



Введение
Ареал певчего дрозда Turdus philomelos охватывает территорию от Испании и Ирана до 70° с. ш. и от Ирландии до Байкала (Tomiałojć, 1997), несколько варьируя в соответствии с изменением характера растительности и наличием труднопреодолимых преград к расселению (Tomiałojć, 1992). При этом более 75 % певчих дроздов мировой фауны гнездятся и зимуют в пределах Европейского континента (Harrison, 2003), при этом на значительной части территории, за исключением Великобритании и Юго-Западной Европы, обитают представители номинативного подвида. Таким образом, территория Российской Федерации дает возможность охватить как центральные участки ареала гнездования, так и его периферию. Значительная протяженность ареала позволяет использовать представителей вида в качестве модельных объектов изучения начальных стадий адаптивных изменений, включая этологическую адаптацию и, в частности, географическую специфику гнездостроительного поведения.
Наряду с этим актуальность исследования усиливают локально наблюдаемые изменения численности представителей вида. Так, на протяжении последних 25 лет в Великобритании отмечено 50 % сокращение численности певчего дрозда по причинам, в основном связанным с антропогенным преобразованием гнездопригодных участков (Gregory et al., 2002). В целом на территории Европы популяционный индекс певчего дрозда классифицируют как moderate decline – умеренно сокращающийся (Song Thrush…, 2016), что привлекает внимание орнитологов к представителям вида и обостряет актуальность подобных исследований.
Естественные различия условий обитания при перемещении в широтном и долготном направлениях согласно существующим концепциям, отражающим представления об адаптивном изменении признаков в географическом аспекте (Mayr, 1956), предполагают реакцию в виде адаптации для успешного обитания в каждой конкретной точке ареала. Проявление адаптации следует ожидать в соответствии с одной из основных функций гнезда – создание оптимального микроклимата для развития потомства в условиях возможных проявлений факторов окружающей среды в районе гнездования (Mainwaring et al., 2004b). При этом возможность проследить характер адаптивных изменений на уровне теплоизоляционных характеристик построек появилась лишь в последние годы благодаря созданию высокочувствительной тепловизионной техники.
Цель нашей работы состояла в сравнении адаптивных характеристик гнезд, определяющих эффективность теплоизоляции. Для достижения цели проведен сравнительный анализ пространственной организации и показателей естественной и безводной массы гнезд в разных точках гнездового ареала. Сопоставлены тепловизионные изображения распределения тепловых полей по поверхности гнезд после прогрева лотка сферическим термостатом в полевых условиях для определения показателей, определяющих эффективность термоизоляции построек, и прямой оценки интенсивности теплопотерь через поверхности гнезда.
Материалы
Материалы исследований получены в ходе полевого сезона 2015 г. в Карелии, на территории Государственного природного заповедника «Костомукшский» (64º27’ N, 30º17’ E), и в 2016 г. в Мордовии, на территории национального парка «Смольный» (54º44’ N, 45º30’ E).
В местах исследований мы проводили массовый поиск гнезд певчего дрозда. Для проведения измерений из числа всех найденных построек мы отбирали лишь гнезда без видимых повреждений, относительно однотипно расположенные на деревьях, на высоте от 1,5 до 2,5 м, для компенсации влияния характера гнездового субстрата. Следует подчеркнуть, что в данной работе мы не обсуждаем вопрос выбора гнездовых субстратов, которые, несомненно, могут повлиять на теплоизоляционные качества примыкающих к ним участков гнезд. Учитывая высокую пластичность певчего дрозда в выборе гнездового субстрата, в том числе и непосредственно в пределах районов наших исследований (Яковлева, Хохлова, 2008; Хохлова, 2011), на данном этапе исследований участки плотного соприкосновения с гнездовым субстратом исключали из анализа распределения тепловых полей.
При проведении сравнительного анализа изымали гнезда, разоренные или брошенные на этапе кладки или первых дней жизни птенцов, с целью исключения влияния естественного изменения характеристик построек при росте птенцов и эксплуатации гнезд, а также по причине того, что работа в обоих регионах проводилась в пределах особо охраняемых природных территорий. При этом сбор гнезд проводили в начале сезона, чтобы избежать гнезд второго цикла и по возможности не включать в анализ постройки повторного гнездования, поскольку зачастую более поздние гнезда характеризуются более низким качеством и могут исказить результаты сравнительного анализа. С учетом всех приведенных выше условий отбора материала в Карелии было собрано и обработано 11 гнезд, в Мордовии – 9 построек. При этом на разных этапах анализа использованы данные только по гнездам, полностью соответствующим критерию однородности значимых признаков (N в каждом случае указано в иллюстрациях к материалам). В анализ распределения тепловых полей включены 77 тепловизионных изображений, формирующих панорамную термограмму всей поверхности каждого гнезда.
Методы
Измерение гнезд включало основные промеры: наружный и внутренний диаметр, высоту и глубину гнезд, и было проведено с помощью металлической линейки с точностью до 0.5 см (измерение с более высокой точностью считаем нецелесообразным ввиду естественной ассиметричности гнезд и отсутствия четкой границы в местах скругления края постройки). Крайние точки измерений отмечали по условной линии, в соответствии с которой проходит скругление более 80 % элементов материала, образующих стенку гнезда. Отдельные элементы, выступающие за пределы основного массива материала, не учитывали. Объем гнезд вычисляли с использованием формулы, выведенной на основе уравнений объема сфероида, тора и цилиндра, производя соответствующие форме гнезда усечения элементов (Simonov, Matantseva, 2015).
После изъятия гнезда взвешивали при естественном уровне увлажнения и после высушивания в низкотемпературной печи до момента стабилизации массы образца для приведения гнезд в равные условия увлажнения материала с целью повышения репрезентативности сравнительного анализа массы построек и в последующем – их плотности.
Непосредственно после изъятия гнезд проводили количественное измерение термоизоляционных характеристик. Для этого гнездо устанавливали на горизонтальную площадку и помещали в чашу гнезда разработанный нами сферический термостат, представляющий собой стальную сферу, заполненную теплоносителем (сплав Розе) с лампой накаливания в центре сферы и термочувствительным элементом на поверхности сферы (Симонов, Матанцева, 2015). Время прогрева каждого гнезда певчего дрозда составляло 20 минут, температура сферического термостата 38 ± 0.5 °C.
По окончании прогрева проводили съемку гнезд с помощью тепловизора Testo 875i (рис. 1) с 4 позиций со смещением относительно оси гнезда на 90° с целью получения картины распределения тепловых полей по поверхности гнезд (рис. 2). Съемку проводили в горизонтальной плоскости с фиксированного и равного расстояния от оси гнезда до объективов тепловизора.
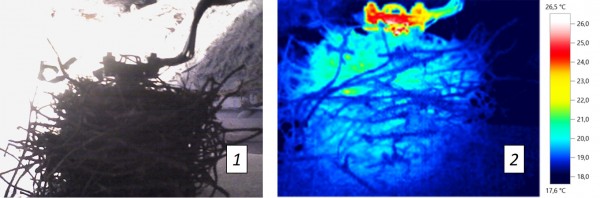
Рис. 1. Фотография гнезда певчего дрозда со сферическим термостатом внутри: 1 – видимый спектр, 2 – инфракрасное изображение
Fig. 1. Photography of a Song Thrush’s nest with the spheroid thermostat inside: 1 – visible-light spectrum, 2 – infrared image
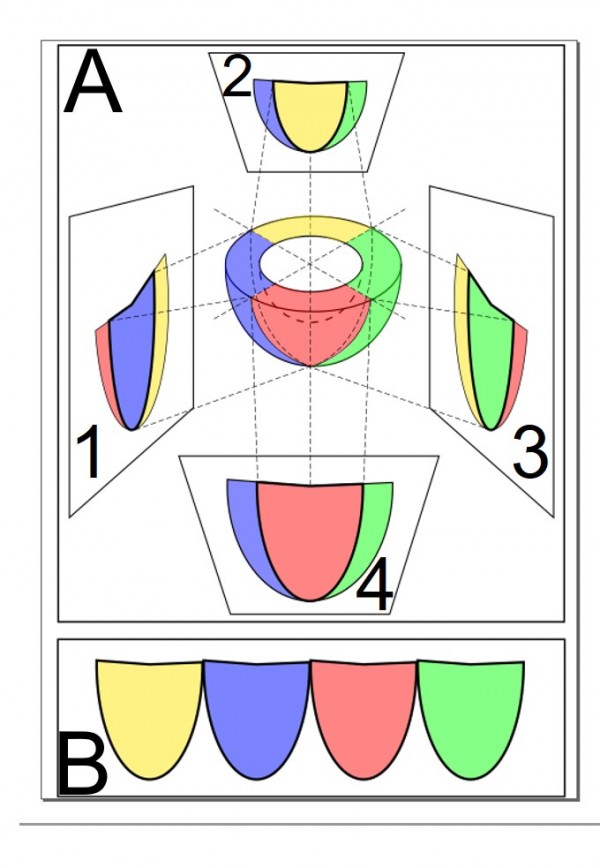
Рис. 2. A: 1–4 – четыре снимка гнезда со смещением 90° относительно оси; B – развертка объемной фигуры в плоскость
Fig. 2. A: 1–4 – four pictures of a nest according to the cardinal directions; B – ‘unrolling’ pictures onto the plane surface
Проведение тепловизионных исследований осуществляли непосредственно в полевых условиях, что гарантировало минимальное воздействие на функциональное состояние построек процесса транспортировки. По естественным причинам условия съемки отдельных построек варьировали. Параметры окружающей среды при проведении съемки контролировали с помощью цифрового комплексного измерительного прибора Testo 410-2, определяющего скорость ветра, температуру и относительную влажность воздуха с точностью 0.1 м/с, 0.1 °C, 0.1 % rH соответственно. При этом в целом при проведении измерений выбирали периоды с относительно однородными условиями окружающей среды.
В основе сравнительного анализа изучаемых параметров гнезд использовали непараметрический аналог двухвыборочного критерия для независимых выборок – U-критерий Манна – Уитни. Связь между наблюдаемыми величинами оценивали с применением непараметрического корреляционного анализа (коэффициент ранговой корреляции Спирмена). Данный тип анализа был выбран по причине его независимости от формы распределения и низкой чувствительности к выбросам. Уровень значимости при проверке статистических гипотез составил 5 %. Статистический анализ проведен с помощью приложения MaxStat® Lite 3.06.
Результаты
На первом этапе сравнения теплоизоляционных качеств гнезд певчего дрозда, построенных в условиях разных широт, были сопоставлены их массовые и габаритные характеристики (табл. 1). В качестве основного параметра, определяющего эффективность термоизоляции, определена плотность гнездовых построек, рассчитываемая на основании измерения гнезд и определения их сухой массы. Согласно полученным данным (рис. 3), плотность гнезд певчего дрозда на более южном участке исследований выше, чем вблизи северной границы ареала (тест Манна – Уитни, U = 68.5, p = 0.01, N1 = 11, N2 = 7).
Таблица 1. Основные характеристики гнезд
| Северные гнезда (N = 9) | Южные гнезда (N = 8) | |||||
| Медиана | Минимум | Максимум | Медиана | Минимум | Максимум | |
| Наружный диаметр, см | 16.5 | 13 | 19 | 15 | 13 | 19 |
| Внутренний диаметр, см | 9 | 8.5 | 11 | 8.5 | 8 | 10 |
| Высота, см | 12.5 | 11 | 15 | 9.5 | 8.5 | 12 |
| Глубина, см | 7 | 6 | 7.5 | 6.5 | 5.5 | 7 |
| Сухая масса, г | 82.5563 | 42.1407 | 109.2366 | 80.11 | 55.82 | 85.71 |
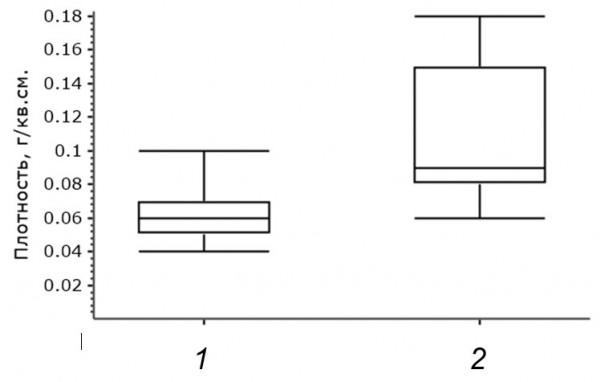
Рис. 3. Плотность гнезд: 1 – северные гнезда, 2 – южные гнезда
Fig. 3. Nest density: 1 – northern nests, 2 – southern nests
При этом географически обусловленных различий в массе в условиях естественного увлажнения и после обезвоживания построек в сушильном шкафу нами не обнаружено (см. табл. 1; тест Манна – Уитни, p во всех случаях больше 0.05). Таким образом, различия в плотности построек должны быть связаны с объемом гнезд. Действительно, объем гнезд, изъятых в Карелии, больше, чем объем гнезд, измеренных в Мордовии (тест Манна – Уитни, U = 71.5, p = 0.01). Причем внутренние промеры и внешний диаметр гнезд статистически не различаются (см. табл. 1; тест Манна – Уитни, p во всех случаях больше 0.05). Причина различий, наблюдаемых в плотности и объеме построек, заключается, очевидно, в том, что гнезда, построенные в Карелии, выше, чем гнезда, построенные южнее (см. рис. 4; тест Манна – Уитни, U = 77.5, p = 0.001).
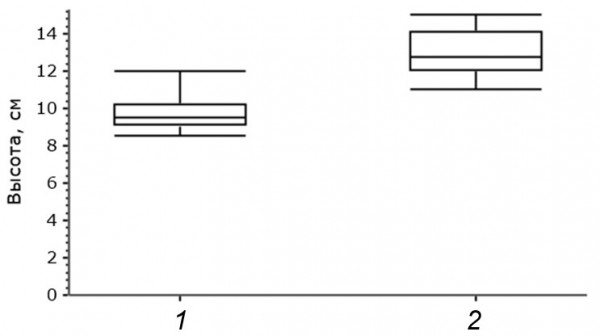
Рис. 4. Высота гнезд: 1 – северные гнезда, 2 – южные гнезда
Fig. 4. Nest height: 1 – northern nests, 2 – southern nests
Для более полной характеристики теплоизоляционных качеств исследуемых гнезд были сопоставлены серии тепловизионных изображений, характеризующие распределение тепловых полей по всей поверхности отдельных построек, полученные при прогревании внутренней поверхности гнезд сферическим термостатом. Комплекс параметров, непосредственно характеризующих распределение тепловых полей по поверхности гнезда, во всех случаях свидетельствует о лучших термоизоляционных свойствах гнезд, найденных на севере, в сравнении с гнездами, построенными на юге (табл. 2).
Таблица 2. Результаты сравнения параметров распределения тепловых полей по поверхности гнезд, тест Манна – Уитни
| Прирост температуры* | Медиана на севере, °С
(N = 11) |
Медиана на юге, °С
(N = 7) |
U | p |
| Минимальный | 0.5 | 2.0 | 73.0 | 0.002 |
| Максимального по площади эквитермального участка | 2.0 | 3.0 | 67.0 | 0.010 |
| Максимальный | 5.5 | 7.0 | 75.5 | 0.001 |
Примечание. * – градиент температуры в данном случае является разностью между температурой прогретого участка поверхности гнезда и температурой окружающих непрогретых поверхностей.
Сопоставление различных параметров, имеющих отношение к геометрии и термоизоляции гнезд, позволило определить, что более плотные гнезда характеризуются меньшим показателем наибольшей площади эквитермальных участков наружной поверхности постройки (ранговая корреляция Спирмена, r = – 0.719, p = 0.008). Как и следовало ожидать, теплоизоляционные характеристики менее плотных гнезд превышают таковые более плотных построек. Максимальные эквитермальные участки рыхлых построек больше, прирост температуры на этих участках ниже. Распределение тепловых полей по поверхности плотных построек более мозаичное, прирост температуры выше.
Обсуждение
Приступая к анализу отдельных рассматриваемых проявлений адаптации к условиям, сопровождающим репродуктивный период птиц, следует отметить, что в задачи исследования не входил анализ этологических адаптаций в виде изменения аттентивности (интенсивности обогрева содержимого гнезда), предшествующих проявлению физических адаптаций в виде изменения параметров построек.
Результаты, полученные на основе наших данных, свидетельствуют о различиях между физическими характеристиками гнезд двух рассматриваемых территорий. Географически обусловленные различия построек животных были неоднократно отмечены ранее (King et al., 1964; Deeming et al., 2012; Mainwaring et al., 2012 и др.). И если различия в целом, скорее всего, являются выражением географической адаптации к конкретным условиям среды, то причины каждого из них в отдельности могут быть различными.
В частности, относительно недавнее исследование гнезд черного дрозда Turdus merula в Великобритании показывает существенное увеличение всех основных размеров гнезд при движении в широтном направлении на 7 градусов (Mainwaring et al., 2014a). Различие между выбранными пунктами в нашем исследовании в широтном направлении превышало 10 градусов, дополнительно включая разницу в долготном направлении порядка 15 градусов. Однако, несмотря на значительное удаление пунктов исследования друг от друга, практически все габаритные характеристики построек оказались одинаковыми. Исключение составила высота гнезда за счет разницы в толщине дна постройки – на севере гнезда певчего дрозда имели более толстое дно. Преимущество увеличения толщины термоизоляционного материала очевидно – таким образом сокращаются энергопотери в любой термоизоляционной конструкции. Возможно, наблюдаемое соотношение параметров построек может быть обусловлено, помимо прочего, особенностями сезонов исследований, поскольку полевые работы в рассматриваемых районах проводились в разные сезоны. Однако фенологические условия проведения исследований существенно не различались, а строгий отбор гнездовых построек во многом компенсировал риск попадания в выборку поздних гнезд.
При этом возникает вопрос: за счет чего происходит увеличение толщины материала при отсутствии значимых различий в массе гнездовой постройки? По-видимому, причина этого заключается в характере доступного материала и его пространственной организации. Основание гнезд певчего дрозда в Карелии в местах проведения исследований построено с использованием тонких еловых веточек, в то время как гнезда в Мордовии построены с использованием тонких веточек сосны. Причина этого заключается в низком количестве ели в районе исследований в Мордовии. Характер ветвления фрагментов ели и сосны, используемых при постройке гнезда, значительно различается. Фрагменты веточек сосны в постройках практически лишены боковых отростков. Веточки ели, напротив, имеют большое число боковых отростков, образуя объемную пространственную структуру. Таким образом, укладка равной массы материала без дополнительного уплотнения образует массив различной толщины и плотности.
Последнее особенно важно в связи с тем, что наравне с основанием постройки большое значение для успешного выведения потомства должны иметь физические параметры стенок конструкций. В связи с особенностями гнезд дроздов, состоящих из внутренней плотной чаши и внешней более рыхлой оболочки, мы отдельно сопоставляли толщину и теплоизоляционные характеристики внутренних плотных чаш, однако никаких географически обусловленных различий в этих параметрах не обнаружено. По-видимому, основная функция внутренней чаши состоит в сохранении формы постройки, защите от дождя и ветра, в то время как основную функцию теплоизоляции выполняет более рыхлый наружный слой. Поскольку внутренний слой не вносит существенного вклада в географически обусловленные различия теплоизоляционных качеств, в данной работе мы не представляем его отдельного обсуждения, и весь дальнейший анализ относится к гнездам целиком.
Согласно полученным данным, достоверных различий общей толщины стенок между гнездами разных районов не отмечено. При этом показатели термоизоляции северных гнезд выше, чем аналогичные параметры гнезд, построенных в более южных условиях. На фоне отсутствия различий в массе конструкций это может свидетельствовать о существовании перераспределения плотности укладки гнездового материала внутри конструкции. Перераспределение плотности материала позволяет увеличить объем воздуха, заключенного между отдельными конструктивными элементами и являющегося основным теплоизолятором построек. Таким образом, согласно предварительным заключениям, сделанным на основе полученных данных, гнезда, построенные на севере, при равных габаритных характеристиках стенок, построены более рыхло (точнее, их наружная часть, исключая плотный лоток) и, как следствие, менее интенсивно пропускают тепловое излучение. Если рассматривать такие постройки как отдельные физические тела, их центр тяжести несколько смещен книзу по сравнению с более компактными гнездами, построенными в более южных условиях. Подобное перераспределение материала и плотности может решать две задачи: приводить к улучшению теплоизоляционных свойств стенок за счет наполнения конструктивных элементов воздухом, а также приводить к снижению теплопотерь через основание гнезда за счет более рыхлой пространственной организации материала основания гнезда и увеличения его массы.
Заключение
Естественные ограничения репродуктивного периода, имеющие особенное значение в жизни мигрирующих животных, являются одним из факторов естественного отбора, приводящего к проявлению и закреплению адаптаций, повышающих успешность размножения. Давно известно, что этапу появления адаптаций на физиологическом уровне предшествует появление этологических адаптаций (Северцов, 1922; Tinbergen, 1953; Промптов, 1956; Hinde, 1970 и др.). Но должен существовать момент, когда этологические адаптации исчерпывают свою эффективность и возникает момент появления физических адаптаций. В этом отношении гнезда подставляют особый интерес, поскольку в определенном смысле занимают пограничное положение между двумя уровнями адаптаций, т. е. изменяется лишь гнездостроительное поведение, но не организм животного, а внешнее проявление поведенческих актов представляет собой законченную объемную структуру, обладающую уникальными свойствами, определяющими существование представителей вида в конкретных условиях. Характеристики гнездовой постройки должны соответствовать всем возможным климатическим явлениям в районе гнездования, причем технологические решения, определяющие адаптивные признаки гнезд, практически неизбежно должны быть свойством локального населения птиц и поддерживаться филопатрическими связями.
Предварительные итоги сравнительного анализа гнезд певчего дрозда свидетельствуют об адаптивном изменении свойств гнездовых построек. Гнезда, построенные вблизи северной границы ареала, имеют более рыхлую конструкцию и лучшие термоизоляционные качества конструктивных элементов; эквитермальные тепловые поля локализованы и структурированы, обеспечивают относительно равномерную теплоизоляцию. При этом теплоизоляционная эффективность более южных гнезд ниже, эквитермальные тепловые поля на поверхности построек распределены более мозаично, что не компенсирует теплопотерь всей плоскости. Дальнейшее накопление материалов по вопросам географической изменчивости габаритных и теплоизоляционных характеристик гнездовых построек позволит провести более детальный сравнительный анализ адаптивных качеств гнезд.
Библиография
Северцов А. Н. Общие вопросы эволюции. Эволюция и психика // Собрание сочинений. М.: Изд-во АН СССР, 1922. Т. 3. 530 с.
Симонов С. А., Матанцева М. В. Анализ теплоизоляционных качеств гнезд птиц с использованием тепловизионной аппаратуры // Современные проблемы науки и образования. 2015. № 5. URL: http://www.science-education.ru/128-22166 (дата обращения: 20.07.2016).
Промптов А. Н. Очерки по проблеме биологической адаптации поведения воробьиных птиц . М.; Л.: Изд-во АН СССР, 1956. 311 с.
Хохлова Т. Ю. Популяционные адаптации к условиям севера таежной зоны близкородственных видов птиц с полицикличным размножением (на примере дроздов рода Turdus): Дис. ... канд. биол. наук . СПб., 2011. 357 с.
Яковлева М. В., Хохлова Т. Ю. О влиянии хищников на продуктивность гнездования белобровика (Turdus iliacus) и певчего дрозда (T. philomelos) в южной Карелии // Труды Государственного природного заповедника «Кивач». Вып. 4. Петрозаводск: Изд-во ПетрГУ, 2008. С. 135–144.
Deeming D. C., Mainwaring M. C., Hartley I. R., Reynolds S. J. Local temperature and not latitude determines the design of Blue Tit and Great Tit nests // Avian Biology Research. 2012. Vol. 5. №4. P. 203–208. DOI: 10.3184/175815512X13528874959581.
Gregory R. D., Wilkinson N. I., Noble D. G., Robinson J. A., Brown A. F., Hughes J., Proctor D. A., GIibbons D. W., Galbraith C.A. The population status of birds in the United Kingdom, Channel Islands and Isle of Man: an analysis of conservation concern 2002-2007 // British Birds. 2002. № 95. P. 410–450.
Harrison G. Song thrush Turdus philomelos // Warwickshire, Coventry and Solihull Local Biodiversity Action Plan. 2003. URL: http://www.warwickshire.gov.uk/biodiversity (дата обращения: 20.07.2016).
Hinde R. A. Animal behaviour. New York: McGraw-Hill, (1966) 1970. 876 p.
King J. A., Maas D., Weisman R. G. Geographic variation in nest size among species of Peromyscus // Evolution. 1964. Vol. 18. № 2. P. 230–234. URL: http://www.jstor.org/stable/2406395 (дата обращения: 20.07.2016).
Mainwaring M. C., Hartley I. R., Bearhop S., Brulez K., du Feu C. R., Murphy G., Plummer K. E., Webber S. L., Reynolds S. J., Deeming D. C. Latitudinal variation in blue tit and great tit nest characteristics indicates environmental adjustment // Journal of Biogeography. 2012. Vol. 39. Issue 9. P. 1669–1677. DOI: 10.1111/j.1365-2699.2012.02724.x.
Mainwaring M. C., Deeming D. C., Jones C. I., Hartley I. R. Adaptive latitudinal variation in Common Blackbird Turdus merula nest characteristics // Ecology and Evolution. 2014a. Vol. 4. № 6. P. 851–861. DOI:10.1002/ece3.952.
Mainwaring M. C., Hartley I. R., Lambrechts M. M., Deeming D. C. The design and function of birds’ nests // Ecology and Evolution. 2014b. Vol. 20. № 4. P. 3909–3928. DOI:10.1002/ece3.1054.
Mayr E. Geographical character gradients and climatic adaptation // Evolution. 1956. Vol. 10. № 1 P. 105–108. URL: http://www.jstor.org/stable/2406103?origin=JSTOR-pdf.
Simonov S. A., Matantseva M. V. The study of thermal insulation in bird nests: justification of used parameters // Science Almanac. 2015. Vol. 9. № 11. P. 1051–1055. DOI: 10.17117/na.2015.09.1051.
Song Thrush (Turdus philomelos) // EBCC Population index, 1980–2005, Europe. URL: http://www.ebcc.info/index.php?ID=271 (дата обращения: 20.07.2016).
Tinbergen N. Social Behaviour in Animals. London: Methuen, 1953. 150 p.
Tomiałojć L. Colonization of dry habitats by the Song Thrush Turdus philomelos: is the type of nest material an important constraint // Bull. Br. Ornithol. Club. 1992. Vol. 112. P. 27–34.
Tomiałojć L. Song Thrush Turdus Philomelos // The EBCC atlas of European breeding birds: their distribution and abundance / European Bird Census Council; Ed. by W. J. M. Hagemeijer, M. J. Blair et al. London: T & A D Poyser, 1997. P. 548–549.
Благодарности
Мы очень признательны руководству и сотрудникам Федерального государственного бюджетного учреждения «Государственный заповедник "Костомукшский"» и Федерального государственного бюджетного учреждения «Национальный парк "Смольный"» за возможность проведения исследований на охраняемой территории и всестороннюю поддержку при организации и осуществлении полевых исследований. Особую благодарность мы выражаем д. б. н. Т. Ю. Хохловой за знакомство с рукописью статьи и ценные дополнения, а также рецензентам за предложенные рекомендации. Работа выполнена по гранту Президента РФ по государственной поддержке молодых российских ученых – кандидатов наук (проект № MK-3599.2015.4).

 © 2011 - 2026
© 2011 - 2026