



Введение
Заповедная акватория Кандалакшского залива с населяющей ее биотой в течение многих десятилетий интенсивно исследуется как эталон морских экосистем, с акцентом на оценке состояния кормовой базы морских рыб и птиц, в том числе гаги обыкновенной Somateria mollissima (Дорош, 1963; Бианки и др., 1979; Краснов и др., 2009). Данная работа является частью обширного детального изучения явлений и процессов, протекающих в прибрежных морских экосистемах, по программе «Летописи природы Кандалакшского государственного природного заповедника».
Брюхоногий моллюск Littorina littorea L. является одним из массовых видов, обитающих на литорали Белого моря (Рубинчик, 1962; Голиков, Кусакин, 1978; Моллюски Белого моря, 1987). Литторина этого вида редко становилась объектом исследований, ее биология и экология изучены недостаточно. Моллюск играет большую роль в морских экосистемах, т. к. является типичным представителем мелководных бентосных сообществ и важным звеном в их трофической составляющей. В качестве источника питания Littorina littorea использует детрит, фрагменты макроводорослей, реже животную пищу. Продукты жизнедеятельности, в частности пеллеты и слизь, выделяемая железами ноги моллюска, служат источником пищи для других гидробионтов (Бескупская, 1963; Белое море, 1995; Davies, 1999). Все Gastropoda в пищевом рационе гаги составляют 15 % от общего количества употребляемых этими птицами моллюсков. Конкретно на L. littorea приходится 5 % от общих 15 % (Бианки и др., 1979).
Рост – комплексный биологический процесс, являющийся неотъемлемой частью развития организмов и реализующийся в тесной взаимосвязи с условиями окружающей среды, в которых обитает живой объект (Мина, Клевезаль, 1976; Озернюк, Зотин, 2006; Седловская, 2012). Большинство научных работ, связанных с исследованием L. littorea, описывают влияние плотности популяции на скорость роста моллюсков (Kemp, Bertnees, 1984; Goodfriend, 1986; Yamada, Mansour, 1987). Темп роста литторин регулируется, прежде всего, термогалинными условиями обитания.
L. littorea – это стенобатный, эвритермный тепловодный и стенотопный моллюск, обитающий в основном на каменистых грунтах (Федяков, 1986). Литторина придерживается нижнего горизонта литорали, иногда крупные особи можно встретить в среднем горизонте. Предпочитает температуру воды не ниже 2,5 ˚С, а нерест у этих моллюсков начинается при температуре не ниже 8–10 ˚С (Рубинчик, 1962; Федяков, 1986). Отмечено, что L. littorea отрицательно относится к низким значениям солености воды. У моллюска наблюдается угнетение таких процессов жизнедеятельности, как дыхание, при значении солености в диапазоне от 10 до 15 ‰ (Бергер, 1970). Однако существуют данные, которые свидетельствуют о высокой устойчивости личинок литторин к крайне низким (8 ‰) и крайне высоким (50 ‰) значениям солености (Саранчова и др., 2006).
Непосредственно на литорали в большинстве случаев встречаются только крупные взрослые экземпляры L. littorea (в возрасте от 5 лет). Молодые моллюски (в возрасте до 4 лет) не могут противостоять резко изменяющимся условиям среды, которые воздействуют на все живые организмы, населяющие литораль. Поэтому особи, не достигшие пятилетнего возраста, чаще держатся ниже нуля глубин (Рубинчик, 1962; Бескупская, 1963).
Цель нашей работы – сравнить интенсивность роста брюхоногого моллюска Littorina littorea L. в двух местах его массового обитания в Кандалакшском заливе: о. Ряшков и Порья губа.
Материалы
Материал для исследования собирали в июле 2014 года на базе Кандалакшского государственного заповедника, на островах губы Порья и острове Ряшков (рис. 1).
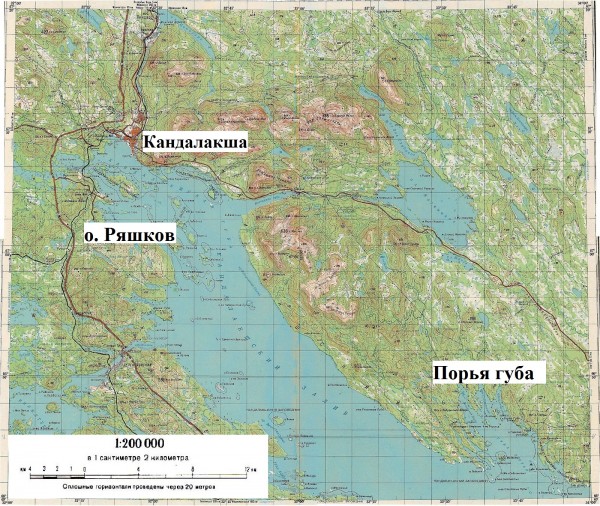
Рис. 1. Карта Кандалакшского залива Белого моря с указанием мест исследований
Fig. 1. Map of the Kandalakshsky gulf of the White Sea showing the locations of research places
Места сбора материала расположены в разных областях залива и имеют различающийся температурно-соленостный режим вод, а также разную степень антропогенного влияния.
Особенностью Порьей губы считается то, что холодное глубинное течение подходит близко к поверхности воды, поэтому здесь наблюдается резко контрастная стратификация температурно-солевого режима в весенне-летне-осенний период. С другой стороны, южная экспозиция губы способствует хорошему прогреванию мелководий материкового склона. Приливно-отливные течения перемешивают прогретую воду мелководий с глубинной холодной водой, что в результате создает резкие колебания гидрологических условий по сравнению с другими районами Кандалакшского залива (Шкляревич, 1998; Брязгин, 2005). Глубина Порьей губы от 20 до 80 м, максимум обнаружен в центре губы – 120 м (Шмелинг и др., 1964). Нижний горизонт литорали представлен каменистой грядой или мягкими илисто-песчаными фракциями и населен Ascophyllum nodosum + Fucus vesiculosus + Mytilus edulis.
Остров Ряшков – один из крупных островов вершины залива, располагающийся в более мелководном районе по сравнению с Порьей губой. Вокруг острова находится отмель с глубинами менее 20 м, на которой располагаются сплошные поселения мидий Mytilus edulis – мидиевые банки. Нижний горизонт литорали представлен мягкими илисто-песчаными фракциями с разбросанными глыбами, населенными Ascophyllum nodosum + Fucus vesiculosus + Mytilus edulis.
В ходе работы собрано 199 экземпляров Littorina littorea L., в том числе в Порьей губе – 61, на о. Ряшков – 138 особей. При этом в рассчетах использованы значения высот раковин 115 животных: в Порьей губе – 57, на о. Ряшков – 58.
Методы
В обоих местах исследования от уреза воды в момент сбора проб до нуля глубин десятикратно определены значения солености и температуры морской воды, а затем вычислены их средние арифметические. Для определения температуры был использован термометр в металлическом чехле со стаканчиком, а для измерения плотности – ареометр. В дальнейшем значения плотности воды переводились в соленость по океанологическим таблицам (Зубов, 1957). Для иллюстрации того, что исследуемый объект чувствителен к низкой солености, в вершине Кандалакшского залива определены ее значения в некоторых теоретически возможных местах обитания этого моллюска, а также установлено присутствие или отсутствие литорины в них.
Сбор моллюсков в обоих местах их обитания осуществлялся во время полного отлива, т. к. только в таких условиях есть возможность работать на нижнем горизонте литорали. Крупные Littorina littorea, как правило, держатся во время отлива на обсохших камнях, водорослях или грунте, а более мелкие экземпляры пережидают временные неблагоприятные безводные условия под водорослями или в щелях между камнями. Собирались литторины непосредственно руками, каждая особь в отдельности, т. к. на литорали эти моллюски находятся разрозненно и легко отделяются от субстрата. Во время сбора литорины помещались в небольшие пластиковые контейнеры с морской водой, чтобы животные при транспортировке к месту лабораторной обработки не потеряли в весе и не погибли. При этом пробы содержались в прохладном месте. Временной интервал между работой на литорали и началом работы в лаборатории не превышал трех часов.
Обработка материала сводилась к измерению у каждого моллюска высоты раковины с помощью штангенциркуля (точность 0.1 мм) (рис. 2а). Кроме того, определялся возраст каждого моллюска. Методика определения возраста основана на подсчете годичных колец, или отметок на поверхности раковины, которые образуются каждый год во время зимней приостановки роста (рис. 2б) (Алимов и др., 1990).

Рис. 2. Измерение высоты раковины (а); раковина Littorina littorea L. с годовыми отметками (б)
Fig. 2. Measurement of shell height (а); shell of Littorina littorea L. with annual marks (b)
Математическая обработка материала сводилась к построению линий и уравнений регрессии с использованием пакета анализа данных в среде MS Excel. При этом была использована криволинейная регрессия. Нелинейная форма связи характеризуется следующей особенностью: равномерное изменение одного признака сопровождается неравномерным изменением значения другого признака (Коросов, Горбач, 2010; Ивантер, Коросов, 2011). Работа с криволинейной регрессией заключалась в преобразовании (логарифмировании) исходных эмпирических данных, на основе которых был осуществлен линейный регрессионный анализ, произведена оценка значимости коэффициентов регрессии (критерий Стьюдента) и адекватности выбранной модели (критерий Фишера). После обратного преобразования коэффициентов линейной регрессии были построены уравнения криволинейной регрессии.
Линии регрессии построены на основе значений высот раковин моллюсков в возрастном диапазоне от 6 до 12 лет, т. к. представители данных возрастных группировок присутствуют в обеих выборках.
Методика сравнения коэффициентов регрессии заключалась в оценке гипотезы об одинаковом положении линий регрессии относительно осей координат с помощью критерия Стьюдента. При этом были определены суммы квадратов отклонений значений независимой переменной от своих средних с помощью статистической функции в среде Excel, рассчитаны значения остаточной дисперсии для каждой выборки и общей остаточной дисперсии, а также значение обобщенной ошибки коэффициентов регрессии. Оценка значимости отличий коэффициентов производилась по формуле:
Т = а1 - а2 / ma1,2,
где Т – значение критерия Стьюдента, а1 - а2 – разница коэффициентов уравнений регрессии, ma1,2 – обобщенная ошибка коэффициентов регрессии (Ивантер, Коросов, 2011). Обобщенная ошибка вычисляется по формуле:
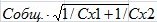 ,
,
где Собщ. – общая остаточная дисперсия, Сх1 и Сх2 – сумма квадратов отклонений х от своих средних. Общая остаточная дисперсия рассчитывается посредством извлечения квадратного корня из выражения: (n1 - 2) · С2ост1 + (n2 - 2) · С2ост2 / n1 + n2 - 4, где n – объем выборки, а С2ост1 и С2ост2 – величина остаточной дисперсии для каждой выборки.
Кроме того, для каждой линии регрессии определен доверительный интервал (область, в которой с вероятностью 95 % находится истинная линия регрессии), интервал прогноза (область, в пределах которой с вероятностью 95 % ожидаются новые значения вариант) и выполнено прогнозирование значений зависимой переменной (у) путем подстановки в уравнение регрессии соответствующих значений независимой переменной (х). Границы доверительного интервала и интервала прогноза рассчитывались по формулам, предложенным в учебном пособии Э. В. Ивантера и А. В. Коросова (2011). Формула расчета границ доверительного интервала линии регрессии:
уi ± T · myi,
где 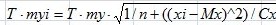 , уi – значение, рассчитанное по уравнению регрессии для xi, T – нормированное отклонение (критерий Стьюдента) при заданном уровне значимости (a) 0.05 и числе степеней свободы (df) n - 1, myi – ошибка линии регрессии, n – объем выборки, (xi – Mx)^2 – мера отклонения значения xi от средней Мх, Сх – сумма квадрата отклонений всех значений х от своей средней Мх .
, уi – значение, рассчитанное по уравнению регрессии для xi, T – нормированное отклонение (критерий Стьюдента) при заданном уровне значимости (a) 0.05 и числе степеней свободы (df) n - 1, myi – ошибка линии регрессии, n – объем выборки, (xi – Mx)^2 – мера отклонения значения xi от средней Мх, Сх – сумма квадрата отклонений всех значений х от своей средней Мх .
Границы интервала прогноза найдены по формуле:
yi ± T · Syi,
где 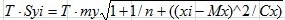 , Syi – расчетное значение признака у.
, Syi – расчетное значение признака у.
Оценка адекватности модели исходным эмпирическим данным произведена с помощью дисперсионного анализа.
Расчет средних арифметических значений размеров высоты раковины моллюсков произведен для тех возрастных рядов, количество особей в которых ≥ 4. Достоверность отличий между значениями средних размеров раковин моллюсков из разных мест обитания оценивалась с помощью критерия Стьюдента.
Результаты
Температурные и соленостные условия – фундаментальные факторы среды, регулирующие темпы роста исследуемого объекта. Среднее значение солености воды для о. Ряшков составило 17.76 ‰, температуры – 17.11 ˚С. Для вод Порьей губы среднее значение солености – 20.64 ‰, а температуры – 14.92 ˚С. В кутовой части залива зафиксированные нами значения солености находятся в пределах от 2 до 12 ‰: юго-западный мыс эстуария реки Нива – 1.36 ‰; о. Овечий – 10.28 ‰; о. Малый – 10.94 ‰; о. Еловый – 11.20 ‰. Поэтому Littorina littorea не встречается на литорали в этом районе, где морские поверхностные воды периодически подвергаются сильному опреснению.
В обоих местах исследования были обнаружены моллюски в возрасте от 3 до 12 лет. На о. Ряшков найдена одна двухлетняя особь и в целом выявлено преобладание моллюсков в возрасте от 3 до 6 лет. Сравнение средних значений высоты раковины L. littorea показало, что шести-, девяти- и десятилетние особи из Порьей губы крупнее таковых с о. Ряшков (табл. 1).
Таблица 1. Средние значения высоты раковины Littorina littorea L. в каждом возрастном ряду и их статистическая оценка
| r | о. Ряшков | Порья губа | df | Тэ | Тt (0.05) | ||
| n | M±m | n | M±m | ||||
| 2 | 1 | 11.00 | 0 | ||||
| 3 | 32 | 11.30±0.21 | 1 | 14.40 | |||
| 4 | 30 | 13.38±0.26 | 2 | 13.90 17.10 | |||
| 5 | 18 | 16.39±0.38 | 1 | 22.40 | |||
| 6 | 11 | 18.25±0.33 | 4 | 20.68±1.04 | 13 | 2.22711 | 2.16037 |
| 7 | 10 | 20.64±0.53 | 13 | 21.55±0.43 | 21 | 1.33334 | 2.07961 |
| 8 | 6 | 23.23±0.69 | 15 | 24.09±0.38 | 19 | 1.09176 | 2.09302 |
| 9 | 10 | 24.23±0.37 | 10 | 26.26±0.40 | 18 | 3.72555 | 2.10092 |
| 10 | 12 | 24.65±0.31 | 8 | 27.24±0.55 | 18 | 4.10233 | 2.10092 |
| 11 | 6 | 26.92±0.69 | 4 | 27.78±0.84 | 8 | 0.79112 | 2.306 |
| 12 | 2 | 29.10 29.30 | 3 | 28.70 30.10 36.40 | |||
Примечание: r – возраст; n – количество особей в возрастном ряду; M±m – среднее значение высоты раковины в возрастном ряду с доверительным интервалом; df – число степеней свободы; Тэ – эмпирическое значение критерия Стьюдента; Tt – теоретическое значение критерия Стьюдента (в скобках заданный уровень значимости).
Зависимость размеров особей от возраста в разных местообитаниях изучали с помощью регресионного анализа (рис. 3).
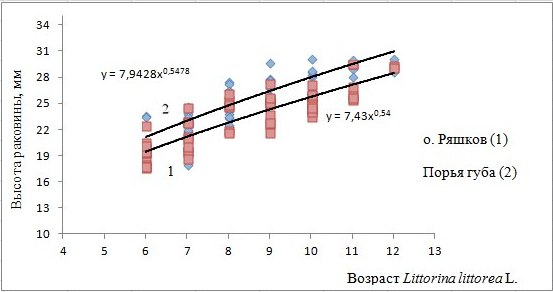
Рис. 3. Модели, описывающие зависимость высоты раковины от возраста моллюска: y = 7.43x0.54 для моллюсков с о. Ряшков (1), y = 7.9428x0.5478 для моллюсков Порьей губы (2)
Fig. 3. Models describing the dependence of the height of shell on its age: y = 7.43x0.54 for Littorina littorea from the island Ryachkov (1), y = 7.9428x0.5478 for mollusks from the Porya Bay (2)
Выбранная модель (y = b × xa) адекватно описывает эмпирические данные, это говорит о том, что зависимость между признаками есть. Расчетное значение критерия Фишера для выборки из Порьей губы составляет 117.48 и превышает теоретическое 4.02. Судя по коэффициенту детерминации (0.68) вариация значения высоты раковины моллюсков на 68 % объясняется вариацией независимой переменной – возраста. Для выборки с о. Ряшков теоретическое значение критерия (4.013) меньше расчетного (173.66), коэффициент детерминации составил 0.75.
Расчетные значения критерия Стьюдента для коэффициентов а и b в уравнении 1 (о. Ряшков) составили 13.17 и 22.44 соответственно при теоретическом значении критерия 2.003. Оба коэффициента уравнения 2 (Порья губа) также значимо отличаются от нуля: вычисленное значение критерия для коэффициента а составило 10.78, для b – 19.02. Табличное значение критерия Стьюдента при уровне значимости 0.05 равняется 2.004.
Ключевым моментом работы являлось сравнение коэффициентов регрессии. Согласно формулам, получили расчетные значения: Собщ. = 0.304, ma1,2 = 0.0164, T = 0.47, Tt (0.05;111) = 1.98.
Коэффициенты регрессионных уравнений достоверно не отличаются.
В результате точечного прогноза значений уi вычислен доверительный интервал и интервал прогноза для среднего значения высоты раковины в каждом значении х (возраста моллюска) (табл. 2).
Таблица 2. Доверительный интервал и интервал прогноза теоретического значения высоты раковины моллюска для каждого возраста
| Возраст | о. Ряшков | Порья губа | ||
| Зона доверительного интервала (T·myi) | Зона интервала прогноза (T·Syi) | Зона доверительного интервала (T·myi) | Зона интервала прогноза (T·Syi) | |
| 6 | 19.55±0.35 | 19.55±1.65 | 21.19±0.52 | 21.19±2.17 |
| 7 | 21.25±0.27 | 21.25±1.63 | 23.06±0.38 | 23.06±2.14 |
| 8 | 22.84±0.21 | 22.84±1.63 | 24.81±0.29 | 24.81±2.12 |
| 9 | 24.34±0.22 | 24.34±1.63 | 26.47±0.29 | 26.47±2.12 |
| 10 | 25.76±0.28 | 25.76±1.64 | 28.04±0.38 | 28.04±2.14 |
| 11 | 27.12±0.36 | 27.12±1.65 | 29.54±0.52 | 29.54±2.17 |
| 12 | 28.43±0.46 | 28.43±1.68 | 30.98±0.68 | 30.98±2.21 |
Таким образом, среднее значение высоты раковины моллюсков с о. Ряшков, например, в шестилетнем возрасте с надежностью 95 % будет находиться в интервале от 19.2 до 19.9 мм, а при условии увеличения числа наблюдений прогнозируемое значение высоты раковины будет находиться в интервале от 17.9 до 21.2 мм.
Доверительный интервал линии регрессии и интервал прогноза новых значений высоты раковины моллюсков с о. Ряшков и из Порьей губы имеет вид (рис. 4, 5).
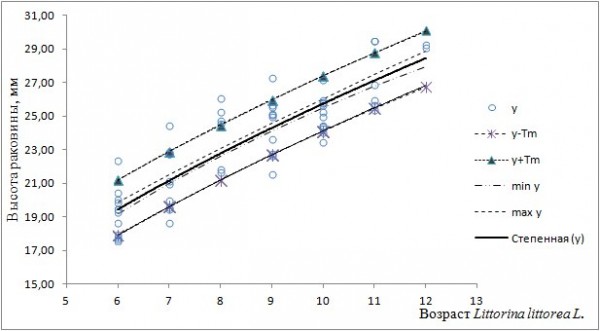
Рис. 4. Линия регрессии (y), ее доверительный интервал (min y; max y) и интервал прогноза (y-Tm; y+Tm) для модели yi = 7.43x0.54 (о. Ряшков)
Fig. 4. The regression line (y), its confidence interval (min y; max y) and forecast interval (y-Tm; y+Tm) for the model yi = 7.43x0.54 (the island Ryachkov)
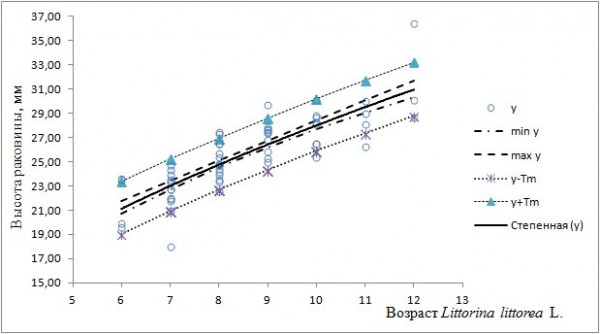
Рис. 5. Линия регрессии (y), ее доверительный интервал (min y; max y) и интервал прогноза (y-Tm; y+Tm) для модели yi = 7.9429x0.5478 (Порья губа)
Fig. 5. The regression line (y), its confidence interval (min y; max y) and forecast interval (y-Tm; y+Tm) for the model yi = 7.9429x0.5478 (the Poria Bay)
Дисперсионный анализ показал, что расчетное значение критерия Фишера для модели yi = 7.43x0.54 (171.21) и yi = 7.9429x0.5478 (120.28) превышает табличное значение 4.013 и 4.02 соответственно.
Исходя из подробного анализа регрессионные модели для обеих выборок адекватно описывают зависимость высоты раковины от возраста моллюска.
Обсуждение
Изучение экологии морских гидробионтов сводится к исследованию таких процессов жизнедеятельности, которые корректируются изменяющимися условиями водной среды. Главным образом на морские литоральные организмы оказывают влияние температура и соленость воды, а также гидродинамическая активность и обеспеченность питанием.
Littorina littorea – пойкилотермный организм, поэтому говорить о том, что температура не оказывает на него влияния, бессмысленно. Как показали наши исследования, на о. Ряшков среднее значение температуры морской воды выше такового в Порьей губе. Известно, что губа Порья характеризуется поднятием глубинных холодных вод, что сказывается на температуре поверхностной воды. В кутовой части залива зафиксированные нами значения солености находятся в пределах от 2 до 12 ‰. Причиной сильного опреснения, прежде всего, является то, что в этом месте в Белое море впадает сразу несколько крупных рек: Нива, Лупче-Савино, Нижняя Лувеньга. Особи Littorina littorea в местах, где было произведено измерение солености воды, обнаружены не были. Остров Ряшков располагается мористее, и воды, омывающие его, имеют усредненное значение солености 17,76 ‰. Тогда как в Порьей губе – 20,64 ‰. Это, с одной стороны, можно объяснить интенсивностью перемешивания поверхностных и глубинных (более соленых) вод в Порьей губе, с другой – близостью расположения губы к Баренцеву морю, в котором воды имеют более высокие значения солености по сравнению с водами Белого моря.
Исходя из вышесказанного, соленость воды является определяющим фактором среды для Littorina littorea, и моллюски из Порьей губы находятся в более благоприятных соленостных условиях.
Исследование показало, что интенсивность роста Littorina littorea в двух местах ее массового обитания в Кандалакшском заливе достоверно не отличается. Статистическая значимость модели, описывающей зависимость высоты раковины моллюска от возраста, подтверждена тремя способами: при доказательстве адекватности модели, отражающей зависимость исходных величин высот раковин от возраста, при оценке значимости коэффициентов уравнений регрессии и при построении доверительных интервалов и интервалов прогнозов с последующей оценкой адекватности теоретической модели.
Если обратиться к рис. 3, видно, что сравниваемый коэффициент уравнения, построенного на основе данных высоты раковины моллюсков из Порьей губы, несколько больше, и расположение кривой роста на графике выше. Как видно из табл. 1, средние значения высоты раковины моллюсков в Порьей губе больше таковых с острова Ряшков. В трех возрастных рядах (6, 9, 10 лет) отличия между средними арифметическими оказались значимы. Исходя из этого можно ожидать, что скорость роста Littorina littorea на островах Порьей губы будет выше. Для того чтобы была возможность представить дополненную картину скорости роста моллюсков в этих двух местах обитания, необходимо в будущем исследовании акцентировать внимание на молодых моллюсках в возрасте от 2 до 5 лет.
На сегодняшний день мы располагаем фактическими данными только по солености и температуре морской воды в местах обитания Littorina littorea. Следует планировать продолжение и совершенствование исследований.
Заключение
Зависимость высоты раковины моллюсков от их индивидуального возраста удовлетворительно описывается уравнениями криволинейной регрессии, а скорость роста – линиями регрессии. В Кандалакшском заливе в двух местах массового обитания исследуемого объекта (о. Ряшков и губа Порья) темпы роста моллюсков достоверно не различаются. Отмечено, что при пополнении выборки из Порьей губы значениями высот раковин особей в возрасте от 2 до 5 лет в этом месте обитания можно ожидать более ускоренный темп роста животных по сравнению с о. Ряшков. Кроме того, показано, что соленость воды – фактор, который влияет на распространение моллюсков в Кандалакшском заливе: при значении солености поверхностных вод ниже 12 ‰ Littorina littorea на литорали обнаружены не были.
Таким образом, изучение скорости роста Littorina littorea существенно дополняет сведения о биологии и экологии моллюска в конкретных условиях обитания.
Библиография
Алимов А. Ф., Макарова Г. Е., Максимович Н. В. Методы изучения двустворчатых моллюсков . Л.: Изд-во Зоологического ин-та АН СССР, 1990. 196 с.
Белое море. Биологические ресурсы и проблемы их рационального использования / Под. ред. О. А. Скарлато. Ч. 1. СПб.: Изд-во Зоологического ин-та РАН, 1995. 250 с.
Бергер В. Я. Дыхание некоторых литоральных моллюсков в процессе акклиматизации к изменениям солености воды // Экология. № 5. Свердловск: Наука, 1970. С. 68–72.
Бескупская Т. И. Питание некоторых массовых литоральных беспозвоночных Белого моря // Труды Кандалакшского заповедника. Труды биологической станции МГУ. 1963. Вып. 4. Т. 2. С. 135–169.
Бианки В. В., Бойко Н. С., Нинбург Е. А., Шкляревич Г. А. Питание обыкновенной гаги Белого моря // Экология и морфология гаг в СССР. М.: Наука, 1979. С. 126–170.
Брязгин В. Ф. Crustacea мелководной Порьей губы Кандалакшского залива Белого моря // Проблемы изучения, рационального использования и охраны ресурсов Белого моря: Материалы 9-й междунар. конф. 11–14 октября 2004 года. Петрозаводск, 2005. С. 67–71.
Голиков А. Н., Кусакин О. Г. Раковинные брюхоногие моллюски литорали морей СССР . Л.: Наука, 1978. 292 с.
Дорош Е. П. Запасы кормовых беспозвоночных для птиц и рыб на литорали островов Кандалакшского залива Белого моря // Труды Кандалакшского заповедника. 1963. Вып. 4. С. 54–67.
Зубов Н. Н. Океанологические таблицы . Л.: Гидрометеорологическое изд-во, 1957. 406 с.
Ивантер Э. В., Коросов А. В. Введение в количественную биологию: Учеб. пособие . Петрозаводск: Изд-во ПетрГУ, 2011. 302 с.
Краснов Ю. В., Шкляревич Г. А., Горяцев Ю. И. Характер и особенности питания обыкновенной гаги Somateria mollussima в Белом море // Доклады Академии наук. 2009. № 2. С. 282–285.
Коросов А. В., Горбач В. В. Компьютерная обработка биологических данных: Метод. пособие . Петрозаводск: Изд-во ПетрГУ, 2010. 84 с.
Мина М. В., Клевезаль Г. А. Рост животных: анализ на уровне организма . М.: Наука, 1976. 291 с.
Моллюски Белого моря / Под ред. О. А. Скарлато. Л.: Наука, 1987. 328 с.
Озернюк Н. Д., Зотин А. А. Сравнительный анализ роста мидий Mytillus edulis из разных районов Белого моря // Известия РАН. Сер. биологическая. 2006. № 2. С. 188–192.
Рубинчик М. А. К биологии размножения Littorina littorea L. Белого моря // Биология Белого моря. Т. 1. М.: Изд-во МГУ, 1962. С. 215–230.
Саранчова О. Л., Ушакова О. О., Беляева Д. В. Устойчивость личинок массовых видов беспозвоночных Белого моря к экстремальному изменению солености // Биология моря. СПб., 2006. Т. 32. № 6. С. 428–435.
Седловская С. М. Рост и продуктивность беспозвоночных животных: Метод. рекомендации . Витебск: УО «ВГУ им. П. М. Машерова», 2012. 48 с.
Федяков В. В. Закономерности распределения моллюсков Белого моря . Л., 1986. 126 с.
Шкляревич Г. А. Водоросли и беспозвоночные животные мелководий Порьей губы // Материалы по разнообразию биоты Кандалакшского заповедника – 1 / Под. ред. А. С. Корякина. Кандалакша, 1998. С. 12–78.
Шмелинг Г., Шилов Ю., Дракин А. Лоция Белого моря . URL: http://parusa.narod.ru/bib/books/ws_loc (дата обращения 21.12.2015).
Davies M. Role of mucus-trails and trail-following in the behavior and nutrition of the periwinkle Littorina littorea // Ecology Centre University of Sunderland. Marine Ecology Progress series of the United Kingdom, 1999. Vol. 179. P. 247–257.
Goodfriend G. A. Variation in land-snail form and size and causes: a rewiew // Systematic Zoology. 1986. Vol. 35. P. 204–223.
Kemp P., Bertness M. D. Snail shape and growth rates: evidence for plastic shell allometry in Littorina littorea // Proceeding of the National Academy of Sciences USA. 1984. Vol. 81. P. 811–813.
Yamada S. B., Mansour R. A. Growth inhibition of native Littorina saxatillis (Olivi) by introduced L.littorea // Exp. Marine Biology and Ecology. 1987. Vol. 105. P. 187–196.
Благодарности
Авторы выражают благодарность администрации Кандалакшского государственного природного заповедника за предоставление возможности осуществить данное исследование. Авторы благодарят А. В. Коросова за помощь в математической обработке данных и участие в обсуждении полученных результатов.

 © 2011 - 2026
© 2011 - 2026