



Введение
Вопросам роста сеголеток лягушек, начальные стадии развития которых проходят в нестабильных условиях временных водоемов, посвящены подробные исследования в природе и эксперименты. Целью было выяснить, как зависят их размеры от вариабельности размера икринок, от особенностей роста и развития личинок, от исходных размеров лягушат при появлении на суше и специфики их роста в первые недели жизни.
Показано, что удельная скорость роста может зависеть или не зависеть от размеров прошедших метаморфоз особей, что большая изменчивость этой величины характерна как для мелких, так и для более крупных особей. Изменчивость размеров осенью может зависеть или не зависеть от времени выхода сеголеток и т. д. (Смирина, 1980; Ищенко, Леденцов, 1985; Ляпков, 1989 и др.).
Авторы подчеркивают, что во всех проявлениях изменчивости размеров на всех стадиях жизненного цикла бесхвостых земноводных существенную роль должны играть абиотические факторы, однако исследования подобного рода, основанные на анализе многолетних рядов данных, нам неизвестны.
В изученных условиях у сеголеток лягушек есть обычно менее двух месяцев для того, чтобы достичь необходимых размеров и сформировать достаточные запасы энергии для успешной продолжительной зимовки (Кутенков, 2009). И сеголеткам популяций севера Европы приходится осуществлять эти процессы в условиях изменчивого и неустойчивого климата.
Цель нашего исследования, основанного на результатах многолетних наблюдений, состояла в выяснении роли погодных условий в период нагула сеголеток на формирование их размерной вариабельности перед зимовкой и количественная оценка влияния этих факторов.
Материалы
Работа проведена в заповеднике «Кивач» (южная Карелия, Кондопожский р-н), где с начала 1980-х гг. вели наблюдения за популяцией R. temporaria (Кутенков, 1998). Занимаемая ею территория площадью около 60 га включает удобный зимовочный водоем (неглубокая порожистая река), хвойно-лиственный лес и лугá, используемые как нагульные местообитания, а также около 60 нерестилищ лягушек (средние координаты 62° 16′ 35″ с. ш., 33° 59′ в. д.). В центре участка расположена метеостанция заповедника, непрерывные наблюдения на которой проводятся с 1969 г. (Скороходова, 2008).
Завершившие метаморфоз лягушата широко и достаточно быстро расселяются по территории, концентрируясь в открытых стациях и по опушкам, где и осуществляют нагул. Их рост фактически прекращается в первую декаду сентября из-за снижения температуры воздуха, и до ухода на зимовку у них остаются 1.5–2 недели. В годы наблюдений в период с 28 августа по 8 сентября делали однократные обходы луговых участков, собирали сеголеток, с помощью штангенциркуля измеряли у них длину тела (от конца морды до клоаки, точность до 0.1 мм) и выпускали обратно. Промеры производили в 1981 г. и, с отдельными пропусками, в 1988–2013 гг., всего 22 года наблюдений. В разные годы в выборках оказывались по 8–80 экз., в среднем 34 экз. Пропуски, а также малые выборки обычно были вызваны очень низкой численностью животных новой генерации в отдельные годы. Всего за время работ измерили 741 экз. сеголеток.
Погодные характеристики с мая по сентябрь получены из данных метеостанции заповедника. Ежегодно фиксировали даты начала икрометания, вылупления головастиков и начала выхода сеголеток в репродуктивных водоемах в пределах территории контрольной популяции R. temporaria.
Методы
Основные этапы репродуктивного цикла популяции травяной лягушки наступали в районе исследований в следующие сроки (приведены средние многолетние даты за последние 33 года): начало икрометания — 30 апреля, вылупление головастиков — 11 мая, появление первых сеголеток (начало нагула) — 6 июля, окончанием нагула считали 1 сентября: среднесуточная температура воздуха первой декады сентября составляет для Кивача всего 10.7 °С (Скороходова, 2008).
Для анализа были избраны следующие переменные. Среднемесячная температура воздуха: трех летних месяцев (Т1), июля (Т2), первого месяца нагула сеголеток (средняя для 30 дней от даты появления сеголеток в данном году) (Т3), июля и августа (Т4). Сумма осадков, мм: в июле (Ос1), августе (Ос2) июле и августе (Ос3); количество суток с дождями, ≥1 мм: в июле (Ос4), августе (Ос5), июле и августе (Ос6). В качестве переменных длительности той или иной стадии развития и роста выбрали следующие сроки: дней от выклева головастиков до появления сеголеток (Д1), дней нагула в июле и августе (Д2), дней нагула до 1 сентября (т. е. общая продолжительность нагула в данный сезон) (Д3).
Зависимой переменной служили средние размеры сеголеток в осенних выборках (Lсгл, мм).
Для оценки зависимости многолетнего варьирования размеров тела сеголеток перед зимовкой использовали метод множественного регрессионного анализа с пошаговым отбором независимых переменных (подробности см. Kutenkov, Mosiyash, 2000; Кутенков, 2009).
Статистическую обработку осуществляли в пакетах Statistica 6.0, MS Exel и StatGraphics for Windows.
Результаты
Вариация индивидуальных размеров тела сеголеток в конце нагула в суммарной выборке за все годы составляла от 13 мм до 36 мм. Вариация средних размеров в выборках разных лет (Lсгл) была от 16.6 до 27.2 мм, т. е. различалась более чем в полтора раза; средняя многолетняя величина составила 21.8 мм. Частотное распределение длины тела в суммарной выборке (рис. 1) таково, что размеры двух третей особей укладываются в диапазон 19–25 мм.
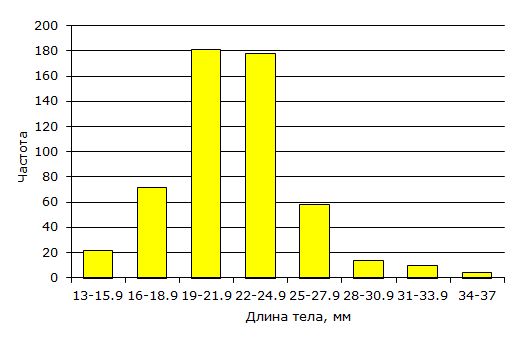
Рис. 1. Распределение сеголеток травяной лягушки по длине тела после окончания периода нагула (n = 540)
Fig. 1. Size (body length) distribution of youngs of the year of сommon frog at the end of foraging period (n = 540)
Вариации показателей температуры, количества и длительности осадков и продолжительности отдельных периодов жизни R. temporaria, использованных в расчетах, приведены в таблице. Ее анализ позволяет заметить следующее. Флуктуации погодных условий и сроков в годы наших наблюдений несущественно отличаются от таковых, рассчитанных для последней трети столетия, разве что показатели температуры воздуха и сумм осадков (средние и минимальные) оказались в нашем случае несколько выше. Это позволяет считать использованную «выборку» лет репрезентативной.
Значения летних температур влияют на конечные размеры сеголеток сильнее, чем любые другие действующие факторы. Обилие и продолжительность осадков в июле (Ос1, Ос4) негативно влияют на итоговые размеры сеголеток, тогда как их суммарные характеристики за период нагула (Ос3, Ос6) не играют никакой роли.
Таблица. Варьирование погодных характеристик и продолжительности периодов роста и развития личинок и сеголеток Rana temporaria в годы наблюдений и за последние 34 года. Сила влияния (ранговая корреляция Спирмена) на варьирование средних значений длины тела сеголеток
| Вариация | Сила | ||||||
| Показатель* | в годы наблюдений (n = 22) | в 1980–2013 гг. (n = 34) | влияния, n = 22 | ||||
| сред. | max | min | сред. | max | min | rs (p) | |
| Т1 | 15.0 | 16.8 | 13.1 | 14.9 | 16.8 | 12.8 | 0.84 (0.000) |
| Т2 | 16.5 | 21.3 | 13.1 | 16.3 | 21.3 | 13.1 | 0.55 (0.009) |
| Т3 | 17.2 | 21.9 | 14.1 | 16.7 | 21.9 | 12.8 | 0.64 (0.001) |
| Т4 | 15.7 | 18.7 | 14.1 | 15.4 | 18.7 | 12.8 | 0.68 (0.0005) |
| О1 | 81.8 | 132.3 | 32.5 | 76.3 | 132.3 | 23.4 | -0.58 (0.004) |
| Ос2 | 93.5 | 241.9 | 27.2 | 87.8 | 241.9 | 19.8 | 0.13 (н.д.) |
| Ос3 | 175.4 | 324.1 | 82.0 | 165.4 | 324.1 | 50.4 | 0.08 (н.д.) |
| Ос4 | 11 | 18 | 5 | 10 | 18 | 5 | -0.25 (0.25) |
| Ос5 | 11 | 19 | 4 | 11 | 19 | 4 | 0.07 (н.д.) |
| Ос6 | 21 | 29 | 11 | 21 | 29 | 11 | 0.1 (н.д.) |
| Д1 | 59 | 74 | 44 | 58 | 74 | 43 | -0.53 (0.001) |
| Д2 | 21 | 31 | 5 | 22 | 31 | 5 | 0.48 (0.02) |
| Д3 | 52 | 67 | 36 | 54 | 72 | 36 | 0.50 (0,02) |
Примечание. * — определение показателей приведено в разделе «Методы».
Процедура пошагового отбора независимых переменных, значения которых приведены в таблице, выделила те из них, влияние которых на конечные размеры сеголеток (Lсгл) было наибольшим. Уравнение множественной регрессии имеет вид:
Lсгл = 11.71 + 1.16 · T1 – 0,02 · Ос1 – 0.1 · Д1
Зависимость достоверна при уровне значимости р = 0.0005. Коэффициент детерминации этой регрессии R2 = 0.613 (множественная корреляция r = 0.78). Иными словами, вариация конечных размеров сеголеток на 61.3 % определяется выделенными переменными. При этом на среднемесячную температуру июня-августа приходится 32.1 % суммарной доли влияния, на долю суммы осадков июля 9.6 % и на продолжительность личиночного развития — 19.6 %. Графическое сравнение наблюдаемых и рассчитанных значений размеров сеголеток дано на рис. 2.
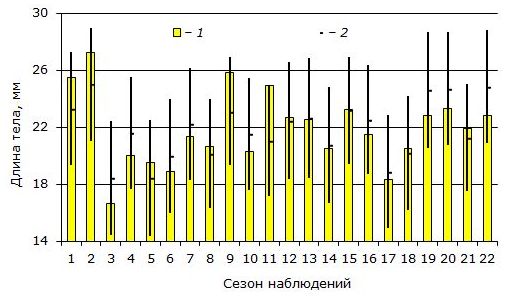
Рис. 2. Многолетняя вариация средних размеров сеголеток Rana temporaria после окончания периода нагула.
1 – наблюдаемые значения; 2 – рассчитанные значения, вертикальные линии соответствуют 95 % доверительному интервалу для средних значений
Fig. 2. Multiannual variations of the mean body length of Rana temporaria juveniles at the end of foraging period.
1 – observed data; 2 – predicted values computed using multiply regression method, with 95% confidence interval for means
Рассмотрим, как работают абиотические составляющие процесса роста сеголеток по отдельности и во взаимодействии. Примеры связей итоговых размеров сеголеток с отдельными переменными факторами внешней среды в изученных условиях показаны на рис. 3.
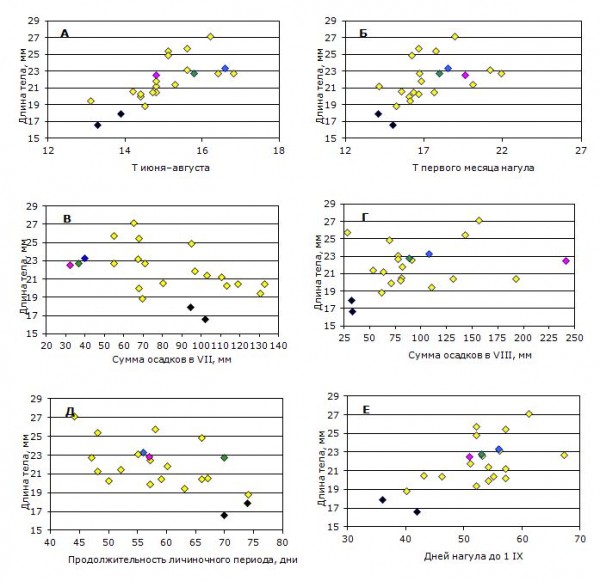
Рис. 3. Зависимости средних размеров сеголеток Rana temporaria к осени от характеристик периода нагула.
А — от средних температур июня – августа (rs = 0.84, p = 0.000) и Б – первых 30 дней нагула (rs = 0.64, p = 0.001); В — от количества осадков в июле (rs = -0.58, p = 0.004) и Г — в августе (rs = 0.13, недост.); Д – от продолжительности личиночного развития (rs = -0.53, p = 0.001) и Е — общей продолжительности нагула (rs = 0.50, p = 0.002). Зеленая метка соответствует генерации 2001 г., лиловая — 2003 г. и темно-голубая — генерации 2011 г.; черным цветом помечены генерации, имевшие минимальные размеры (1990 и 2008 гг.). Пояснения в тексте
Fig. 3. Dependences of mean body length of Rana temporaria juveniles on the parameters of foraging period by the beginning of autumn
А — on the mean air temperature in June – August (rs = 0.84, p = 0.000) and Б — on the mean air temperature of the first 30 days of their life on land (rs = 0.64, p = 0.001); В — on the amount of rainfall in July (rs = -0.58, p = 0.001) and Г — in August (rs = 0.13, n.s.); Д — from the duration of larval period (rs = -0.53, p = 0.001) and E — on the whole duration foraging (up to 1st of September). Green mark represents frog generation of 2001, purple — of 2003, and dark-blue — generation of 2011; black marks represent generation of 1990 and 2008, the least in size. Explanation is in the text.
Зависимость размеров сеголеток от количества осадков в июле и в августе оказалась противоположной по знаку (табл., рис. 3 В, Г). Следует отметить, что удаление из августовской выборки (рис. 3 Г) трех «отскакивающих» от общей тенденции точек увеличивает степень связи рассматриваемых переменных более чем вдвое (rs = 0.33, p = 0.16).
Отрицательная зависимость конечной длины тела сеголеток от обилия осадков в июле не является линейной: после засушливых периодов начала наземной жизни (сумма осадков менее 50 мм) сеголетки, вопреки общей тенденции, явно испытывали некоторое угнетение роста. В такие условия попали поколения 2001 (зеленая метка), 2003 (лиловая) и 2011 (темно-голубая метка) годов. Проследим за «приключениями» генераций этих лет (рис. 3).
Сеголетки поколения 2003 г. росли при средних значениях факторов «температура воздуха» (А, Б на рис. 3) и «продолжительность» (Д, Е), однако по количеству осадков в первую и вторую половины нагула этот сезон оказался контрастным и экстремальным. Август 2003 г. был самым дождливым за всю историю метеонаблюдений в «Киваче». За один из дней выпало 86.7 мм, а за месяц — 241.9 мм осадков. Средняя многолетняя сумма осадков за август — самый обильный на дожди месяц года — составляет 84.1 мм, а за год — 628.2 мм (Скороходова, 2008, обработка 1966–2005 гг.). Иными словами, только за одни сутки в августе 2003 г. воды выпало больше месячной нормы, а за весь месяц — почти 40 % нормы годовой. Тем не менее этот экстремум (напомним, в первую половину нагула обилие осадков сказывается отрицательно) никак не повлиял на размеры сеголеток: рассчитанные по уравнению и полученные в поле размеры сеголеток совпали до десятых долей мм (значения 22.5 и 22.5 мм). Их размеры близки таковым у генераций 2001 и 2011 гг.
Сеголетки поколения 2001 г. (зеленая метка) имели один из самых длительных периодов личиночного развития (рис. 3 Д), однако он был обусловлен очень ранним выклевом головастиков — 1 мая, при средней многолетней 11 мая (за 34 года наблюдений). Все остальные контролируемые параметры периода нагула в 2001 г. оказались для этих лягушат оптимальными, исключая засушливость июля.
Поколения 1990 и 2008 гг., имевшие к осени минимальные в нашем многолетнем ряду средние размеры, выделены на рис. 3 черными значками. Они росли, по сути, в пессимальных условиях. Сроки их личиночного развития сильно затянулись: в 2008 г. первые сеголетки появились только 25 июля (продолжительность личиночного периода 74 дня) при средней многолетней 6 июля, и для роста им оставалось всего 36 дней (недобор до средней многолетней продолжительности нагула составил 30 %). В 1990 г. выход начался 13 июля. Относительно благоприятные по количеству осадков условия июлей обошли лягушат этих двух генераций стороной, а короткий нагул пришелся в основном на прохладный август (рис. 3 Б) с крайне низким количеством осадков (рис. 3 Г) — фактически «июль» в жизни этих поколений пришелся на август и был крайне неблагоприятным по двум основным показателям, использованным в нашем анализе. Для сравнения укажем, что сеголетки генерации 1988 г., имевшие самые крупные средние размеры в нашей выборке — 27.2 мм, завершили личиночное развитие всего за 44 дня и имели второй по продолжительности период нагула (рис. 3 Е).
Обсуждение
Размеры ювенильных особей бесхвостых земноводных умеренной климатической зоны — один из важнейших показателей их жизнеспособности: чем крупнее сеголетки, тем устойчивее они к неблагоприятным внешним воздействиям и тем больше у них шансов дожить до зимы и пережить ее (Ищенко, Леденцов, 1985; Ляпков, 1986, 1997; Feldman, 1987; Harper, Semlitsch, 2007). Цепь зависимостей, приводящая к итоговому разнообразию размеров тела сеголеток у видов, развитие которых проходит в нестабильной обстановке временных водоемов, достаточно сложна.
Морфологическая изменчивость сеголеток таких видов закладывается еще на стадии ооцитов. Исходная вариабельность размеров яиц включает как генетическую, так и средовую составляющую. Из разных по размеру яиц появляются головастики, обладающие разными скоростями роста и развития, а непредсказуемые условия личиночного развития (температура, химизм, плотность населения головастиков и пр.) способны видоизменять ход этих процессов, что сказывается и на сроках метаморфоза, и на исходных размерах сеголеток (обзор см.: Ищенко, 1989).
Конечные размеры ювенильных особей перед зимовкой определяются их исходными размерами, вариабельностью скорости роста, избирательной смертностью в начале наземной жизни. Проведенные специальные оценки влияния размерных, плотностных и временных переменных на морфологическую изменчивость сеголеток R. temporaria, R. arvalis и R. sylvatica (Ищенко, Щупак, 1979; Смирина, 1980; Ищенко, Леденцов, 1985; Ляпков, 1986, 1988, 1989, 1995; Ляпков и др., 2000; Harper, Semlitsch, 2007) показали, что зависимость эта весьма замысловата. В тезисной форме ее можно выразить следующим образом.
1. На начальные размеры сеголеток существенно влияют как размах изменчивости скорости роста и темпа развития головастиков, так и сроки выхода лягушат на сушу (имеется в виду растянутость сроков выхода сеголеток данной генерации). Однако сеголетки, выходящие в разное время, могут иметь одинаковые размеры, а выходящие одновременно — разные.
2. Компенсационный рост, т. е. зависимость темпов роста животных одного возраста от размеров тела, ведущая к уменьшению разнообразия размеров, — ситуация вполне обычная (Мина, Клевезаль, 1976). Однако в ходе перечисленных исследований обнаружено, что скорость роста в период нагула может как зависеть, так и не зависеть от начальных размеров сеголеток (они растут с одинаковой скоростью). Скорость роста может зависеть или не зависеть от времени появления на суше. Конечные размеры сеголеток к осени существенно зависят (торможение роста) от исходной плотности лишь при ее уровне, превышающем некоторую критическую величину, когда снижается обеспеченность сеголеток пищей.
3. Смертность в начале наземной жизни может оказаться связанной со сроками выхода (чем позже появляются лягушата по сравнению с первыми, тем она выше), но не зависеть от исходных размеров или, наоборот, быть в значительной мере размерно-зависимой; авторы едины и категоричны во мнении, что смертность в наименьших размерных группах почти всегда повальна.
Вырисовывающаяся столь «разболтанная» во всех блоках-стадиях схема развития и роста является, с одной стороны, следствием отбора на максимальное разнообразие морфологии и физиологии сеголеток и, с другой — связана с нивелирующим эффектом действующих факторов нестабильной среды. Очевидно также, что максимально возможная разнокачественность личинок и ювенильных особей позволяет обеспечить пополнение популяции при любых флуктуациях местного климата.
Результаты нашего исследования позволяют сделать вывод, что в изученных условиях главенствующую роль в определении того, каковы будут конечные размеры сеголеток травяной лягушки, играют погодный и временной факторы.
В уравнение множественной регрессии c наибольшей степенью влияния вошла среднесуточная температура трёх летних месяцев. Объясняется это следующим. Чем теплее июнь — вторая половина периода личиночного развития — тем раньше начнется выход сеголеток, и тем дольше будет продолжаться нагул. Июль — это первые недели нагула, когда сеголетки растут тем быстрее, чем выше их температура тела (= температура приземного слоя воздуха). Для интенсификации роста лягушата проявляют практически круглосуточную активность (Залежский, 1938; Кутенков, 2009), и высокие температуры июля и августа являются необходимым подспорьем быстрого протекания метаболических реакций (зависимость конечных размеров тела сеголеток от среднесуточных температур этих месяцев положительна и достоверна, rs соответственно 0.56 и 0.47).
Дожди в изученных условиях имеют заметное, но явно второстепенное значение: территория заповедника «Кивач» относится к значительно увлажненным районам Карелии (Скороходова, 2008), т. е. летняя сырость здесь норма. Тем не менее как высокое обилие осадков, так и относительная сухость в первые недели жизни сеголеток неудобны для их роста. В августе, когда лягушата подросли, картина меняется (см. рис. 3Г).
Из использованных в анализе показателей сроков в уравнение регрессии вошла продолжительность личиночного периода (с отрицательным знаком), с которой длина тела сеголеток коррелировала несколько больше, чем с общей продолжительностью нагула (Д1 и Д3 в таблице). Однако сроки личиночного развития в наших условиях взаимосвязаны с продолжительностью периода нагула сеголеток отрицательной и почти функциональной зависимостью (rs = -0.91, p = 0.0000), что вполне естественно. Поэтому можно утверждать, и это также очевидно, что чем больше времени проведут на суше сеголетки до наступления холодов, тем бóльших размеров они достигнут.
Исследователи отмечают, что сокращение периода личиночного развития земноводных как правило однозначно сопровождается выходом более мелких сеголеток. Для R. temporaria данный эффект описан и при благоприятном ходе развития (Ляпков, 1986), и при преждевременном усыхании репродуктивных водоемов (Laurila et al., 2002). Это происходит потому, что скорость роста личинок возрастает медленнее, чем скорость их развития, и животные достигают стадии метаморфоза при меньших размерах тела (Мина, Клевезаль, 1976). В то же время показано, что когда сроки метаморфоза в данной популяции растянуты, сеголетки, выходящие последними и имеющие относительно крупную начальную длину, всё же к концу нагула не догоняют вышедших раньше своих более мелких ровесников (Ляпков, 1989). Основываясь на наших наблюдениях, можно утверждать, что как бы ни был растянут во времени метаморфоз, дата его начала в значительной степени определяет итоговые размеры сеголеток, какими бы сложными и замысловатыми не оказывались реальные траектории роста отдельных групп лягушат.
Заключение
Статистический анализ многолетнего ряда данных по варьированию длины тела сеголеток в конце периода нагула позволил установить следующее. Решающим фактором формирования межгодовой морфологической изменчивости в условиях флуктуирующих погодных характеристик оказывается действие температуры. Именно температура определяет продолжительность личиночного периода, а также длительность и условия нагула. Конечные размеры тела сеголеток перед зимовкой связаны обратной зависимостью с продолжительностью личиночной стадии и положительной — с временем нагула. Эффект осадков оказывается противоположным на разных отрезках времени наземной жизни.
Библиография
Ищенко В. Г. Репродуктивная тактика и демография популяций земноводных // Пробл. популяционной экологии земноводных и пресмыкающихся. Итоги науки и техники. Сер. зоология позвоночных / ВИНИТИ. М., 1989. Т. 17. С. 5–51.
Ищенко В. Г., Леденцов А. В. Экологические аспекты постметаморфического роста остромордой лягушки // Экологические аспекты скорости роста и развития животных. Свердловск: УНЦ АН СССР, 1985. С. 11–21.
Ищенко В. Г., Щупак Е. Л. Внутрипопуляционная изменчивость скорости роста и развития личинок остромордой лягушки (Rana arvalis Nilss) // Популяционные механизмы динамики численности животных. Свердловск: УНЦ АН СССР, 1979. С. 49–60.
Залежский Г. В. К динамике численности некоторых видов амфибий // Сборник трудов науч. студенческих кружков МГУ, 1938. Вып. 2. Биология. С. 3–28.
Кутенков А. П. О роли погодного фактора в динамике популяции травяной лягушки (Rana temporaria) на Севере // Экология, 1998. № 1. С. 49–54.
Кутенков А. П. Экология травяной лягушки (Rana temporaria) на Северо-Западе России . Петрозаводск, 2009. 140 с.
Ляпков С. М. Факторы, обусловливающие изменчивость размеров и сроков выхода сеголеток травяной (Rana temporaria) и остромордой (R. arvalis) лягушек // Зоол. журн., 1986. Т. 65, вып. 1. С. 88–97.
Ляпков С. М. Выживаемость сеголеток бурых лягушек (Rana temporaria и R. arvalis) в начале наземной жизни // Зоол. журн., 1988. Т. 67, вып. 10. С. 1519–1529.
Ляпков С. М. Изменчивость темпов роста бурых лягушек в начале их наземной жизни // Вопросы герпетологии: Материалы 7-й Всесоюзной герпетол. конф. Киев, 1989. С. 144–145.
Ляпков С. М. Факторы, обусловливающие гибель сеголеток травяной (Rana temporaria) и остромордой (R. arvalis) лягушек в начале наземной жизни // Зоол. журн., 1995. Т. 74, вып. 1. С. 92–106.
Ляпков С. М. Влияние размеров неполовозрелых травяных (Rana temporaria) и остромордых (R. arvalis) лягушек на их выживаемость во время зимовки // Зоол. журн., 1997. Т. 76, вып. 3. С. 356–363.
Ляпков С. В., Черданцев В. Г., Черданцева Е. М., Северцов А. С. Выживаемость и рост сеголеток бурых лягушек по мере их расселения от нерестового водоема // Зоол. журн., 2000. Т. 79, вып. 6. С. 729–741.
Мина М. В., Клевезаль Г. А. Рост животных. Анализ на уровне организма . М.: Наука, 1976. 291 с.
Скороходова С. Б. О климате заповедника «Кивач» // Труды Государственного природного заповедника «Кивач». Вып. 4. Петрозаводск, 2008. С. 3–34.
Смирина Э. М. О темпе роста и выживаемости травяных лягушек (Rana temporaria) в первые годы жизни // Зоол. журн., 1980. Т. 59, вып. 12. С. 1831–1840.
Feldman R. Wanderdynamik und gewichtsentwicklung junger Grasfrösche (Rana temporaria) // Natur und heimat. 1987. V. 47. № 2. S. 45–52.
Harper E., Semlitsch R. Density dependence in the terrestrial life history stage of two anurans // Oecologia. 2007. V. 153. P. 879–889.
Kutenkov A., Mosiyash S. On the dynamics of population of Common frog (Rana temporaria) in the North-West of Russia // Russian j. of gerpetol. 2000. V. 7. № 2. P. 123–134.
Laurila A., Karttunen S., Merilä J. Adaptive phenotypic plasticity and genetics of larval life histories in two Rana temporaria populations // Evolution. 2002. V. 56. № 3. P. 617–627.

 © 2011 - 2026
© 2011 - 2026