



Введение
В настоящее время в экосистеме Баренцева моря важную роль играют два вида инвазивных крабов, имеющих промысловое значение: камчатский краб (Paralithodes camtschaticus) и обыкновенный краб-стригун или краб-стригун опилио (Chionoecetes opilio). Появление первого вида связано с целенаправленной интродукцией из районов нативного ареала, которая проходила во второй половине XX века. Многолетний мониторинг экспансии камчатского краба позволил весьма подробно исследовать и описать особенности его расселения и адаптации в баренцевоморском регионе (Камчатский краб…, 2003). На основе научных рекомендаций была организована успешная промысловая эксплуатация этого гидробионта на юго-востоке Баренцева моря. Результаты исследований показали, что в настоящее время в основных районах обитания камчатского краба процессы акклиматизации завершены или близки к своему завершению (Пинчуков, 2011; Pinchukov, Sundet, 2011).
Причины инвазии второго вида, краба-стригуна, до сих пор неясны и могут быть связаны как с непреднамеренным переносом личинок с балластными водами, так и с самостоятельной миграцией взрослых особей под влиянием естественных факторов среды (Кузьмин и др., 1998). Результаты десятилетнего мониторинга показывают, что этот краб образовал самовоспроизводящуюся популяцию в Баренцевом море, занимающую обширную акваторию. В то же время процессы акклиматизации этого вида пока не завершены. Краб активно осваивает новые районы, а также увеличивает свою численность на уже заселенных акваториях.
В данной работе впервые подробно анализируется динамика расселения краба-стригуна в Баренцевом море, а также рассматриваются факторы, влияющие на процессы акклиматизации. На основе моделирования пространственного распределения вида дается прогноз его дальнейшего расселения при различных температурных условиях.
Материалы
В работе использованы данные ПИНРО по уловам краба-стригуна в Баренцевом море, полученные в ходе:
1) наблюдений за промыслом донных видов рыб и научных съемок северной креветки в 1996-2003 гг.;
2) комплексной российско-норвежской экосистемной съемки в 2004-2014 гг.
Информация о приловах краба на донном промысле поступала с судов, на которых присутствовали научные группы или инспекторы рыбоохраны. В сообщениях указывались дата и координаты поимки, параметры орудия лова, количество пойманных крабов и их основные биологические характеристики (пол, ширина карапакса, состояние покровов). Общее количество пойманных крабов составило 164 экз. (табл. 1)
Таблица 1. Объем материала по крабу стригуну, полученного в ходе наблюдений за промыслом донных видов рыб и исследовательских съемок северной креветки в 1996-2003 гг.
| Год | Кол-во сообщений | Кол-во крабов, экз. |
| 1996 | 5 | 5 |
| 1997 | 1 | 1 |
| 1998 | 2 | 2 |
| 1999 | 7 | 8 |
| 2000 | 12 | 8 |
| 2001 | 9 | 5 |
| 2002 | 21 | 28 |
| 2003 | 79 | 107 |
Исследования расселения краба-стригуна на регулярной основе стали возможны с началом проведения (2004 г.) российско-норвежских комплексных экосистемных съемок. Ежегодные съемки проводились по стандартной методике в летне-осенний период и охватывали большую часть акватории Баренцева моря, площадь которой в среднем составляет порядка 1500 тыс. км2 (Eriksen, 2012). Каждая съемка проводилась одновременно на 4-5 судах. Сбор материала осуществлялся донным тралом Campelen с горизонтальным раскрытием 25 м, вертикальным – 5 м и вставкой в кутовой части из дели с ячеей 22 мм. Количество донных тралений, выполненных за съемку, варьировало от 650 до 1123. Общее количество пойманных крабов составило 77854 экз. (табл. 2).
Таблица 2. Объем материала по крабу стригуну, полученного в ходе экосистемных съемок Баренцева моря 2004-2014 гг.
| Год | Кол-во тралений | Кол-во тралений с крабом | Кол-во крабов, экз. |
| 2004 | 1123 | 5 | 6 |
| 2005 | 1008 | 10 | 14 |
| 2006 | 999 | 29 | 61 |
| 2007 | 1007 | 56 | 134 |
| 2008 | 776 | 77 | 670 |
| 2009 | 754 | 66 | 284 |
| 2010 | 710 | 58 | 400 |
| 2011 | 775 | 84 | 6657 |
| 2012 | 835 | 121 | 37737 |
| 2013 | 859 | 132 | 19020 |
| 2014 | 650 | 87 | 12871 |
Методы
Для расчетов вероятности встречаемости краба-стригуна акватория Баренцева моря была разбита на полигоны регулярной сеткой с шагом 0,25° северной широты и 1° восточной долготы. Атрибуты полигона включали в себя следующие показатели: номер, координаты, глубина (м), придонная температура (°С), расстояние (км) от геометрического центра акватории распределения краба, поимка краба (1 – краб есть; 0 – краба нет; NA – наблюдение отсутствует). Расстояние от геометрического центра акватории распределения краба рассчитывалось ежегодно для каждой съемки и служило фактором удаленности от ядра начального расселения краба. Для построения распределений вероятности встречаемости краба по годам и прогноза его дальнейшего распространения была выбрана обобщенная линейная модель (GLM), реализованная в пакете biomod2 статистической платформы R. Выбор модели, ее диагностика, а также оценка влияния переменных на результаты моделирования осуществлены с помощью встроенных функций пакета biomod2 3.1 (https://cran.r-project.org/web/packages/biomod2/). В качестве факторов, определяющих вероятность встречаемости краба в каждом полигоне в период проведения съемок, использовались: глубина, температура и расстояние от центра расселения. При прогнозе потенциального ареала краба в Баренцевом море учитывались только глубина и температура, а расстояние от центра расселения исключалось. При этом было выполнено три варианта расчетов при разной температуре: среднемноголетней за 2010-2014 гг., ниже среднемноголетней на 1 °C и выше среднемноголетней на 1 °C. Площадь распространения краба рассчитывалась на акватории с вероятностью встречаемости краба более 50%, т.е. в тех случаях, когда вид можно считать константным (Иоганзен, Файзова, 1978). Построение картограмм результатов моделирования также осуществлялось в R с помощью пакета PBSmapping (https://cran.r-project.org/web/packages/PBSmapping/).
Для расчетов плотности распределения и индексов численности краба по годам была выделена акватория с высокой встречаемостью животных, которая была разбита на районы и подрайоны (страты с диапазоном глубин 0-100, 100-200, 200-300 и более 300 м). Индекс численности в страте (N) рассчитывался площадным методом: N=C*(S/T), где С – средний улов краба на траление, на акватории подрайона с площадью S, а площадь траления (Т) принималась равной 0,022224 км2. Протраленная площадь рассчитана с учетом стандартного 15-минутного траления с горизонтальным раскрытием трала 15 м и скоростью 3,2 узла.
Результаты
Краб-стригун Chionoecetes opilio (Fabricius, 1788) (Brachyura, Majidae) является обычным и массовым представителем донных сообществ шельфа и материкового склона северо-западной Атлантики и северной Пацифики. (Кобякова, 1958; Слизкин, 1982; Галкин, 1985; Davidson et al.,1985). В 1996 г. краб впервые был отмечен в Баренцевом море на северном склоне Гусиной банки в ходе исследовательской съемки запасов северной креветки (Кузьмин и др., 1998). С этого времени наблюдается постепенное увеличение ежегодного количества сообщений о приловах краба на склонах Гусиной и Демидовской банках (Kuzmin, 2000; Павлов, 2006) (рис. 1).
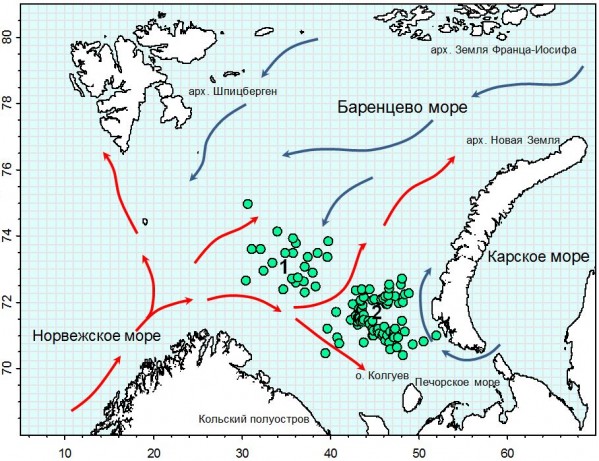
Рис.1. Места поимок краба-стригуна в 1996-2003 гг. в Баренцевом море (1 – Демидовская банка, 2 – Гусиная банка) и основные направления теплых (красные стрелки) и холодных (синие стрелки) течений
Figure 1. Places of catches of snow crab opilio in the Barents Sea (1 – Demidovskaya Bank, 2 – Gusinaya bank) in 1996-2003 and the main directions of warm (red arrows) and cold (blue arrows) currents.
За период исследований с 1996 по 2003 гг. краб-стригун в Баренцевом море отмечен на глубинах 100-330 м в основном на илистых и песчано-илистых грунтах, реже на илисто-глинистых и песчано-глинистых. Температура воды в пределах его обитания в придонном слое варьировала от -1,5 до +4,3 °С, соленость - от 34,5 до 35,1 ‰. В диапазоне температур от -1,1 до +1,3 °С уловы крабов достигали максимальных значений.
С 2002 г. количество сообщений о приловах краба возросло (см. табл. 1). Животные отмечались не только в районах их первой регистрации, но и далеко за пределами склонов Гусиной банки. Наличие функционально половозрелых особей вида, а также нахождение самок с икрой служило подтверждением успешного размножения крабов-стригунов в новом районе обитания. В соответствии с основными направлениями придонных течений был определен наиболее вероятный северо-восточный вектор экспансии краба на личиночной планктонной стадии. Однако отсутствие систематических исследований и промысла не позволяло достоверно оценить наличие крабов в северо-восточных районах Баренцева моря до 2004 г.
Начиная с 2004 г., в ходе экосистемных съемок, исследования показали увеличение ареала, плотности скоплений и численности краба-стригуна в новом регионе (табл. 3). Акватория встречаемости краба в 2005 г. увеличилась в 3 раза, а в 2006 г. еще вдвое по сравнению с предыдущими наблюдениями. За первые 4 года исследований площадь распространения увеличилась в 10 раз. С 2007 г. эта площадь в среднем ежегодно увеличивалась на 10 %. Сокращение площади распространения в 2014 г. скорее всего свидетельствует не о реальных процессах в динамике популяции, вызванных естественными причинами смертности и пополнения, а о технических особенностях съемки, повлекших за собой изменения в уловистости трала для бентосных организмов (устные сообщения Прозоркевича Д. В. и Любина П. А.).
Таблица 3. Площадь распространения, средняя плотность распределения и индекс численности краба-стригуна в Баренцевом море в 2004-2014 гг.
| Год | Площадь распространения, тыс. км | Средняя плотность распределения, экз./км2 | Индекс численности, млн. экз. |
| 2004 | 44 | 1 | 0,63 |
| 2005 | 130 | 3 | 2,21 |
| 2006 | 262 | 7 | 5,24 |
| 2007 | 336 | 19 | 14,31 |
| 2008 | 447 | 116 | 86,38 |
| 2009 | 514 | 36 | 26,91 |
| 2010 | 557 | 76 | 56,25 |
| 2011 | 605 | 1144 | 849,63 |
| 2012 | 569 | 5854 | 4346,32 |
| 2013 | 662 | 3071 | 2280,25 |
| 2014 | 452 | 2234 | 1658,43 |
Наряду с увеличением площади распространения краба, увеличивалась относительная плотность его распределения (табл. 3). До 2007 г. крабы в уловах встречались единично. Средние уловы краба на траление существенно возросли с 2011 г. С этого времени в размерном составе уловов стала преобладать неполовозрелая молодь, которая в настоящее время составляет основу численности популяции. За весь период исследований индекс численности возрос на три порядка; такой взрывной характер численности часто наблюдается у акклиматизированных животных в начале периода акклиматизации. Однако в последние два года отмечается снижение средних уловов краба в съемках. Природа этого снижения пока точно не установлена и требует дальнейшего исследования.
Картирование результатов моделирования распространения краба-стригуна по результатам экосистемных съемок 2004-2014 гг. показало, что расширение ареала проходило в северном, южном и западном направлениях (рис. 2). В 2005 г. восточная граница встречаемости вплотную приблизилась к берегам арх. Новая Земля. В 2008 г. южные и юго-восточные границы распределения также приблизились к своему географическому пределу – прибрежным районам Печорского моря (о-ва Колгуев и Вайгач, п-ов Канин, пролив Карские Ворота). В этот период происходило формирование современных юго-западных границ распространения краба. Лимитирующим фактором дальнейшего распространения при этом стала высокая придонная температура (5-7°С) прибрежных районов Кольского полуострова, при которой вероятность встречаемости краба составила менее 20 %.
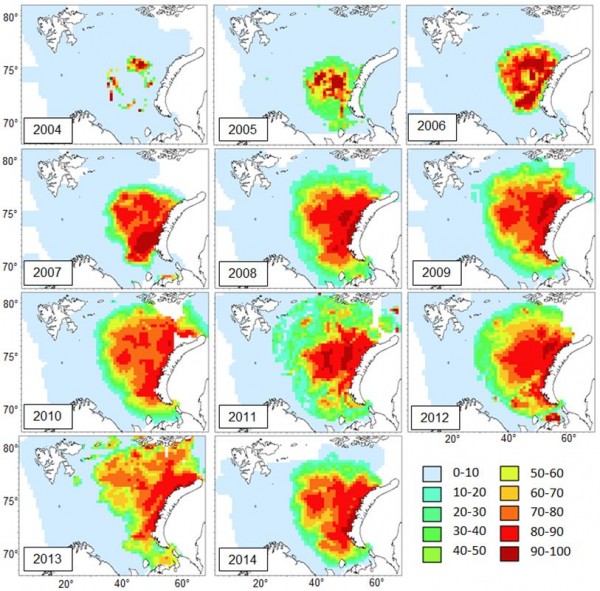
Рис. 2 Вероятность встречаемости (%) краба-стригуна в Баренцевом море в 2004-2014 гг.
Figure 2. Occurrence probability (%) of snow crab opilio in the Barents Sea in 2004-2014.
Расселение краба-стригуна в северном, северо-западном и северо-восточном направлении отмечалось в течение всего периода наблюдений. В 2009 г. краб был отмечен в прибрежных районах арх. Земля Франца-Иосифа (ЗФИ). С 2010 г. краб стал регулярно встречаться не только в районах ЗФИ, но и других районах северо-востока Баренцева моря, достигнув границ проведения экосистемной съемки. В связи с этим мониторинг расселения краба в северном направлении в настоящее время затруднен из-за отсутствия наблюдений. В 2010-2011 гг. замедлилась экспансия краба в северо-западном направлении, не достигнув при этом восточных берегов арх. Шпицберген. Единичные поимки крабов западней 34-35° в. д. в период исследований носили случайный характер и не могут достоверно свидетельствовать о массовом расселении краба в западных районах Баренцева моря.
Анализ фактора удаленности от центра расселения показал, что наиболее активная экспансия краба была отмечена в северо-восточном направлении. Скорость расселения в этом направлении составила в среднем 105 км/год. Расселение краба как в южном, так и северном направлениях проходило со средней скоростью 69 км/год. Продвижение на запад происходило не так активно и в среднем составило 39 км/год.
Анализ распределения уловов краба по глубинам за весь период наблюдений показал, что наиболее часто краб отмечался на глубинах 150-250 м. В то же время краб периодически встречался на мелководьях (до 100 м) и глубоководных впадинах (более 250 м). С расселением краба в восточном и юго-восточном направлении его уловы увеличились на мелководных участках (до 50 м), прилегающих к арх. Новая Земля. В 2012-2013 гг. краб начал встречаться на мелководьях Печорского моря с глубинами 17-20 м. В настоящее время это наименьшая глубина, на которой регистрировались уловы краба в Баренцевом море. Максимальная глубина обнаружения краба была отмечена в 2011 г. и составила 551 м. В этот период расселение краба в северо-восточном направлении достигло границ глубоководного желоба Св. Анны на границе Баренцева и Карского морей.
Результаты экосистемных съемок показали, что границы распространения краба-стригуна варьируют из года в год. Однако с 2010 г. площадь акватории распределения краба значительно не меняется, а границы высоких плотностей остаются практически неизменными. С учетом этого ниже приводятся особенности современного состояния популяции краба-стригуна в Баренцевом море, оцененные на основе объединенных данных исследовательских съемок 2010-2014 гг.
В настоящее время ареал краба-стригуна охватывает 34% площади Баренцева моря. Площадь распространения краба составляет 618 тыс. км2. Краб стал обычным представителем бентосной фауны в восточной части Баренцева моря от о. Колгуев на юге до арх. Франца-Иосифа на севере. Наиболее плотные скопления молоди встречаются в юго-восточном районе (Печорское море) и северо-восточных акваториях у побережья арх. Новая Земля. Скопления половозрелых особей отмечаются в центральных (Центральная возвышеность) и центрально-восточных (Новоземельская банка) районах. Ареал краба достиг своих географических границ на востоке, приблизившись к береговой линии арх. Новая Земля. На юго-востоке также завершилось активное расселение краба; границы его ареала совпадают с южной границей Печорского моря (рис. 3А).
Прогноз дальнейшего расселения можно продемонстрировать с помощью моделирования распределения без учета факторов времени и дистанции от геометрического центра акватории распределения краба, используя только фактор температуры и глубины. При этом долговременное изменение температуры может существенно влиять на распределение краба (рис. 3). При температуре, которая наблюдалась в 2010-2014 гг., акватория встречаемости краба может увеличиться в 2 раза, при этом площадь распространения краба с вероятностью встречаемости более 50% увеличится до 1237 тыс. км2. Расширение ареала произойдет за счет северных и северо-западных районов Баренцева моря (рис. 3Б). При похолодании придонных слоев на 1°C акватория увеличится в 3 раза по сравнению с современным распределением (рис. 3В). Площадь распространения вида может составить 1421 тыс. км2. Похолодание будет способствовать дальнейшей экспансии краба в западном направлении. При увеличении средней придонной температуры на 1 °C экспансия замедлится, а площадь распространения краба при его полной акклиматизации в Баренцевом море может составить 1019 тыс. км2, т.е. на 20% больше площади современного распределения.
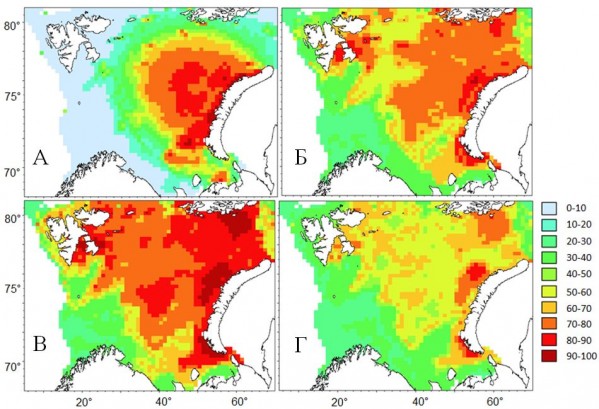
Рис. 3 Вероятность встречаемости (%) краба-стригуна в Баренцевом море в 2010-2014 гг. (А), а также прогноз распределения при среднемноголетней температуре (Б), ниже среднемноголетней на 1 °C (В), выше среднемноголетней на 1 °C (Г).
Figure 3. Occurrence probability (%) of snow crab opilio in the Barents Sea in 2010-2014 (А), and the forecast of its distribution at the temperature equal to the long-term average annual temperature (Б), 1 °C lower than the long-term average annual temperature (В), 1 °C higher than the long-term average annual temperature (Г).
Как указывалось выше, данный прогноз расселения основан на влиянии двух факторов среды: глубины и придонной температуры. Анализ распределения глубин в Баренцевом море и распределения краба по глубинам показал, что фактор глубины не является в значительной степени лимитирующим для дальнейшего расселения краба (рис. 4А). Краб в настоящее время встречается практически во всем диапазоне глубин от 20 до 550 м. Данный диапазон охватывает 98 % площади Баренцева моря. Данные моделирования показывают, что вклад фактора глубины в модель составляет 28 %. Анализ вероятности присутствия краба по глубинам показал, что оптимальный диапазон глубин для краба стригуна находится в пределах от 0 до 300 м (рис. 4Б). Данный диапазон глубин охватывает 83 % площади Баренцева моря.
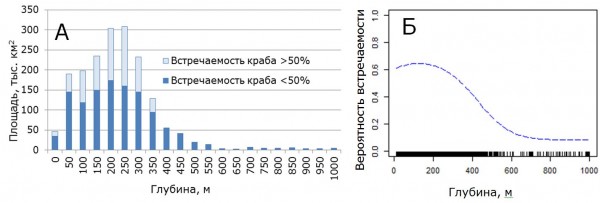
Рис. 4. Площадное распределение глубин Баренцева моря с учетом встречаемости краба-стригуна в 2010-2014 гг. (А) и кривая влияния глубин на вероятность его встречаемости (Б).
Figure 4. Areal distribution of Barents Sea depths taking into account the occurrence of snow crab opilio in 2010-2014 (А) and the curve of the depth influence on the occurrence probability (Б).
Анализ распределения придонной температуры в Баренцевом море и распределения краба показал, что фактор температуры является в значительной степени лимитирующим для дальнейшего расселения краба в районы с повышенным теплосодержанием вод (рис. 5А). Краб в настоящее время встречается в диапазоне придонных температур от -1,5°C до 6,5 °C. Однако наибольшая встречаемость отмечена при придонных температурах от -1,5°C до 3,0 °C. Этот диапазон охватывает 82% учетной площади в Баренцевом море. Данные моделирования показывают, что вклад фактора температуры в модель составляет 72 %. Анализ вероятности присутствия краба показал, что наиболее оптимальный диапазон температур для краба стригуна находится в пределах -1,5°C до 2,0 °C. Данный диапазон глубин охватывает 70 % площади Баренцева моря.
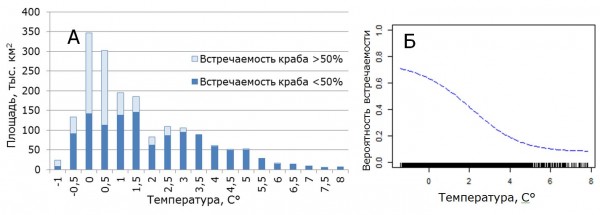
Рис. 5. Площадное распределение придонной температуры Баренцева моря с учетом встречаемости краба-стригуна в 2010-2014 гг. (А) и кривая влияния температуры на вероятность его встречаемости (Б).
Figure 5. Areal distribution of the Barents Sea bottom temperature taking into account the occurrence of snow crab opilio in 2010-2014 (А) and the curve of the temperature influence on the occurrence probability (Б).
Обсуждение
В литературе отмечается, что основными факторами, влияющими на распределение и формирование промысловых скоплений краба, являются температура, глубина и тип грунта (Слизкин, 1982; Михайлов и др., 2003). Предпочитаемый диапазон глубин и температур краба-стригуна в Баренцевом море весьма сходен с условиями обитания популяций краба в традиционных районах его обитания от Северо-Восточной Атлантики до Японского моря. Наиболее холодноводные и глубоководные скопления этого вида, имеющие промысловое значение, были отмечены у берегов Гренландии. В этом районе краб часто живет в области отрицательных температур и глубин более 300-400 м. Максимальная глубина, на которой были обнаружены крабы – 1400 м. Основные глубины обитания промысловых крабов 150-250 м характерны для популяций, обитающих в водах американского побережья Берингова моря, канадской зоны и дальневосточного региона России. Промысловые скопления на глубинах менее 100 м краб образует у восточного побережья Канады. Верхний предел температур для краба ограничивается 9-10 °С. Однако с увеличением среднегодовой придонной температуры до 6 °С плотность поселений краба заметно снижается, а промысловые скопления практически не встречаются. Южная граница обитания краба в Тихом океане проходит у берегов Японии в префектуре Киото. Поверхностные воды в этом районе теплые, поэтому краб обитает в основном на глубинах более 200 м с придонными температурами не выше 5 °С.
Таким образом, учитывая тепловой режим и батиметрическое распределение краба в нативном ареале и Баренцевом море, лимитирующим фактором дальнейшего расселения краба может стать высокая придонная температура. Такие придонные температуры (>6°С) характерны для юго-западной части Баренцева моря.
При оценке распределения и дальнейшего расселения необходимо учитывать направление и скорость течений, тип грунта, соленость и наличие кормового бентоса. В нативном ареале краб предпочитает илистые и песчано-илистые грунты. Однако молодь, которая в большей степени встречается на мелководье, чаще отмечается на песчаных и илисто-гравийных грунтах (Иванов, Соколов, 1997, Слизкин, 1982; Dufour, 1988). Большая часть дна Баренцева моря покрыта песчаным илом. Прибрежные районы, склоны и возвышенности банок покрыты илистым песком. В некоторых районах имеется песок. Юго-восточную часть моря занимают илистые отложения (Добровольский, Залогин, 1982). Таким образом, тип грунта оптимален для жизнедеятельности и не будет препятствовать дальнейшему расселению краба в Баренцевом море.
Одним из основных абиотических факторов, способствующих широкому расселению краба в Дальневосточном регионе, указывается направление течений, переносящих личинок (Слизкин, 1982). В Баренцевом море существует сложная система поверхностных и глубинных течений, самым общим свойством которых является движение вод против часовой стрелки (Новицкий, 1961). Это свойство благоприятно сказалось на успешной акклиматизации вида, направив основной вектор расселения в северо-восточном направлении в район с наиболее оптимальным температурным режимом (менее 3°С) для развития молоди. Дальнейшему расселению краба в северо-западном направлении будут способствовать холодные течения из Арктического бассейна, направленные к югу от Земли Франца-Иосифа и вдоль восточного берега Шпицбергена.
Одним из возможных факторов, влияющих на распространение краба в прибрежных районах, может стать соленость придонных вод, которые часто подвержены значительному опреснению (Anger, 2003). В то же время известно, что краб достаточно часто образует плотные скопления при пониженной солености вплоть до 32 ‰ (Слизкин, 1982). Учитывая, что 2/3 акватории Баренцева моря находятся под влиянием атлантических вод и даже на поверхности моря соленость воды превышает 34‰, а на остальной акватории моря соленость колеблется в пределах 32-34 ‰ (Добровольский, Залогин, 1982), фактор солености не будет играть значимую роль в дальнейшем расселении краба.
Таким образом, основные абиотические факторы среды в Баренцевом море, влияющие на распределение краба-стригуна, не препятствуют акклиматизации вида, колонизации акваторий и формированию скоплений в северном и северо-западном районах Баренцева моря. Многие авторы в числе факторов, влияющих на распределение краба-стригуна, справедливо указывают следующие биотические особенности: наличие кормового бентоса и хищников (Кобякова, 1958; Слизкин, 1982; Галкин, 1985). Бесспорно, что для формирования плотных скоплений краба, на основе которых можно организовать успешный промысел, необходимы существенные пищевые ресурсы. Весьма важно при этом отсутствие каннибализма и массовых хищников, которые могли бы подорвать численность популяции на ранних стадиях развития. Однако большинство авторов отмечают важность наличия кормовой базы в контексте формирования промысловых скоплений. К сожалению, недостаток данных о распределении кормового бентоса в северо-западных районах Баренцева моря не позволяет количественно доказать эту гипотезу. Однако наличие пищевых ресурсов для краба-стригуна можно подтвердить косвенно через присутствие видов, имеющих схожие пищевые предпочтения, в районах, перспективных для расселения краба-стригуна.
Анализ питания краба-стригуна в Баренцевом море показал, что наиболее близкими пищевыми конкурентами для него являются два вида краба-аборигена Hyas araneus и Lithodes maja, которые весьма часто отмечаются в северо-западных районах моря (Павлов, 2007). Кроме того, постоянное присутствие в этих районах рыб-бентофагов также свидетельствует о наличии удовлетворительной кормовой базы в этих районах для дальнейшего расселения краба-стригуна. В то же время оценка достаточности этих пищевых ресурсов для формирования промысловых скоплений требует дальнейших исследований.
Заключение
Динамика расселения краба-стригуна в Баренцевом море с 2004 г. соответствует процессу акклиматизации и формирования новой популяции. За период исследований площадь распространения этого вида увеличилась в 10 раз, а численность возросла на три порядка.
Основным фактором, лимитирующим дальнейшее распространение краба, может стать придонная температура на юго-западе моря. Однако потенциал дальнейшего расселения краба в Баренцевом море остается весьма высоким. Прогностические оценки показывают, что его ареал может увеличиться в два раза за счет северо-западных районов Баренцева моря и прибрежных акваторий арх. Шпицберген.
Похолодание будет способствовать дальнейшей экспансии краба в западном направлении и увеличению его численности в традиционных районах обитания. При потеплении придонных слоев экспансия замедлится.
Библиография
Добровольский А.Д., Залогин Б.С. Моря СССР. М.: Изд-во МГУ, 1982. 192 с.
Галкин Ю.И. К вопросу об увеличении промысловой продуктивности Белого и Баренцева морей путем акклиматизации // Экологические исследования перспективных объектов матикультуры в Белом море. Л.: Изд-во Зоол. Ин-т АН СССР, 1985. C. 122-133.
Иванов Б.Г., Соколов В.И. Краб-стригун Chionoecetes opilio (Crustacea Decapoda Brachyura Majidae) в Охотском и Беринговом морях // Arthropoda Selecta. 1997. Т. 6, вып. 3-4. С. 63-86.
Иоганзен Б.Г., Файзова Л.В. Об определении показателей встречаемости, обилия, биомассы и их соотношения у некоторых гидробионтов // Тр. ВГБО. 1978. T. 22. С. 215-225.
Камчатский краб в Баренцевом море. Изд. 2–е перераб. и доп. Мурманск: Изд-во ПИНРО, 2003. 383 с.
Кобякова З.И. Десятиногие раки (Decapoda) района южных Курильских островов // Исследования Дальневосточных морей. М.-Л. 1958. Т. 5, С. 220-248.
Кузьмин С.А., Ахтарин С.М., Менис Д.Т. Первые нахождения краба-стригуна Chionoecetes opilio (Decapoda, Majidae) в Баренцевом море // Зоол. журн. 1998. Т. 77. № 4. С. 489-491.
Пинчуков М.А. Особенности расселения и динамика запаса камчатского краба в Баренцевом море // Ж. Рыбное хозяйство, юбилейный спецвыпуск, 2011. С. 65-67.
Михайлов В.И., Бандурин К.В., Горничных А.В., Карасев А.Н. Промысловые беспозвоночные шельфа и материкового склона северной части Охотского моря //Магадан: Изд-во МагаданНИРО. 2003. 281 c.
Новицкий В.П. Постоянные течения северной части Баренцева моря // Труды ГОИН. 1961. Вып. 64. С.3-32.
Павлов В.А. Новые данные о крабе-стригуне Chionoecetes opilio (Fabricius, 1788) Баренцева моря // Тез. докл. VII Всерос. конф. по промысловым беспозвоночным (памяти Б.Г. Иванова), Мурманск, 9-13 октября 2006 г. М.: Изд-во ВНИРО. 2006. С. 109-111
Павлов В.А. Питание краба-стригуна опилио Chionoecetes opilio (Fabricius, 1788) в Баренцевом море // Морские промысловые беспозвоночные и водоросли: биология и промысел. К 70-летию со дня рождения Бориса Георгиевича Иванова. Труды ВНИРО. 2007. Т. 147. М. Изд-во ВНИРО. С. 99-107.
Слизкин А.Г. Распределение крабов-стригунов рода Chionoecetes и условия их обитания в северной части Тихого океана // Изв. ТИНРО. 1982. Т. 106. С. 26-33.
Anger, K. Salinity as a key parameter in the larval biology of decapod crustaceans. Invertebr. Reprod. Dev. Vol. 43. 2003. P. 29–45.
Davidson, K. Morphological, Electrophoretic, and Fecundity Characteristics of Atlantic Snow Crab, Chionoecetes opilio, and Implications for Fisheries Management / K. Davidson, J.C. Roff, R.W. Elner // Canadian Journal of Fisheries and Aquatic Sciences. 1985. Vol. 42, No.3. – P. 474-482.
Dufour, R. 1988. Overview of the distribution and movement of snow crab (Chionoecetes opilio) in Atlantic Canada. In: Proceedings of the International Workshop on Snow Crab Biology, December 8–10, 1987, Montreal, Quebec. G. S. Jamieson, and W. D. McKone (eds.) Can. MS. Rep. Fish. Aquat. Sci., 2005. P. 75–82
Eriksen, E. (ed.) 2012. Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea, August – September 2012. Joint IMR/PINRO report series, 2012-2. ISSN 1502‐8828, 108 pp.
Kuzmin S.A. Distribution of snow crab Chionaecetes opilio (Fabricius) in the Barents Sea // ICES CM 2000/U:21. 2000. –7 p.
Pinchukov M.A. and Sundet J.H. Red king crab. In book: The Barents Sea. Ecosystem, Resources, Management. Tapir academic press. Half a century of Russian – Norwegian cooperation. Edited by Tore Jakobsen and Vladimir K. Ozhigin. 2011. P. 160-167.

 © 2011 - 2026
© 2011 - 2026