



Введение
В настоящее время промысловая добыча диких животных окончательно уступила место охотничьему трофейному отстрелу. Охотничий трофейный туризм, особенно в странах СНГ, является одним из наиболее интенсивно развивающихся и престижных форм использования природных ресурсов и, как отрасль хозяйственной деятельности человека, требует рационального подхода и должен регулироваться на научных данных по численности и половозрастной структуре популяции трофейных видов. Одними из наиболее ценных и желанных объектов трофейной охоты являются различные виды копытных, среди которых горные бараны, занимающие одно из ведущих мест.
Данное исследование, выполненное в условиях Казахского мелкосопочника, является частью цикла работ по изучению состояния популяций отдельных подвидов горных баранов в странах Средней и Центральной Азии, где практикуется трофейная охота на архаров. Практическая направленность исследований определялась необходимостью разработки долгосрочной программы по рациональному использованию и управлению их популяциями в условиях нарастающего влияния различных форм антропогенных воздействий на популяцию. Наряду с практическими целями полученные данные имеют выраженный теоретический аспект и необходимы для познания особенностей формирования и функционирования отдельных составляющих популяционных структур, что практически не делалось для таких редких и малоизученных видов, как горные бараны. Известно, что структурная организация популяции формируется под влиянием различного рода внешних и внутренних воздействий и отражает интегрированный ответ популяции на эти воздействия. Поэтому она выступает в качестве одной из важнейших общепопуляционных адаптаций, обеспечивающих оптимальный режим функционирования и в конечном итоге устойчивость популяций в сложных и динамичных условиях среды.
Именно половозрастная и пространственно-временная структура популяций животных и их динамика на фоне средовых изменений – наименее изученный аспект популяционных регуляций. В мировой экологической литературе нет ни одной обобщающей сводки по данной проблеме, хотя бы в пределах одного класса животных. Рядом авторов на примере отдельных видов рассматривалась лишь взаимная зависимость изменений полового состава популяций и их численности, часто с весьма противоречивыми результатами.
Общая цель работы заключалась в оценке текущего состояния и выявлении закономерностей динамики структурно-функциональной организации популяции казахстанского архара в зависимости от пространственно-временной динамики средовых факторов, а также различных форм и интенсивности антропогенных воздействий.
Под структурно-функциональной организацией популяции мы понимаем относительно устойчивое во времени соотношение половых и возрастных групп в популяции, определенным образом распределенных в пространстве и связанных между собой в единое функциональное целое (Магомедов и др., 2001). В качестве параметров структурно-функциональной организации популяции казахстанского архара рассматриваются особенности изменения размеров и состава стад отдельных популяционных групп, пространственно-временной характер использования ими территории, а также половой и возрастной состав и особенности изменения этих и других параметров популяций в зависимости от конкретных условий среды обитания.
Материалы
Бараны, населяющие Казахский мелкосопочник, большинством исследователей отнесены к казахстанской или североказахстанской форме архаров (Ovis ammon collium, Severtzov, 1873) (Цалкин, 1951; Бербер, Калмыков, 1994; Байдавлетов, 1996; Бекенов и др., 1999; Федосенко, 2000). Казахстанский архар обитает в Центральном и Восточном Казахстане, где занимает большую часть Казахского нагорья, на юге идет до гор Бектаута и Катань-Эмель (северное Прибалхашье), восточнее, в Семипалатинской области, он обычен в горах Карачингиз, Акчатау и др. В Восточном Казахстане водится в Калбинском Алтае, Тарбагатае и Сауре. Его ареал простирается и за пределы Казахстана.
Районы проведения наших исследований приходились на центральную часть Казахского мелкосопочника (Центральноказахстанский мелкосопочник) на территорию Карагандинской и частично Семипалитинской областей. Участки сбора полевого материала охватывали горные массивы Кызылтау, Калмаккырган, Аркалык, Едрей, Мыржик. Климат резко континентальный, выражается в суровости зимы, высоких летних температурах, малой продолжительности весны и осени, больших годовых и суточных амплитудах температуры воздуха, сухости воздуха, малом количестве осадков (Альпатьев и др., 1976). Количество осадков сильно колеблется по годам, составляя в среднем около 300 мм в год. Лучшим увлажнением выделяются горные кряжи, на которых выпадает за год около 400 мм осадков. Основное количество осадков приходится на летний период (70–80 %), поэтому зимы преимущественно малоснежные (около 20–25 см). Для зимнего периода характерны частые и сильные метели (до 40 дней) (Гвоздецкий, Михайлов, 1978).
Преимущественное распространение имеют типчаково-ковыльные степи, местами значительную роль в травостое могут играть более ксерофитный ковыль тырсик (Stipa sareptana), тонконог (Koeleria gracilis), овсец (Helictotrichon desertorum). Весной развиваются эфемеры и эфемероиды, а по западинам и балкам встречаются заросли степных кустарников – чилиги и спиреи (Гвоздецкий, Михайлов, 1978). В горных массивах мелкосопочника проявляется высотная зональность. Наблюдаются различия в травостое в зависимости от экспозиции склона. Северные склоны хорошо представлены разнотравьем, тогда как южные – более ксерофитными видами с узкими и довольно жесткими листьями.
В связи с сильным сокращением поголовья домашнего скота за последние годы пастбищная нагрузка на растительность в районе проведения исследований слабо выражена. Следствием этого является накопление ветоши и степные пожары, охватывающие зачастую огромные территории. Пожары сильно сказываются на кормовой базе архаров в зимний, наиболее тяжелый в кормовом отношении, период. Судя по опросным данным, на территории процветает также браконьерство.
В период исследований (с 4 по 18 ноября 2002 г.) визуально было отмечено 449 особей архара на территории площадью 1544 км2.
Методы
В качестве основного метода учета численности по ряду причин был выбран маршрутный метод с использованием автомобилей. Подсчеты животных проводились в светлое время суток одновременно на трех автомобилях по заранее определенным суточным маршрутам. В ряде случаев параллельно использовались учетные площадки, которые закладывались через каждые 3–5 км по основной линии маршрута (трансекты).
Общее количество отмеченных животных и их половых и возрастных групп, места их обнаружения, направление движения и время обнаружения наносились на план или крупномасштабную карту местности и фиксировались в памяти электронных носителей.
При изучении половой и возрастной структуры популяций выделялись самки, самцы и сеголетки обоих полов. Определенные трудности возникали при определении полуторагодовалых особей, в этом возрасте самцов и самок очень трудно отличить на расстоянии. Поэтому при сборе эмпирического материала мы их условно относили к самкам, а в дальнейшем они разделялись при анализе демографической структуры. В связи с тем что рога самок небольшие по размерам и плохо просматриваются на расстоянии, эмпирические данные по количеству животных старших возрастов получены только для самцов. Благодаря годовым кольцам на роговых чехлах и форме рогов возраст самцов даже в природе определялся с точностью до одного года.
Для выяснения возрастной структуры популяции также использовались рога погибших животных. По кольцам на роговых чехлах можно безошибочно определить возраст не только самцов, но и самок, и даже время года, когда животное погибло. Нами были обнаружены и осмотрены 37 черепов самцов и всего 2 черепа самок. Использован также материал по погибшим животным (черепа самцов), который хранится в музее Академии наук Казахстана (74 черепа). Этот метод широко используется при анализе возрастной структуры полорогих (Коли, 1979; Риклефс, 1979; Пианка, 1981). Кроме того, нами был получен материал и по возрастам отстреленных животных (всего 126 животных, из которых 2 самки), однако при анализе возрастной структуры мы его не использовали, т. к. отстрел носит избирательный по полу и возрасту характер и поэтому не отражает реальной демографической структуры в популяции. Незначительное количество черепов самок и молодых, видимо, связано с тем, что их более легкие черепа растаскиваются хищниками.
Выравнивание возрастных рядов и построение возрастного распределения в популяции архаров (по полу и возрасту) рассчитывались на основе специальных логарифмическо-полиминальных и интегральных уравнений (Ахмедов, Магомедов, 2000; Магомедов и др., 2001).
Пространственно-временную структуру и характер использования территории популяцией архара изучали на основе материала, полученного в результате анализа встреч животных в зависимости от характера местообитания. В качестве отдельных характеристик местообитаний рассматривались высота над уровнем моря, ориентация склонов относительно солнца, а также отдельные элементы рельефа – долины, края долин и подножия склонов, сами склоны и платообразные поднятия с гребнями хребтов.
В качестве критериев защитных свойств местообитаний баранов использовали данные по общему характеру рельефа, степени его расчлененности (количество балок на единице площади), средней крутизне склонов, их протяженности, соотношении выровненных (пологих участков) и изрезанных склонов, их взаиморасположении и др.
Результаты
Как показали наши исследования, распределение архаров по территории было относительно равномерным. Плотность популяции по отдельным горным массивам существенно не различалась и колебалась в пределах 0.24–0.35 особи на 1 км2, составляя в среднем 0.28 ± 0.02 ос./км2 (табл. 1).
Таблица 1. Плотности населения архаров на участках проведения исследований
| № | Участки проведения исследований (горный массив) | Количество отмеченных животных, (n) | Площадь проведения учетов, (км2) | Плотность популяции, (ос./км2) |
| 1 | Кызылтау | 101 | 315 | 0.32 |
| 2 | Калмаккырган | 48 | 184 | 0.26 |
| 3 | Аркалык | 25 | 103 | 0.24 |
| 4 | Едрей | 96 | 342 | 0.28 |
| 5 | Мыржик | 74 | 296 | 0.25 |
| 6 | Коныртемирши | 105 | 304 | 0.35 |
| Всего | 449 | 1544 | 0.28 |
Тем не менее в распределении животных по территории выявлены определенные закономерности. Хотя в период проведения исследований архары отмечались нами и в открытых совершенно ровных межгорных долинах, большинство животных все же придерживалось мелкосопочников и выходов горных пород на поверхность, т. е. территорий с расчлененным рельефом. Отмечены прямые зависимости плотности архаров от площади и высоты горных массивов (рис. 1). Чем больше площадь горного массива и чем он выше, тем большей протяженностью и крутизной характеризуются склоны и резче выражена сеть оврагов. Очевидно, это связано с тем, что на пересеченной территории архары находят наиболее оптимальные кормовые и защитные условия.
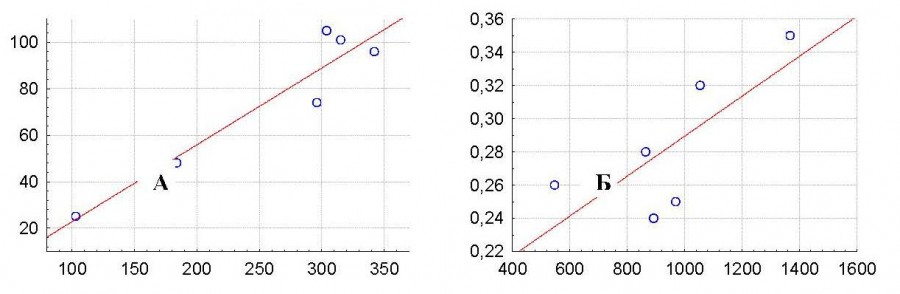
Рис. 1. Зависимость количества отмеченных животных от площади горного массива (r = 0.97; P < 0.05) (А) и высоты над уровнем моря (r = 0.75; P > 0.05) (Б). По оси абсцисс: А – площадь горного массива (км2); Б – отметки абсолютных высот горного массива (м); по оси ординат: А – количество отмеченных животных (особей); Б – плотность населения архаров (ос/км2)
Fig. 1. The dependence of the number of marked animals on the area of the mountain range (r = 0.97; P < 0.05) (A) and altitude (r = 0.75; P > 0.05) (B). The X-axis: A – the area of the mountain range (km2); B – markы of the absolute heights of the mountain range (m); the Y-axis: A – the number of marked animals (individuals); B – the population density of argali (individuals/km2)
Отмеченные зависимости численности и плотности от площади горного массива и степени расчлененности территории, помимо кормового фактора, во многом определяются большими возможностями защиты от хищников и человека. На более расчлененных и больших по площади горных образованиях, т. е. менее проходимых для автотранспорта, численность популяции выше, чем на более доступных для человека участках. На роль пересеченного рельефа как защитных условий местообитаний баранов указывает и поведение животных, которые при опасности всегда устремлялись к ближайшим горным массивам.
Составной частью социальной структуры копытных, во многом определяющей тип и эффективность функционирования популяций, является стадность. В целом индекс стадности в популяции архаров на территории проведения исследований в период гона составил 4.24 ± 0.30. Наибольшее количество животных в период проведения исследований сосредоточено в гаремах (39.2 %), затем в смешанных (31.4 %) и самочьих группах (23.4 %). Высокий индекс стадности характерен для смешанных групп, а для самочьих групп и гаремов он значительно ниже (табл. 2).
Таблица 2. Типы групп архаров и индексы их стадности
| Типы групп | Количество групп | Количество особей | Индекс стадности | ||
| N | % | n | % | ||
| Одиночки | 25 | 23.6 | 25 | 5.6 | 1.0 |
| Самочьи | 24 | 22.6 | 105 | 23.4 | 4.38 ± 0.61 |
| Самцовые | 1 | 0.9 | 2 | 0.4 | 2.0 |
| Гаремы | 36 | 34.0 | 176 | 39.2 | 4.89 ± 0.38 |
| Смешанные | 20 | 18.9 | 141 | 31.4 | 6.81 ± 0.72 |
| Всего | 106 | 449 | 4.24 ± 0.30 | ||
Наибольшее количество животных было сосредоточено в группах размером от 3 до 8 особей. В целом в группах такого размера держатся 65.7 % животных популяции. Самая большая группа архаров, отмеченная нами на данной территории, состояла из 17 животных (рис. 2).
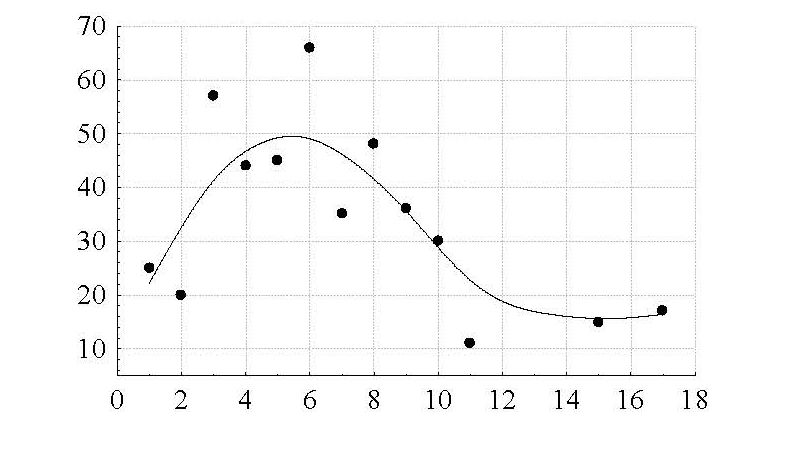
Рис. 2. Распределение казахстанского архара по группам в зависимости от размеров групп. По оси абсцисс – размер групп; по оси ординат – количество особей в группах
Fig. 2. The distribution of Kazakhstan argali into groups depending on group sizes. The X-axis – the size of groups; the Y-axis – the number of individuals in groups
Показано, что среди множества возможных факторов индекс стадности зависит от двух параметров: высоты горного массива над уровнем моря и относительного количества самцов в популяции. С увеличением высоты горного массива, косвенно отражающей защищенность местообитания, индекс стадности закономерно падает (r = -0.85; P < 0.05), что характерно для многих горных копытных (Wilson, 1975; Баскин, 1976; Коли, 1979; Дарман, 1990). Другим параметром, влияющим на индекс стадности архаров, является относительное количество самцов в популяции: в период гона самцы отбивают самок от основных групп и образуют гаремы, характеризующиеся низкими индексами стадности. Оставшиеся без гаремов одиночные самцы также снижают общие показатели индекса стадности (рис. 3).
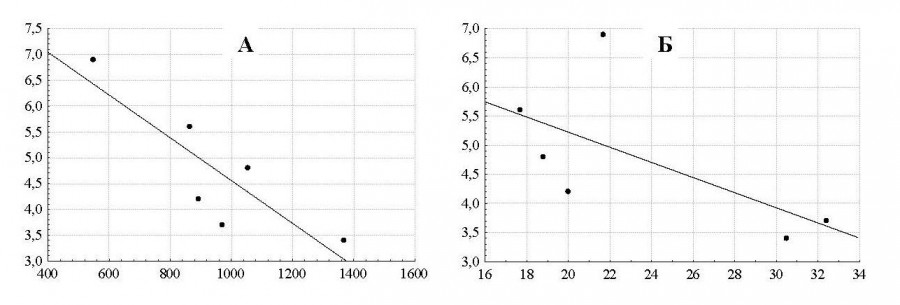
Рис. 3. Зависимость индекса стадности популяции казахстанского архара от высоты местообитания (А): Y = 8.701 - 0.004x; (r = -0.85; P < 0.05) и относительного количества самцов в популяции (Б): Y = 7.822 - 0.13x; (r = -0.63; P > 0.05). По оси абсцисс: А – высота над уровнем моря (метры), Б – относительное количество самцов в популяции (%); по оси ординат – индекс стадности
Fig. 3. The dependence of Kazakhstan argali herd population index on habitat height (A): Y = 8.701 - 0.004x; (r = -0.85; P < 0.05) and relative number of males in the population (B): Y = 7 822 - 0.13 x; (r = -0.63; P > 0.05). The X-axis: A – altitude (meters), B – the relative number of males in the population (%); the Y-axis – herd index
Известно, что для архаров, как и для большинства горных копытных, характерны половозрастные различия в использовании территории, которые наиболее выражены в летний период. Период проведения наших исследований совпал с периодом гона. И в это время большинство животных находятся в гаремах и смешанных группах, и по характеру использования территории самцы и самки практически не различаются. На территории проведения исследований практически все поголовье архаров держалось мелкосопочников – 412 особей (91.8 %), в межгорных долинах отмечено всего 37 особей (8.2 %). Из животных, отмеченных в мелкосопочниках, большая часть встречена на склонах гор (78.6 %), значительно меньше – в межгорных ложбинах (17.5 %) и совсем мало – на гребнях сопок (3.9 %).
На склонах горных массивов (мелкосопочников) нами было отмечено 324 архара, из которых 43.8 % придерживалось северных склонов, 29.0 % – склонов восточной экспозиции, 14.5 и 12.7 % соответственно – западных и южных склонов. Предпочтение архарами склонов северных экспозиций, видимо, объясняется доминированием здесь мезофитных видов растений, хорошо поедаемых баранами.
Архары встречались в диапазоне высот от 400 до 1300 м н. у. м., где основное их поголовье (83.3 %) придерживалось высоты 550–850 м н. у. м., а максимальное количество встреченных животных приходилось на высоту 650 м н. у. м. (19.6 %). Эти высоты соответствуют средней части склонов и занимают наибольшие площади на территории проведения исследований.
Таким образом, в начале зимнего периода наиболее предпочитаемыми участками архаров являются средние части склонов северных экспозиций горных массивов. Важнейшими факторами, определяющими пространственную структуру популяции, являются орографические условия местообитаний, характер распределения растительного покрова, а также метеорологические условия данного периода.
Важнейшая популяционная характеристика животных – их половая и возрастная структура, отражающая текущее состояние вида в целом и отдельных его популяций, интенсивность их размножения и смертности по различным возрастным группам. Как и у многих других видов, и особенно у копытных, она выступает и в качестве основного параметра популяции, который необходимо учитывать при ее эксплуатации. Поэтому при проведении исследований мы старались уделить особое внимание половозрастному составу рассматриваемой популяции и их групп.
К сожалению, в период проведения исследований гон у архаров уже начался, поэтому на одной и той же территории можно было встретить как самочьи, так и самцовые группы, но большинство из отмеченных групп были смешанными.
Как видно из табл. 3, в популяции архаров практически на всех горных массивах центральной части Казахского мелкосопочника преобладают взрослые самки; они составляют от 36.0 до 52.1 % от всей популяции (в среднем 44.28 ± 3.07). Количество самцов старше 2 лет и сеголеток примерно равное (23.52 ± 2.58 % и 21.00 ± 3.82 % соответственно). Наименьшей половозрастной группой являются годовалые животные (9.33 ± 0.62 %). Половое соотношение среди взрослых особей (старше 2 лет) составляет 1.9:1.0 в пользу самок, т. е. в среднем в популяции на одного самца приходятся около 2 самок. Доля сеголеток на одну самку в целом составляет 0.43.
Таблица 3. Половозрастная структура популяции архаров на отдельных горных массивах центральной части Казахского мелкосопочника
| Район исследований (горный массив) | Самки | Сеголетки | Годовалые | Самцы | Всего | ||||
| n | % | n | % | N | % | n | % | ||
| 1. Кызылтау | 52 | 51.1 | 19 | 18.8 | 11 | 10.9 | 19 | 18.8 | 101 |
| 2. Калмаккырган | 18 | 37.5 | 13 | 21.7 | 4 | 8.3 | 13 | 21.7 | 48 |
| 3. Едрей | 9 | 36.0 | 9 | 36.0 | 2 | 8.0 | 5 | 20.0 | 25 |
| 4. Аркалык | 50 | 52.1 | 19 | 19.8 | 10 | 10.4 | 17 | 17.7 | 96 |
| 5. Мыржик | 37 | 50.0 | 5 | 6.8 | 8 | 10.8 | 24 | 32.4 | 74 |
| 6. Коныртемирши | 41 | 39.0 | 24 | 22.9 | 8 | 7.6 | 32 | 30.5 | 105 |
Анализ половой и возрастной структуры популяций в зависимости от высоты массива над уровнем и от общей площади горного массива выявил некоторые связи между этими параметрами. Относительное количество самок и годовалых показывает положительную связь с площадью горного массива (r = 0.80; P > 0.05 – для самок; r = 0.62; P > 0.05 – для годовалых), тогда как количество сеголеток – отрицательную (r = -0.70; P > 0.05). Таким образом, в течение первого года жизни на таких массивах незначительно выше (r = 0.40; P > 0.05).
Хотя половозрастная сегрегация самок и самцов – явление широко распространенное, слабое проявление отмеченных выше закономерностей, скорее всего, связано с периодом гона, когда самцы горных баранов бродят по территории и не проявляют привязанности к определенной территории.
Для анализа демографической структуры популяции нами были использованы материалы по черепам погибших животных (111 черепов самцов в возрасте от 1 до 11 лет) и данные визуальных наблюдений. Анализ показывает, что доля самцов в популяции начинает резко снижаться с 4-летнего возраста и сходит на нет к 15 годам, тогда как снижение доли самок в популяциях идет гораздо медленнее, а в популяции встречаются самки 18 лет (рис. 4).
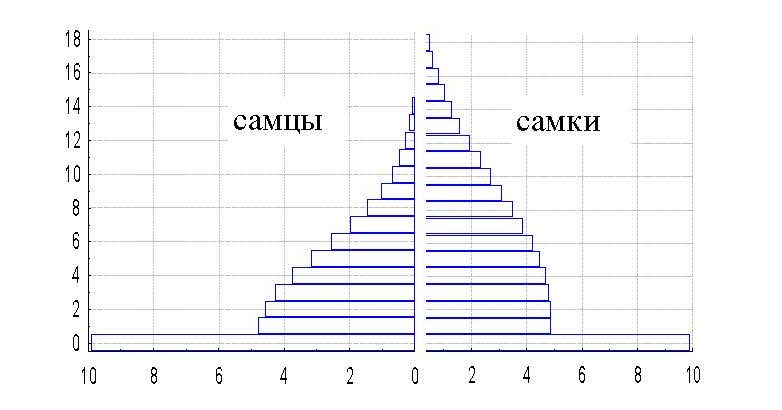
Рис. 4. Возрастная пирамида популяции казахстанского архара. По горизонтали – количество животных (%); по вертикали – возраст (годы)
Fig. 4. Age pyramid of the population of Kazakhstan argali. The X-axis – the number of animals, %; the Y-axis –age (years)
Отмеченные различия распределения самок и самцов по возрастным кластерам являются результатом различий в их удельной смертности в разные возрастные периоды (рис. 5). Относительно высокие показатели удельной смертности характерны для сеголеток и животных старших возрастных групп, наименьшие – для молодых животных в возрасте 1–4 года. Выделяются более высокие показатели удельной смертности самцов начиная с 3-летнего возраста, что может объясняться преимущественным легальным или нелегальным отстрелом самцов охотниками. Согласно опросным данным, на данной территории при трофейной охоте добывается не менее 100 особей животных в год в возрасте от 7 до 14 лет.
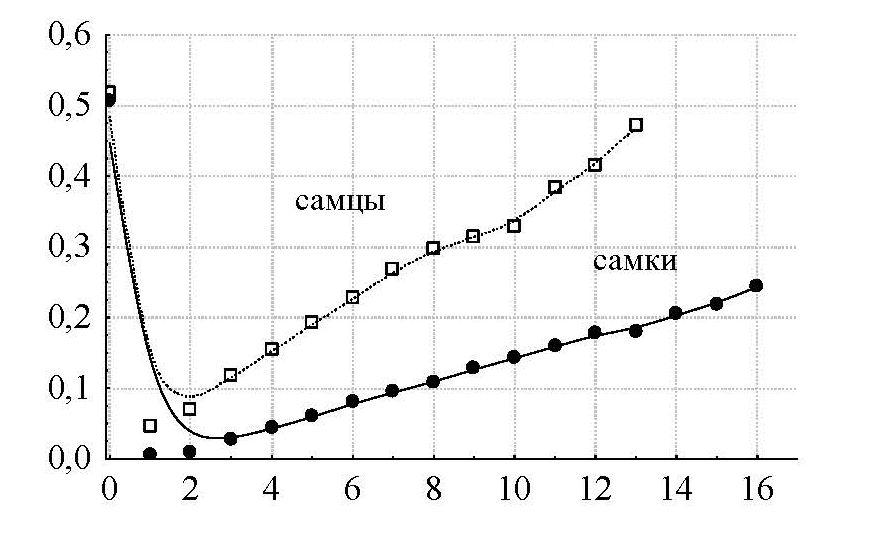
Рис. 5. Удельная смертность самок и самцов в популяциях архаров в зависимости от возраста в центральной части Казахского мелкосопочника. По оси абсцисс – удельная смертность; по оси ординат – возраст (годы)
Fig. 5. Specific mortality of females and males in argali populations depending on age in the Central part of the Kazakh upland. X-axis – specific mortality; Y-axis – age (years)
Заключение
В связи с сильным сокращением поголовья домашнего скота за последние годы пастбищная нагрузка на растительность в районе проведения исследований практически не выражена. Существенное влияние на популяции архаров, видимо, уже оказывают степные пожары, охватывающие зачастую огромные территории и лишающие животных зимних кормов.
Одним из основных факторов, ограничивающих численность популяций, несомненно, является браконьерский отстрел животных. Степень влияния браконьерства на численность популяции зависит от орографических особенностей местообитаний, определяющих защитные условия территории. Надо сказать, что эволюционно приуроченность животных к территориям с защитными условиями сформировалась как ответная реакция на деятельность хищников. В современных условиях, хотя фактор хищничества продолжает сохранять определенное значение в распределении животных, на первый план выходят антропогенные факторы, а именно браконьерство. Эти две причины, определяющие гибель животных, имеют много общего в том плане, что степень их влияния на популяции прямо определяется степенью защищенности территории. Однако, если при нападении хищников даже небольшие защитные элементы ландшафтов (отдельные сопки, скалы или даже скальные останцы) позволяют животным избежать гибели, то при воздействии со стороны человека к защитным условиям предъявляются более жесткие требования (удаленность от населенных пунктов и автодорог, труднопроходимость для человека и автотранспорта, хороший обзор и т. д.). Следует также отметить, что характер элиминации особей копытных при воздействии со стороны хищников носит более «доступный» характер, чаще всего связанный с тем, что хищники изымают из популяции дефектных, больных или старых особей. Во время браконьерской охоты присутствует слабая избирательность и явная избирательность по крупным самцам-производителям при трофейной охоте.
Исследованная нами популяция казахстанского архара по своей природе и характеру занимаемых местообитаний всегда представлялась наиболее уязвимой по сравнению с другими подвидами горных баранов Центральной Азии, где большую роль в ограничении численности играет их доступность для охотников.
Согласно литературным данным, численность архаров в пределах Казахского мелкосопочника до середины 1960-х гг. была высокой и стабильной и составляла около 16000 особей. Затем началось снижение численности, и к 1970 г. она уменьшилась более чем наполовину (около 7000 особей) (Савинов, 1974). Процесс снижения численности продолжался и в последующие годы. В 1980-х гг. насчитывалось уже 5000 особей (Федосенко, Капитонов, 1983). Первая половина 1990-х гг. также характеризовалась снижением численности (2082–2428), а к концу 1990-х гг. она едва превышала 1000 экз. (1997 г.), хотя на отдельных участках плотность продолжала оставаться высокой (Бекенов, Байдавлетов, 1997; Бекенов и др., 1999; Федосенко, 2000). Резкое снижение численности казахстанского архара со второй половины 1960-х до начала 1990-х гг. было связано исключительно с хозяйственной деятельностью человека – интенсификацией сельского хозяйства в этом регионе (освоением целины и ростом поголовья домашних животных). После 1990-х гг. – беспрецедентным ростом браконьерства на фоне распада СССР и утраты законопослушания, тяжелым материальным и финансовым положением природоохранных организаций, появлением высокоэффективного нарезного оружия у населения. Все более заметную роль стали играть и такие факторы, как резкое увеличение количества волков, ежегодные степные пожары, лишающих баранов зимних кормов, проведение трофейных охот в период гона и т. д.
Все это находит подтверждение и в наших исследованиях: отмечена четкая положительная связь между численностью и плотностью архаров, с одной стороны, и площадью горного массива и степенью расчлененности территории – с другой. На более расчлененных и больших по площади горных массивах, т. е. менее проходимых для автотранспорта, численность популяции выше, чем на более доступных для человека участках. На роль пересеченного рельефа как защитных условий местообитаний баранов указывает и поведение животных: хотя мы встречали животных и в открытой степи, увидев нас, они всегда устремлялись к ближайшим горным массивам.
В популяции архаров практически на всех горных массивах центральной части Казахского мелкосопочника преобладают взрослые самки, составляющие от одной трети до половины всей популяции. Количество самцов старше 2 лет в среднем не превышало 25 % от всей популяции. Доля самцов начинает резко снижаться с 4-летнего возраста и сходит на нет к 15 годам. Естественной определяющей причиной изменений в демографической структуре популяции являются кормовые условия зимнего и летнего периодов. Ранее нами было показано, что летние кормовые условия определяют упитанность копытных к началу зимнего периода (Ахмедов, Магомедов, 1995, 2000). Самцы в период гона, который у большинства видов копытных проходит в начале зимы, теряют практически все жировые запасы, накопленные в летне-осенний период. В результате этого выживаемость их оказывается полностью зависимой от обеспеченности кормами в зимний период. Самки в период гона практически не теряют жировые запасы, и выживаемость их в зимний период зависит от кормовых условий не только зимнего, но и летнего периода. В итоге в зимний период, когда доступность и питательность кормов резко снижается, выживаемость самцов оказывается ниже, чем у более упитанных самок. Дополнительным подтверждением этого является снижение доли самцов по направлению с юга на север. На северных окраинах ареала из-за более глубокого снежного покрова доступность кормов в зимний период ниже, чем в южных районах. Мы считаем, что у горных баранов именно это является одной из причин, определяющих половозрастную структуру популяции. На это накладываются и последствия степных пожаров, после которых большие территории оказываются практически непригодными для зимовки животных. Пожары в северных районах охватывают большие площади благодаря лучшему развитию здесь степной растительности. К тому же, покидая гари, животные вынуждены использовать участки, которые были сохранены под зимние пастбища домашних животных и где, помимо конкуренции с овцами, они становятся более доступными для волков и охотников.
По литературным данным, к 2000-м гг. существенно возросла и роль волка в смертности архаров на территории Казахского мелкосопочника (Бекенов, Байдавлетов, 1997; Бекенов и др., 1999; Федосенко, 2000). По данным А. К. Федосенко (2000), на долю волков в эти годы приходилось от 40 до 73 % всех погибших животных.
На рассматриваемой территории проводится трофейный отстрел животных, в период с 1990 по 2001 г. было отстреляно 124 самца в возрасте от 7 до 14 лет. Таким образом, наряду с доступностью и сезонной динамикой кормовых условий (пожары, выпас скота, климатические условия, давление волков и др.), в качестве нового фактора, оказывающего влияние на демографическую структуру популяций, на данной территории выступает избирательный трофейный отстрел животных.
Библиография
Альпатьев А. М., Архангельский А. М., Подоплелов Н. Я., Степанов А. Я. Физическая география СССР (Азиатская часть) . М.: Высш. шк., 1976. 357 с.
Ахмедов Э. Г., Магомедов М.-Р. Д. Зависимость динамики поло-возрастной структуры популяций дагестанского тура (Capra cylindricornis Blyth) от условий обитания и плотности популяций // Зоологический журнал. 1995. Т. 74. Вып. 7. С. 109–118.
Ахмедов Э. Г., Магомедов М.-Р. Д. Закономерности формирования демографической структуры популяции дагестанского тура (Capra cylindricornis) // Зоологический журнал. 2000. Т. 79. Вып. 4. С. 461–470.
Байдавлетов Р. Ж. Современное состояние популяции архара в Центральном Казахстане // Состояние териофауны в России и ближнем зарубежье. М., 1996. С. 21–26.
Баскин Л. М. Поведение копытных животных . М.: Наука, 1976. 293 с.
Бекенов А. Б., Байдавлетов Р. Ж. Научный эксперимент «архар» // Редкие виды млекопитающих России и сопредельных стран. М., 1997. С. 10.
Бекенов А. Б., Байдавлетов Р. Ж., Федосенко А. К., Вейнберг П. И. О состоянии популяции архара в Карагандинской области // Проблемы охраны и устойчивого использования биоразнообразия животного мира Казахстана. Алматы, 1999. С. 13–14.
Бербер А. П., Калмыков И. Б. Архар в Карагандинской области // Охота и охотничье хозяйство. 1994. № 11. С. 12–14.
Гвоздецкий Н. А., Михайлов Н. И. Физическая география СССР (Азиатская часть) . М.: Мысль, 1978. 508 с.
Дарман Ю. А. Стадность косули как популяционный показатель // V съезд Всесоюзного териологического общества АН СССР (29 янв. – 2 февр. 1990 г., Москва): Тез. докл. М., 1990. Т. 2. С. 141–142.
Коли Г. Анализ популяций позвоночных . М.: Мир, 1979. 362 с.
Магомедов М.-Р. Д., Ахмедов Е. Г., Яровенко Ю. А. Дагестанский тур (популяционные и трофические аспекты экологии) . М.: Наука, 2001. 138 с.
Пианка Э. Эволюционная экология . М.: Мир, 1981. 396 с.
Риклефс Р. Основы общей экологии . М.: Мир, 1979. 424 с.
Савинов Е. Ф. Результаты авиаучета архара в Казахском нагорье // Современное состояние и пути развития охотоведческой науки в СССР. Киров, 1974. С. 168–169.
Федосенко А. К. Архар в России и сопредельных странах . М., 2000. 291 с.
Федосенко А. К., Капитонов В. И. Архар // Млекопитающие Казахстана. Алма-Ата, 1983. Т. 3. Ч. 3. С. 144–209.
Цалкин В. И. Горные бараны Европы и Азии . М.: Изд-во МОИП, 1951. 343 с.
Wilson E. O. Sociobiology: The new synthesis. Cambridge: Harvard Univ. Press, 1975. 696 p.
Благодарности
Автор выражает свою искреннюю благодарность охотоведам и егерям Карагандинской области Казахстана, Тарасенко Анатолию Николаевичу, Даровских Сергею Николаевичу, Ботову Вадиму Иннокентьевичу, Туганову Денису Евгеньевичу и др., которые способствовали успешному выполнению данной работы.
Особая благодарность своему ученику и коллеге Ахмедову Эльдару Гасановичу, а также Субботину Андрею Евгеньевичу, обеспечившему экспедицию финансовой поддержкой от Фонда Международного сафари-клуба (Safari Club International Foundation USA).

 © 2011 - 2026
© 2011 - 2026