



Введение
Некоторые стволовые насекомые способны размножаться в массе и вызывать изменение состава верхнего яруса таежных экосистем. Допустимо предполагать, что жуки при дополнительном питании выбирают виды деревьев, в которых развивались их личинки. Ниже следует описание результатов трехлетних наблюдений за пищевыми предпочтениями черного пихтового усача Monochamus urussovi Fisch. – многочисленного вида на пихте сибирской Abies sibirica Ledeb. Для сравнения был выбран малый черный усач M. sutor L. – обычный таежный вид, который не образует очагов массового размножения и никогда не был обнаружен на пихте (рис. 1).
Оба вида давно привлекают внимание энтомологов, и их названия на русском языке в разных трудах различны. M. urussovi называли большим черным хвойным, большим черным еловым, большим еловым усачом (или дровосеком) (Коломиец, 1960; Земкова, 1965; Криволуцкая, 1965; Петренко, 1965; Богданова, 1976; Кондаков и др., 1979; Гусев, 1984; Никитский, Ижевский, 2005). M. sutor также упоминали как малого черного елового, черного елового, малого черного усача (или дровосека) (Коломиец, 1960; Земкова, 1965; Петренко, 1965; Агафонова, Антонов, 2014).

Рис. 1. Черный пихтовый и малый черный усачи
Fig. 1. Black fir and small black sawyers
M. urussovi размножается в массе только на пихте и только в Сибири (Прозоров, 1958; Исаев, 1971; Исаев и др., 1982; Ветрова, 1986, 1987; Исаев и др., 1988; Ветрова и др., 1998), хотя встречается повсюду от восточных до западных границ бывшего Советского Союза: в коллекции Зоологического института РАН в Санкт-Петербурге хранятся лишь единичные находки этого вида из Финляндии и Польши, а в Карелии вид заслуживает внесения в Красную книгу (Е. Б. Яковлев, личное сообщение).

Рис. 2. Личинка черного пихтового усача
Fig. 2. A larva of black fir sawyer
Из-за своих крупных размеров личинки жука (рис. 2) не могут развиваться на подросте, а только на деревьях диаметром не менее чем 10–12 см, хотя молодые пихты в очагах страдают от объедания молодыми имаго при дополнительном питании. Поэтому M. urussovi может считаться вредоносным насекомым по отношению к лесосырьевой базе, но не к пихтарнику как лесной экосистеме. Пищевые предпочтения имаго прежде не изучали.
Целью работы было определение избирательности дополнительного питания жуков черного пихтового и малого черного усачей в отношении разных видов деревьев.
Материалы
Работа проведена на северном макросклоне Западного Саяна. Куколки и молодые жуки M. urussovi были извлечены из стволов пихты сибирской, жуки M. sutor отловлены в природе, поэтому было невозможно определить, в каких деревьях развивались личинки второго вида; однако в месте вылова имаго M. sutor нет других хвойных деревьев, кроме сосны. В опытах использованы 132 особи M. urussovi и 40 особей M. sutor.
Методы
Жукам (в т. ч. выведенным из куколок) предлагали на выбор побеги семи видов деревьев: пихты сибирской Abies sibirica Ledeb., ели сибирской Picea obovata Ledeb., сосны кедровой сибирской Pinus cembra Du Tour, сосны обыкновенной Pinus sylvestris L., лиственницы сибирской Larix sibirica Ledeb., березы бородавчатой Betula pendula Roth, ивы козьей Salix caprea L. Все побеги были одинаковой длины и диаметра.
Каждого жука по одному помещали в отдельный пластмассовый садок стандартного размера 30 х 20 см, где были разложены побеги, и накрывали стеклом; хвоя и листья с побегов были предварительно удалены. В течение суток жуки питались корой побегов, после чего измерялась площадь коры, съеденной насекомыми: когда жук грызет кору, он пятится назад, так что повреждение имеет форму почти правильного прямоугольника (незначительно закруглены края): длину и ширину погрыза измеряли под бинокуляром с микрометрической линейкой, площадь погрыза вычисляли умножением длины на ширину. Повторно жуки в опыте не использовались.
Результаты
Опыт показал, что черный пихтовый усач явно предпочитает питаться корой сосны, отдавая предпочтение по сравнению с пихтой даже березе (табл. 1); в обследованных районах обнаружены упавшие березовые стволы с круглыми выходными отверстиями черного пихтового усача, диаметр которых достигает 11 мм, поэтому их нельзя перепутать с выходными отверстиями других ксилофагов, а на стоящих березах найдены типичные насечки, которые самки выгрызают в коре для откладывания яиц (Андреева и др., 1999) (рис. 3); при этом вспышки численности усача в березовых древостоях не отмечены.
Таблица 1. Средняя площадь погрызов M. urussovi на предложенных породах (мм2) с учетом доверительного интервала 90 %; число подопытных жуков – 132 особи
| Пихта | Ель | Кедр | Сосна | Лиственница | Береза | Ива |
| 13.03 ± 1.40 | 51.11 ± 2.55 | 4.00 ± 0.22 | 107.08 ± 4.33 | 6.04 ± 0.21 | 22.76 ± 1.77 | 6.86 ± 0.36 |
Похожий результат был получен в отношении короеда Dendroctonus ponderosae, потомство которого, перечисленное на число отложенных яиц, много меньше именно на том виде сосны, из стволов которого вышли самки, чем на других американских видах сосен, как нативных, так и интродуцированных (Amman, 1982).

Рис. 3. Насечки черного пихтового усача на пихте и березе, выходные отверстия, проделанные молодыми жуками при выходе из пихтового ствола. Разница в форме насечек обусловлена толщиной и жесткостью коры: у пихты она тоньше и мягче, поэтому в бересте, чтобы отложить яйцо в луб, самке приходится выгрызать более длинные и глубокие щели
Fig. 3. Black fir sawyer egg-laying slots in fir and birch bark, holes made by young beetles emerging from fir trunk. The difference between shape of the slots depends on thickness and hardness of the bark, which is considerably thinner and softer in fir, than in birch. Hence, to lay eggs under birch bark, females have to make longer and deeper slots
Пищевые предпочтения малого черного хвойного усача отличаются от таковых черного пихтового усача (табл. 2). Этот вид полностью избегает пихту, однако также предпочитает кору и луб сосны.
Таблица 2. Средняя площадь погрызов M. sutor на предложенных породах (мм2) с учетом доверительного интервала 90 %; число подопытных жуков – 40 особей
| Пихта | Ель | Кедр | Сосна | Лиственница | Береза | Ива |
| 0 | 16.85 ± 1.37 | 15.85 ± 2.10 | 121.85 ± 2.05 | 8.12 ± 0.91 | 4.02 ± 0.31 | 7.12 ± 1.01 |
Обсуждение
Ксилофаги при дополнительном питании переносят офиостомовые грибы (Ветрова и др., 1992) (рис. 4), возбуждающие синеву древесины – микоз, который вызывает порыжение (такие побеги в научном обиходе именуются «рыжиками»), покраснение и усыхание хвои на дистальных частях побегов выше погрыза, тем самым помогая жукам ослабить дерево, прежде чем отложить под кору яйца (Исаев и др., 1982, 1988); в ослабленном дереве прекращается синтез защитных смолистых веществ, и личинки жуков могут благополучно расти. Мутуалистические взаимоотношения, сложившиеся между ксилофагом и грибом, определяют характер дополнительного питания молодых имаго: для устойчивого существования популяции насекомого необходимо, чтобы часть деревьев в лесном насаждении была заражена синевой древесины.
Выбор корма для дополнительного питания жуками M. sutor не удивителен: жуки предпочитают питаться на том же виде дерева, в котором развились их личинки.
Черный пихтовый усач ведет себя иначе. Этот вид распространен по всей лесной территории умеренных широт от Финляндии, Польши и Белоруссии до Монголии, Магаданской области и Камчатки. Южная граница его ареала проходит по пойменным соснякам Приднепровья, казахскому мелкосопочнику и Северному Китаю – везде, где растет сосна. Естественно предположить, что исторически вид M. urussovi, как и другие виды р. Monochamus, сформировался в тесной связи с сосной, с чем можно предположительно связать пищевые предпочтения, проявленные жуками в условиях эксперимента.
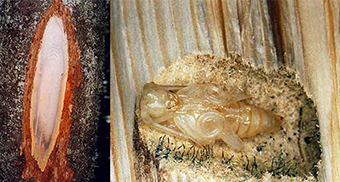
Рис. 4. Синева древесины и плодовые тела офиостомового гриба, окружающие куколку усача в колыбельке
Fig. 4. Log bluing and fruit bodies of Ceratocystis sp., surrounding a pupa of black fir sawyer in its pupal chamber
Тем не менее в сосняках вспышки численности M. urussovi никогда не были отмечены. Отчего массовое размножение происходит только в пихтарниках – разновидности леса, характерной исключительно для сибирской южной тайги, хотя жуки, личинки которых развивались в пихтовых стволах, при дополнительном питании редко избирают пихту, но размножаются в массе именно на ней, а не на сосне?
Возможный ответ: на сосне у усача есть эффективные конкуренты – быстро размножающиеся короеды, которые вытесняют усача на не предпочитаемую им пихту. На пихте у усача в Сибири конкурентов мало. Можно упомянуть в числе жуков ребристого рагия Rhagium inquisitor L., пальцеходного лубоеда Xylechinus pilosus Ratz., полосатого древесинника Trypodendron lineatum Oliv., темную хвойную златку Buprestis haemorrhoidalis Hbst., хвойное сверлило Hylecoetus flabellicornis Schn.; среди перепончатокрылых – большого хвойного рогохвоста Uroceros gigas L. (Андреева и др., 1999). Перечисленные виды не образуют очагов массового размножения в пихтовых древостоях, хотя некоторые отмирающие деревья бывают плотно заселены пальцеходным лубоедом Xylechinus pilosus Ratz. Однако для крупных личинок M. urussovi мелкие личинки других видов могут послужить калорийным кормом.
В значимости конкурентов для вытеснения M. urussovi с сосны допустимо сомневаться, поскольку в европейской части России черный пихтовый усач успешно развивается на ели, где конкурентов у него гораздо больше, чем на сосне. Несмотря на возможность питания личинок тканями елового ствола, массовое размножение усача в ельниках не происходит, а в Сибири ельники, свободные от других видов деревьев, встречаются редко – только в поймах рек.
У усача, помимо конкурентов, есть только один природный враг – черный дятел, желна Dryocopus martius L (рис. 5). Однако черный дятел – оседлый вид, который питается личинками усача только зимой, а в теплые сезоны предпочитает легкую добычу: открыто живущих насекомых, прежде всего – муравьев.

Рис. 5. Усыхающая пихта в очаге усача и пихтовый ствол, расклеванный желной в поисках личинок (на заднем плане видны «рыжики»)
Fig. 5. Dead fir tree in the area of black fir sawyer mass outbreak and a fir trunk treated by black woodpecker (red fir shoots can be seen in the background)
Чем объяснить массовое размножение черного пихтового усача в пихтарниках? Возможно, приуроченность очагов этого ксилофага исключительно к пихтовым лесам связана с неспособностью его симбионтов – офиостомовых грибов – развиваться на других хвойных деревьях кроме пихты (речь идет только о тех видах грибов, которые связаны с M. urussovi, но не с другими ксилофагами), однако результаты исследований специфичности возбудителей синевы древесины в отношении разных видов хвойных деревьев авторам неизвестны, по крайней мере, покраснение концевых частей побегов на других хвойных не отмечено.
Другое гипотетическое объяснение может быть основано на данных по истории лесов. Опубликованы результаты исследований, говорящие о важности эколого-географического фактора в формировании трофических связей насекомых с растениями (Nylin, Janz, 1996), соответственно, важен и исторический возраст растительных сообществ. Нельзя сказать, что темнохвойная тайга – молодой тип растительных формаций; более точно говорить о молодости лесных экосистем, находящихся севернее Алтае-Саянской горной страны.
Равнинные пихтовые древостои сформировались менее чем чем 10 тыс. л. н., после того, как отступило последнее в Сибири Зырянское оледенение, относящееся к позднему плейстоцену (Большая..., 1972; Геологический…, 1973). Спорово-пыльцевой и радиоуглеродный анализы почв показывают, что последний пик распространения темнохвойной тайги на юге Сибири датируется менее чем 1 000 л. н., а в оптимальную фазу голоцена (ок. 5 000 л. н.) здесь были развиты лесостепные ландшафты с березовыми и сосновыми лесами (Савина, 1976, 1986). Сделан вывод, что в настоящее время и в ближайшем будущем в горных лесах темнохвойные формации распространяются и будут распространяться от горной черневой тайги вниз на подтайгу; расширение площади, занятой темнохвойными породами на равнинах, может происходить при ведущей роли ели (но не пихты и кедра) на склонах водоразделов (Савина, 1976). О том же говорит и исключительная активность массовых насекомых – вредителей пихты сибирской: сибирского коконопряда Dendrolimus superans sibiricus Tschetw. и черного пихтового усача в равнинной тайге (Гродницкий и др., 2002; Шабалина и др., 2002; Гродницкий, 2004).
Отсюда может следовать, что у пихты не было исторического времени, достаточного для развития приспособлений к эффективной защите от заселения усачом, отчего возникают вспышки численности M. urussovi.
Заключение
При дополнительном питании черный пихтовый усач достоверно предпочитает сосновые побеги пихтовым. Вместе с тем массовое размножение этого усача отмечено только в древостоях с преобладанием пихты сибирской. Проведенное исследование не позволяет утверждать, что вспышки численности определяются особым предпочтением жука к питанию на пихте. Возможно, жесткая приуроченность очагов к пихтарникам связана со сравнительной молодостью экосистем, где пихта является породой-эдификатором. По всей видимости, пихтовая тайга юга Сибири является переходной, исторически недолговечной формацией.
Библиография
Агафонова Т. А., Антонов И. А. Каталог насекомых-ксилофагов хвойных пород Байкальской Сибири и Северного Приамурья . Иркутск: Изд-во Ин-та географии им. В. Б. Сочавы СО РАН, 2014. 215 с.
Андреева Т. П. и др. Насекомые сибирских лесов. Первый атлас цветных фотографий для специалистов лесного хозяйства . Красноярск: Центр защиты леса, 1999. 96 с.
Богданова Д. А. Стволовые вредители и динамика численности их на вырубках // Проблема динамики численности насекомых-вредителей таежных лесов. Красноярск: Изд-во Института леса и древесины им. В. Н. Сукачева СО АН СССР, 1976. С. 12–29.
Большая советская энциклопедия . М.: Советская энциклопедия, 1972. Т. 9. 624 с.
Ветрова В. П. Воздействие на пихтовые древостои разреженных и повышенно плотных популяций большого черного хвойного усача // Лесоведение. 1986. № 1. С. 43–50.
Ветрова В. П. Консортивные связи большого черного хвойного усача с пихтой сибирской в таежных биогеоценозах: Автореф. дис. ... канд. биол. наук . Красноярск, 1987. 22 с.
Ветрова В. П., Исаев А. С., Пашенова Н. В., Константинов М. Ю. Оценка угрозы массового размножения черного пихтового усача в темнохвойных насаждениях Нижнего Приангарья после повреждения сибирским шелкопрядом // Лесоведение. 1998. № 3. С. 58–67.
Ветрова В. П., Пашенова Н. В., Гродницкий Д. Л. Реакция пихты сибирской на заражение грибами – симбионтами черного пихтового усача // Лесоведение. 1992. № 3. С. 24–32.
Геологический словарь . М.: Недра, 1973. Т. 2. 456 с.
Гродницкий Д. Л. Сибирский шелкопряд и судьба пихтовой тайги // Природа. 2004. № 11. С. 49–56.
Гродницкий Д. Л., Разнобарский В. Г., Солдатов В. В., Ремарчук Н. П. Деградация древостоев в таежных шелкопрядниках // Сибирский экологический журнал. 2002. Т. 9. № 1. Приложение. С. 3–12.
Гусев В. И. Определитель повреждений лесных, декоративных и плодовых деревьев и кустарников . М.: Лесная промышленность, 1984. 472 с.
Земкова Р. И. Стволовые вредители темнохвойных лесов Западного Саяна . Красноярск: ИЛиД СО АН СССР, 1965. 123 с.
Исаев А. С. Динамика численности большого черного усача и меры борьбы с ним в пихтовых лесах Красноярского края // Проблемы защиты таежных лесов. Красноярск, 1971. С. 61–64.
Исаев А. С., Киселев В. В., Ветрова В. П. Роль дополнительного питания большого черного хвойного усача при его взаимодействии с кормовым растением // Консортивные связи дерева и дендрофильных насекомых. Новосибирск: Наука, 1982. С. 19–27.
Исаев А. С., Рожков А. С., Киселев В. В. Черный пихтовый усач Monochamus urussovi (Fisch.) . Новосибирск: Наука, 1988. 396 с.
Кондаков Ю. П., Кнор И. Б., Петренко Е. С. Эколого-хозяйственные группировки лесных насекомых байкальского бассейна // Фауна лесов бассейна озера Байкал. Новосибирск: Наука, 1979. С. 44–77.
Коломиец Н. Г. Итоги изучения вредителей кедра и задачи борьбы с ними // Проблемы кедра / Тр. по лесному хозяйству Сибири. Вып. 6. 1960. С. 175–183.
Криволуцкая Г. О. Скрытостволовые вредители в темнохвойных лесах Западной Сибири . М.; Л.: Наука, 1965. 129 с.
Никитский Н. Б., Ижевский С. С. Жуки-ксилофаги – вредители древесных растений России . М.: Лесная промышленность, 2005. 120 с.
Прозоров С. С. Большой черный усач Monochamus urussovi Fisch. на пихте сибирской // Труды Сибирского лесотехнического института. 1958. Сб. 21. Вып. 2. С. 15–121.
Петренко Е. С. Насекомые-вредители лесов Якутии . М.: Наука, 1965. 167 с.
Савина Л. Н. Новейшая история лесов Западного Саяна (по данным спорово-пыльцевого анализа почв) . Новосибирск: Наука, 1976. 158 с.
Савина Л. Н. Таежные леса Северной Азии в голоцене . Новосибирск: Наука, 1986. 191 с.
Шабалина О. М., Разнобарский В. Г., Гродницкий Д. Л. Живой напочвенный покров и подрост в таежных шелкопрядниках // Сибирский экологический журнал. 2002. Т. 9. № 1. Приложение. С. 23–32.
Amman G. Characteristics of mountain pine beetles reared in four pine hosts // Environmental Entomology. 1982. Vol. 11. № 3. P. 590–593.
Nylin S., Janz N. Host plant preferences in the comma butterfly (Polygonia c-album): Do parents and offspring agree? // Ecoscience. 1996. Vol. 3. № 3. P. 285–289.
Благодарности
Авторы искренне признательны В. П. Ветровой (ИЛ СО РАН, Красноярск) за всестороннюю поддержку работы.

 © 2011 - 2026
© 2011 - 2026