 | 0.00 | 0.05 | 0.00 | 2.46 | 0.00 | Градиент Клементса |
| Ординация в соответствии со второй компонентой | |||||||||||
| Все гнездящиеся виды (только собственные данные) | 1.016 | 0.00 | 1.165 | 0.04 | 0.064 | 0.00 | 0.06 | 0.00 | 2.08 | 0.02 | Градиент Клементса |
| Ординация север-юг | |||||||||||
| Все гнездящиеся виды (только собственные данные) | 1.081 | 0.99 | 1.080 | 0.03 | 0.064 | 0.00 | 0.06 | 0.00 | 1.83 | 0.01 | Случайное распределение |
Таблица 3
Метаанализ размеров тела птиц для модели перекрытия размеров тела (по аналогии с [23]) на границе тайги и лесостепи Зауралья. «Lower tail» и «Upper tail» – количество сайтов, для которых наблюдаемая дисперсия разницы между длиной тела была меньше или больше случайной, вычисленной с помощью нулевой модели. Число в скобках – это число сайтов, для которых различия в дисперсии статистически значимы (p < 0.05). Стандартизированный эффект размера (SES) = (Iobs - Isim)/Ssim , где Isim – это средний показатель для нулевой модели, Ssim – стандартное отклонение, Iobs – наблюдаемый показатель для населения птиц в районе исследований. Для сообществ, структурированных за счет межвидовой конкуренции, рассчитанные индексы должны быть ниже, чем для нулевой модели, а паттерн метаанализа должен быть выше нуля. Одновыборочный t-критерий был использован для проверки гипотезы о том, что SES для множества сообществ не отличается от нуля
Жирным шрифтом выделены группы видов, для которых выявлены статистически значимые различия между наблюдаемой дисперсией различий размеров тела и случайной. Расчеты проводились только для сайтов, содержащих не менее 3 видов. P – на уровне отдельных сайтов – уровень значимости при расчетах t-критерия; на уровне регионального пула видов – уровень значимости при сравнении наблюдаемого индекса и среднего индекса, предсказанного с помощью нулевой модели FF
| Группы видов | Всего сайтов, исполь-зованных для анализа | Lower tail | Upper tail | Среднее значение наблю-даемого индекса | Среднее значение индекса для нулевых моделей | Дисперсия среднего значения индекса для нулевых моделей | t | P |
| Отдельные сайты | ||||||||
| Все гнездящиеся и возможно гнездящиеся виды | 29 | 13 (0) | 16 (6) | 0.0028 | 0.00303 | 0.0000218 | 2.647 | 0.013 |
| Положительная ассоциация видов, связанная с заболоченными стациями | 16 | 10 (0) | 6 (0) | 0.00664 | 0.00915 | 0.000124 | -0.853 | 0.407 |
| Положительная ассоциация видов, связанная с луговыми стациями | 9 | 2 (0) | 7 (6) | 0.031859 | 0.010807 | 0.000138 | 3.470 | 0.008 |
| Положительная ассоциация видов, связанных с древесной растительностью | 14 | 3 (0) | 11 (2) | 0.00337 | 0.003036 | 0.00004 | 2.615 | 0.021 |
| Podicipediformes | 4 | 2 (0) | 2 (0) | 0.01218 | 0.00763 | 0.00005 | 0.623 | 0.577 |
| Anseriformes | 6 | 2 (1) | 4 (0) | 0.00357 | 0.00387 | 0.000007 | -1.610 | 0.168 |
| Galliformes | 3 | 3 (2) | 0 (0) | 0.00033 | 0.0082 | 0.000057 | -77.457 | <0.001 |
| Falconiformes | 7 | 2 (0) | 5 (0) | 0.00345 | 0.00343 | 0.000016 | 1.759 | 0.129 |
| Charadriiformes | 11 | 2 (1) | 9 (1) | 0.00406 | 0.00511 | 0.000042 | 1.392 | 0.194 |
| Strigiformes | 1 | 1 (0) | 0 (0) | 0.0124 | 0.00514 | 0.00002 | - | - |
| Piciformes | 3 | 3 (0) | 0 (0) | 0.00921 | 0.01333 | 0.000123 | -1.372 | 0.304 |
| Passeriformes | 24 | 10 (2) | 14 (10) | 0.014202 | 0.006126 | 0.000087 | 2.964 | 0.007 |
| Sylviidaea | 14 | 6 (0) | 8 (1) | 0.000729 | 0.000396 | 0.000000 | 1.525 | 0.151 |
| Региональный пул видов | ||||||||
| Все гнездящиеся и возможно гнездящиеся виды вместе | - | - | - | 0.00248 | 0.00181 | 0.00000 | - | 0.171 |
| Podicipediformes | - | - | - | 0.00291 | 0.00521 | 0.00002 | - | 0.352 |
| Anseriformes | - | - | - | 0.00627 | 0.00564 | 0.00001 | - | 0.323 |
| Galliformes | - | - | - | 0.00131 | 0.00343 | 0.00001 | - | 0.162 |
| Falconiformes | - | - | - | 0.00106 | 0.00053 | 0.00000 | - | 0.043 |
| Charadriiformes | - | - | - | 0.00106 | 0.00103 | 0.00000 | - | 0.378 |
| Strigiformes | - | - | - | 0.00630 | 0.00514 | 0.00001 | - | 0.315 |
| Piciformes | - | - | - | 0.00507 | 0.00976 | 0.00005 | - | 0.275 |
| Passeriformes | - | - | - | 0.00065 | 0.00020 | 0.00000 | - | 0.002 |
Заключение
Структура метасообщества птиц на границе тайги и лесостепи Зауралья, содержащего достаточно большое число видов с узкой экологической нишей, достоверно соответствует глисоновскому градиенту (для наименее измененных сайтов). При ординации метасообщества в соответствии с важнейшим экологическим градиентом структура метасообщества птиц соответствует клементсовскому градиенту. Процессы, приводящие к формированию вложенных подмножеств, не характерны. В пределах переходной зоны между тайгой и лесостепью, в связи с небольшой протяженностью района исследований, роль экологического структурирования метасообщества за счет локального изменения переменных среды, по сравнению с влиянием пространственной составляющей, преобладает. Существующий градиент север-юг статистически не связан с изменением видового состава сайтов. При ординации матрицы присутствия-отсутствия в направлении север-юг, что соответствует смене природных зон, когерентность отсутствует, а структура метасообщества соответствует случайному типу. Увеличение сходства между видовым составом сайтов при уменьшении расстояния между этими сайтами статистически не выявлено. Скорее всего, пространственная структура метасообщества формируется весной в результате ежегодной селективной колонизации за счет экологической фильтрации и сортировки видов, а процессы расселения, массовые эффекты и конкуренция, видимо, не существенны при ежегодном формировании метасообщества. В связи с высокой мозаичностью лесостепи в пределах одного сайта имеется множество локальных участков, подходящих для гнездования разных видов. В качестве предположения можно высказать гипотезу о том, что первые стадии формирования сообщества гнездящихся птиц в данный год происходят в послегнездовое время предыдущего года, когда у молодых особей, а у некоторых видов и у взрослых, происходит запечатление (импринтинг) района будущего гнездования. Но для проверки этой гипотезы нужны тщательные исследования послегнездовой жизни всех местных видов и сравнение послегнездового распределения данного года с распределением гнездящихся особей на следующий год. Наличие большого числа видов, распределение которых является независимым или случайным, может быть связано с невысокой численностью этих видов, находящихся весной в условиях свободного выбора мест гнездования. В противоположность этому виды с высокой численностью заселяют все пригодные участки.
Библиография
Горчаковский П. Л. Растительность // Урал и Приуралье. М.: Наука, 1968. С. 211–261.
Коблик Е. А., Редькин Я. А., Архипов В. Ю. Список птиц Российской Федерации . М.: Товарищество научных изданий КМК, 2006. 256 с.
Коровин В. А. Птицы в агроландшафтах Урала . Екатеринбург: Изд-во Урал. ун-та, 2004. 504 с.
Орлова В. В. Западная Сибирь . Л.: Гидромет. изд-во, 1962. 360 с.
Попов С. В. Паттерны совместной встречаемости видов в сообществах птиц на границе тайги и лесостепи Зауралья // Вестник Удмуртского государственного университета. Сер.: Биология. Науки о Земле. 2016. Т. 31. Вып. 3. С. 83–97.
Птицы Советского Союза . Т. 1 / Под ред. Г. П. Дементьева и Н. А. Гладкова. М.: Сов. наука, 1951. 653 с.
Птицы Советского Союза . Т. 2 / Под ред. Г. П. Дементьева и Н. А. Гладкова. М.: Сов. наука, 1951. 485 с.
Птицы Советского Союза . Т. 3 / Под ред. Г. П. Дементьева и Н. А. Гладкова. М.: Сов. наука, 1951. 685 с.
Птицы Советского Союза . Т. 4 / Под ред. Г. П. Дементьева и Н. А. Гладкова. М.: Сов. наука, 1952. 647 с.
Птицы Советского Союза . Т. 5 / Под ред. Г. П. Дементьева и Н. А. Гладкова. М.: Сов. наука, 1954. 808 с.
Птицы Советского Союза . Т. 6 / Под ред. Г. П. Дементьева и Н. А. Гладкова. М.: Сов. наука. 1954. 796 с.
Bert D. G. A multiscale analysis of Nested Species subsets of forest Bird in agricultural Landscapes near Ottawa, Canada. A thesis submitted to the Faculty of Graduate Studies and Research in partial fulfillment of the requirements for the degree of Masters of Science Department of Biology Carleton University Ottawa. Ontario, 2001. 100 p.
Boecklen W. Nestedness, biogeographic theory, and the design of nature reserves // Oecologia. 1997. Vol. 112. P. 123–142.
Diamond J. M. Assembly of species communities // Ecology and evolution of communities. 1975. P. 342–444.
Gotelli N. J., Ellison A. M. Assembly rules for New England ant assemblages // Oikos. 2002. Vol. 99. P. 591–599.
Gotelli N. J., Entsminger G. L. EcoSim: Null models software for ecology. Version 7.0. Acquired Intelligence Inc. & Kesey-Bear. 2001. URL: http://homepages.together.net/~gentsmin/ecosim.htm.
Heino J., Soininen J., Alahuhta J., Lappalainen J., Virtanen R. Metacommunity ecology meets biogeography: effects of geographical region, spatial dynamics and environmental filtering on community structure in aquatic organisms // Oecologia. 2016. Vol. 183 (1). P. 121–137. DOI: 10.1007/s00442-016-3750-y.
Leibold M. A., Mikkelson G. M. Coherence, species turnover, and boundary clumping: elements of meta-community structure // Oikos. 2002. Vol. 97. P. 237–250.
Leibold M. A., Holyoak M., Mouquet N., Amarasekare P., Chase J. M., Hoopes M. F., Holt R. D., Shurin J. B., Law R., Tilman D., Loreau M., Gonzalez A. The metacommunity concept: a framework for multi-scale community ecology // Ecology Letters. 2004. Vol. 7. P. 601–613.
Matthews T. J., Sheard C., Cottee-Jones H. E. W., Bregman T. P., Tobias J. A., Whittaker R. J. Ecological traits reveal functional nestedness of bird communities in habitat islands: a global survey // Oikos. 2015. Vol. 124. № 7. P. 817–826.
Meynard C. N., Quinn J. F. Bird Metacommunities in temperature South American forest: vegetation structure, area and Climate effects // Ecology. 2008. Vol. 89 (4). P. 981–990.
Morrison L. W. Nestedness in insular floras: spatiotemporal variation and underlying mechanisms // Journal of Plant Ecology. 2013. Vol. 6 (6). P. 480–488.
Ozkan K., Svenning J.-C., Jeppesen E. Environmental species sorting dominates forest-bird community assembly across scales // Journal of Animal Ecology. 2013. Vol. 82 (1). P. 266–274. DOI: 10.1111/j.1365-2656.2012.02019.x.
Patterson B. D., Atmar W. Nested subsets and the structure of insular mammalian faunas and archipelagos // Heaney L. R., Patterson B. D. (eds). Island biogeography of mammals. London, Academic Press, 1986. P. 65–82.
Podani J., Schmera D. A comparative evaluation of pairwise nestedness measures // Ecography. 2012. Vol. 35. P. 001–012.
Presley J. S., Higgins C. L., Lopez-Gonzalez C., Stevens R. D. Elements of metacommunity structure of Paraguayan bats: multiple gradients require analysis of multiple ordination axes // Oecologia. 2009. Vol. 160. P. 781–793. DOI: 10.1007/s00442-009-1341-x.
Presley S. J., Higgins C. L., Willig M. R. A comprehensive framework for the evaluation of metacommunity structure // Oikos. 2010. Vol. 119. P. 908–917.
Royan A., Reynolds S. J., Hannah D. M., Prudhomme C., Noble D. G., Sadler J. P. Shared environmental responses drive co-occurrence patterns in river bird communities // Ecography. 2016. Vol. 39. P. 733–742.
Sebastián-González E., Botella F., Paracuellos M., Sanchez-Zapata J. A. Processes driving temporal dynamics in the nested pattern of water bird communities // Acta Oecologica. 2010. Vol. 36. P. 160–165.
Sybertz J., Reich M. Assessing Climate Change Induced Turnover in Bird Communities Using Climatically Analogous Regions // Diversity. 2015. Vol. 7. P. 36–59.
Ulrich W., Almeida-Neto M., Gotelli N. G. A consumer’s guide to nestedness analysis // Oikos. 2009. Vol. 118. P. 3–17.
Ulrich W. Turnover - a FORTRAN program for analysis of species associations. Version 1.1. 2012. URL: www.umk.pl/~ulrichw. Turnover.
Vilmi A., Tolonen K. T., Karjalainen S. M., Heino J. Metacommunity structuring in a highly-connected aquatic system: effects of dispersal, abiotic environment and grazing pressure on microalgal guilds // Hydrobiologia. 2016. P. 1–16. DOI: 10.1007/s10750-016-3024-z.
Wang Y., Chen S., Ding P. Testing multiple assembly rule models in avian communities on islands of an inundated lake, Zhejiang Province, China // Journal of Biogeography. 2011. Vol. 38. P. 1330–1344.
Willig M. R., Presley S. J. Biodiversity and metacommunity structure of animals along altitudinal gradients in tropical montane forests // Journal of Tropical Ecology. 2015. P. 1–16. DOI: 10.1017/S0266467415000589.

 © 2011 - 2026
© 2011 - 2026