



Введение
Среди насекомых-фитофагов, населяющих леса, большая часть видов имеет очень низкую плотность популяций, когда ущерб кормовым растениям наносится очень редко или вообще отсутствует. Но есть группа видов, численность популяций которых может периодически в несколько лет изменяться на 5–7 порядков. Большая часть лесных насекомых с периодическими изменениями численности популяций относится к группе хвое- и листогрызущих насекомых и вызывает сильную дефолиацию кормовых растений, но редко – гибель. Однако массовые размножения хвое- и листогрызущих насекомых и ослабление леса приводят к росту уровня численности стволовых вредителей – насекомых, питающихся тканями коры и древесины. В Сибири очаги стволовых вредителей часто образуются после вспышек массового размножения сибирского коконопряда. Насекомые – стволовые вредители ускоряют процесс гибели растения и древостоев в целом.
Целью настоящей работы является выявление пространственной и временной синхронизации развития вспышек массового размножения комплекса видов насекомых-ксилофагов в лесах Красноярского края. Этот анализ необходим для оценки рисков возникновения вспышек и оптимизации мониторинга насекомых-ксилофагов, повреждающих лесные насаждения. Знание пространственной сопряженности развития вспышек отдельного вида ксилофагов позволяет при обнаружении очага массового размножения этого вида в отдельном лесничестве оценить риск развития его вспышек на других территориях. Знание временной сопряженности развития вспышек нескольких видов ксилофагов в отдельном лесничестве позволяет при обнаружении очагов массового размножения одного вида в этом лесничестве дать оценку рисков возникновения очагов размножения других видов ксилофагов.
Материалы
Данные по повреждению лесных насаждений и информация о вредителях леса были собраны в ходе наземных текущих лесопатологических обследований лесных насаждений сотрудниками Центра защиты леса Красноярского края в различных лесничествах в течение 2007–2015 гг. В процессе лесопатологического обследования производилось определение границ повреждений леса; учет численности вредителей; оценка степени повреждения насекомыми древостоев. Лесопатологическое обследование лесов, заселенных стволовыми вредителями, проводилось сотрудниками путем визуального осмотра ослабленных участков леса. Заселенность насаждений стволовыми вредителями оценивалась по наличию усохших и усыхающих деревьев, по увяданию хвои в кроне, наличию на коре буровой муки, ползающих жуков, входных и вылетных отверстий. Насаждение, в котором количество заселенных стволовыми вредителями деревьев превышает 10 %, считается очагом стволовых вредителей (Лесная энтомология, 2010).
В ходе проведения лесопатологических обследований в лесном фонде Красноярского края были обнаружены очаги массового размножения 15 видов ксилофагов, в частности таких видов стволовых вредителей, как златка бронзовая Dicerca aenea L., златка лиственничная Melanophila guttulata Gebl. (Coleoptera, Buprestidae), усач черный еловый большой Monochamus urussovi Fisch., усач черный еловый малый Monochamus sutor L., усач черный сосновый Monochamus galloprovincialis Ol., скрипун большой осиновый Saperda carcharias L. (Coleoptera, Cerambycidae), лубоед пальцеходный Xylechinus pilosus Ratz., лубоед малый сосновый Tomicus minor Hartig., лубоед большой сосновый Tomicus piniperda L., лубоед большой еловый (дендроктон) Dendroctonus micans Kug., полиграф уссурийский (белопихтовый) Polygraphus proximus Blandf., заболонник березовый Scolytus ratzeburgi Jans., короед шестизубый (стенограф) Ips sexdentatus Boern., короед большой лиственничный (продолговатый) Ips subelongatus Motsch., короед-типограф Ips typographus L. (Coleoptera, Scolytidae).
В табл. 1 приведены сведения о площади очагов стволовых вредителей в лесных насаждениях на территории лесничеств Красноярского края (неопубликованные материалы ежегодных отчетов Центра защиты леса Красноярского края).
Таблица 1. Площади (в га) очагов насекомых-ксилофагов на территории лесничеств Красноярского края по годам
Table 1. Areas (ha) of damage to forest stands by insects-xylophages in the territory of Krasnoyarsk Region
| № | Вид насекомых-ксилофагов | Лесничество | Площади (в га) очагов насекомых-ксилофагов по годам | |||||||
| 2007 | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | |||
| 1 | Полиграф уссурийский (белопихтовый) | Ачинское | 0 | 0 | 753.5 | 753.5 | 863.5 | 1009.4 | 1681.4 | 984.5 |
| 2 | Усач черный еловый большой | Ачинское | 0 | 13.7 | 13 | 13 | 13 | 13 | 13 | 13 |
| 3 | Усач черный еловый большой | Балахтинское | 0 | 41.5 | 35.3 | 33.2 | 0 | 0 | 0 | 0 |
| 4 | Лубоед большой сосновый | Боготольское | 0 | 0 | 0 | 54 | 0 | 0 | 0 | 0 |
| 5 | Полиграф уссурийский | Боготольское | 0 | 0 | 474.6 | 474.6 | 474.6 | 474.6 | 143.3 | 115.3 |
| 6 | Лубоед пальцеходный | Боготольское | 0 | 54 | 54 | 54 | 54 | 54 | 0 | 0 |
| 7 | Усач черный еловый большой | Боготольское | 0 | 373.8 | 350.8 | 350.8 | 350.8 | 350.8 | 25 | 25 |
| 8 | Лубоед малый сосновый | Богучанское | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 9 | Усач черный еловый малый | Богучанское | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 10 | Полиграф уссурийский | Большемуртинское | 0 | 0 | 0 | 0 | 0 | 0 | 475 | 475 |
| 11 | Полиграф уссурийский | Большеулуйское | 0 | 0 | 0 | 0 | 0 | 0 | 676.3 | 676.3 |
| 12 | Короед продолговатый | Борское | 50 | 17.5 | 17.5 | 17.5 | 0 | 0 | 0 | 0 |
| 13 | Усач черный еловый большой | Верхне-Манское | 0 | 659 | 659 | 609 | 609 | 609 | 609 | 609 |
| 14 | Короед шестизубый | Верхне-Манское | 20 | 20 | 66 | 979 | 979 | 979 | 933 | 933 |
| 15 | Златка бронзовая | Гремучинское | 45 | 45 | 0 | 0 | 0 | 0 | 0 | 0 |
| 16 | Короед продолговатый | Гремучинское | 41 | 41 | 41 | 5.4 | 5.4 | 188.1 | 78.4 | 30.2 |
| 17 | Короед шестизубый | Гремучинское | 423 | 213.6 | 213.6 | 213.6 | 213.6 | 213.6 | 213.6 | 213.6 |
| 18 | Лубоед малый сосновый | Гремучинское | 298 | 297.8 | 297.8 | 23.1 | 15.2 | 15.2 | 15.2 | 80 |
| 19 | Лубоед большой сосновый | Гремучинское | 39 | 25.1 | 25.1 | 0 | 23.1 | 259 | 121.3 | 48.6 |
| 20 | Усач черный еловый малый | Гремучинское | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| 21 | Златка лиственничная | Гремучинское | 0 | 0 | 45 | 45 | 45 | 45 | 45 | 96.3 |
| 22 | Скрипун осиновый | Гремучинское | 0 | 0 | 0 | 0 | 0 | 35 | 35 | 12.8 |
| 23 | Лубоед пальцеходный | Гремучинское | 0 | 0 | 0 | 15.2 | 0 | 0 | 0 | 0 |
| 24 | Усач черный еловый большой | Гремучинское | 0 | 3287.4 | 3279.6 | 3241.2 | 3241.2 | 3241.2 | 3203.2 | 3203.2 |
| 25 | Заболонник березовый | Дзержинское | 0 | 0 | 0 | 0 | 0 | 0 | 22 | 0 |
| 26 | Усач черный еловый большой | Дзержинское | 0 | 0 | 0 | 0 | 0 | 2 | 2 | 2 |
| 27 | Лубоед малый сосновый | Дивногорское | 16 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 28 | Усач черный еловый большой | Долгомостовское | 0 | 95.3 | 95.3 | 205.3 | 110 | 110 | 110 | 110 |
| 29 | Короед продолговатый | Долгомостовское | 24 | 24 | 24 | 24 | 0 | 0 | 0 | 0 |
| 30 | Заболонник березовый | Долгомостовское | 1 | 0.7 | 0.7 | 0.7 | 0 | 0 | 0 | 0 |
| 31 | Полиграф уссурийский | Емельяновское | 0 | 0 | 0 | 0 | 0 | 0 | 109.2 | 1785 |
| 32 | Усач черный еловый большой | Енисейское | 0 | 10081 | 10144 | 1711 | 2227 | 2227 | 831 | 831 |
| 33 | Усач черный еловый большой | Ермаковское | 0 | 3280.4 | 3280.4 | 3280.4 | 3280.4 | 3280.4 | 3140 | 3140 |
| 34 | Короед шестизубый | Ирбейское | 206 | 206 | 206 | 206 | 206 | 206 | 206 | 206 |
| 35 | Лубоед большой сосновый | Ирбейское | 21 | 21 | 21 | 0 | 21 | 12 | 12 | 12 |
| 36 | Лубоед малый сосновый | Ирбейское | 0 | 0 | 0 | 21 | 0 | 0 | 0 | 0 |
| 37 | Усач черный еловый большой | Ирбейское | 0 | 35508 | 35508 | 35508 | 35508 | 27250 | 26520 | 0 |
| 38 | Усач черный еловый большой | Казачинское | 0 | 7750 | 150 | 150 | 0 | 0 | 0 | 0 |
| 39 | Короед шестизубый | Каратузское | 0 | 0 | 0 | 0 | 0 | 295 | 295 | 295 |
| 40 | Усач черный еловый большой | Каратузское | 0 | 4065 | 4065 | 3434 | 3434 | 3567 | 1732 | 1732 |
| 41 | Короед шестизубый | Кизирское | 0 | 0 | 0 | 5.8 | 5.8 | 5.8 | 5.8 | 5.8 |
| 42 | Усач черный еловый большой | Кизирское | 0 | 798 | 2961.1 | 2669.3 | 2650.9 | 2496.2 | 2496.2 | 2048.2 |
| 43 | Короед продолговатый | Кодинское | 84 | 84 | 84 | 84 | 84 | 84 | 84 | 84 |
| 44 | Короед шестизубый | Кодинское | 31 | 23.4 | 23.4 | 23.4 | 23.4 | 23.4 | 23.4 | 23.4 |
| 45 | Лубоед большой сосновый | Кодинское | 1580 | 1580 | 1580 | 0 | 1580 | 1580 | 1580 | 1529.8 |
| 46 | Заболонник березовый | Кодинское | 0 | 92 | 92 | 92 | 92 | 92 | 92 | 92 |
| 47 | Златка бронзовая | Кодинское | 0 | 360 | 0 | 0 | 0 | 0 | 0 | 0 |
| 48 | Златка лиственничная | Кодинское | 0 | 0 | 360 | 360 | 360 | 360 | 360 | 360 |
| 49 | Лубоед сосновый малый | Кодинское | 0 | 0 | 0 | 1580 | 0 | 0 | 21.2 | 21.2 |
| 50 | Усач черный еловый большой | Кодинское | 0 | 2391 | 2391 | 2391 | 2391 | 2391 | 2293 | 2578.9 |
| 51 | Полиграф уссурийский | Козульское | 0 | 0 | 724.5 | 884.2 | 887.5 | 505.4 | 1196.3 | 1174.6 |
| 52 | Усач черный еловый большой | Козульское | 0 | 3.9 | 3.9 | 3.9 | 3.9 | 3.9 | 0 | 0 |
| 53 | Лубоед сосновый малый | Красноярское | 30 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 54 | Лубоед большой сосновый | Красноярское | 17 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 55 | Полиграф уссурийский | Красноярское | 0 | 0 | 0 | 0 | 0 | 0 | 366.3 | 366.3 |
| 56 | Усач черный еловый большой | Красноярское | 0 | 89.5 | 84 | 84 | 0 | 0 | 0 | 0 |
| 57 | Усач черный еловый большой | Маганское | 0 | 83 | 83 | 82.5 | 82.5 | 82.5 | 82.5 | 82.5 |
| 58 | Заболонник березовый | Манзенское | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 59 | Короед продолговатый | Манзенское | 21 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 60 | Лубоед еловый большой | Манзенское | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 61 | Усач черный еловый большой | Манзенское | 0 | 67.1 | 48.5 | 22 | 22 | 22 | 22 | 22 |
| 62 | Златка бронзовая | Манское | 123 | 123 | 103 | 54 | 54 | 54 | 8 | 8 |
| 63 | Короед шестизубый | Манское | 978 | 1084.4 | 1025.9 | 1291 | 1305.9 | 1231.9 | 1181.9 | 1280.9 |
| 64 | Златка лиственничная | Манское | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 |
| 65 | Полиграф уссурийский | Манское | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5.1 |
| 66 | Усач черный еловый большой | Манское | 0 | 574.2 | 571.4 | 598.1 | 583.2 | 583.2 | 583.2 | 583.2 |
| 67 | Усач черный еловый малый | Манское | 0 | 0 | 0 | 14.9 | 0 | 0 | 0 | 0 |
| 68 | Полиграф белопихтовый | Мининское | 0 | 0 | 0 | 0 | 0 | 0.4 | 707.22 | 749.1 |
| 69 | Усач черный еловый большой | Мининское | 0 | 8.1 | 8.1 | 8.1 | 0 | 0 | 0 | 0 |
| 70 | Усач черный еловый большой | Мотыгинское | 0 | 42143.6 | 40644.6 | 40361.6 | 40361.6 | 40361.6 | 35780.6 | 15342 |
| 71 | Короед типограф | Назаровское | 27 | 16.2 | 0 | 0 | 0 | 0 | 0 | 0 |
| 72 | Полиграф уссурийский | Назаровское | 0 | 0 | 0 | 0 | 0 | 0 | 1522.4 | 1470.4 |
| 73 | Короед продолговатый | Невонское | 4 | 4.2 | 4.2 | 4.2 | 4.2 | 4.2 | 4.2 | 4.2 |
| 74 | Лубоед сосновый малый | Невонское | 119 | 98.9 | 86.3 | 21.4 | 49.9 | 49.9 | 161.9 | 123.9 |
| 75 | Лубоед большой сосновый | Невонское | 39 | 39.4 | 39.4 | 0 | 21.4 | 21.4 | 21.4 | 21.4 |
| 76 | Заболонник березовый | Невонское | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| 77 | Златка лиственничная | Невонское | 0 | 0 | 0 | 0 | 0 | 0 | 245.5 | 245.5 |
| 78 | Лубоед пальцеходный | Невонское | 0 | 0 | 0 | 49.9 | 0 | 55 | 55 | 55 |
| 79 | Усач черный еловый большой | Невонское | 0 | 104.9 | 104.9 | 104.9 | 104.9 | 104.9 | 137.9 | 137.9 |
| 80 | Короед шестизубый | Пировское | 87 | 87 | 87 | 87 | 87 | 87 | 87 | 70 |
| 81 | Короед типограф | Пировское | 1125 | 1207.6 | 1052.6 | 1052.6 | 1052.6 | 1052.6 | 869.5 | 731.2 |
| 82 | Лубоед малый сосновый | Пировское | 43 | 72.6 | 72.6 | 48 | 59.6 | 59.6 | 59.6 | 59.6 |
| 83 | Лубоед большой сосновый | Пировское | 31 | 48 | 48 | 0 | 48 | 48 | 48 | 34.4 |
| 84 | Заболонник березовый | Пировское | 97 | 97 | 97 | 97 | 97 | 97 | 97 | 0 |
| 85 | Полиграф белопихтовый | Пировское | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 3 |
| 86 | Лубоед пальцеходный | Пировское | 0 | 0 | 0 | 59.6 | 0 | 0 | 0 | 0 |
| 87 | Усач черный еловый большой | Пировское | 0 | 5570.2 | 5532.7 | 5532.7 | 5532.7 | 5532.7 | 5343.2 | 3442 |
| 88 | Лубоед пальцеходный | Саяно-Шушенское | 1 | 1 | 1 | 53.9 | 0 | 0 | 0 | 0 |
| 89 | Златка бронзовая | Саянское | 16 | 16.3 | 0 | 0 | 0 | 0 | 0 | 0 |
| 90 | Лубоед малый сосновый | Саянское | 107 | 82.4 | 53.9 | 0 | 53.9 | 53.9 | 53.9 | 53.9 |
| 91 | Златка лиственничная | Саянское | 0 | 0 | 16.3 | 16.3 | 16.3 | 16.3 | 16.3 | 16.3 |
| 92 | Усач черный еловый большой | Саянское | 0 | 1.2 | 0 | 0 | 0 | 0 | 0 | 0 |
| 93 | Усач черный еловый большой | Северо-Енисейское | 0 | 330.5 | 330.5 | 309.2 | 25.9 | 25.9 | 25.9 | 5.9 |
| 94 | Усач черный еловый большой | Саяно-Шушенское | 0 | 956 | 956.1 | 956.1 | 0 | 0 | 956.1 | 956.1 |
| 95 | Усач черный еловый малый | Таёжинское | 73 | 68.9 | 24.9 | 24.9 | 20 | 20 | 20 | 20 |
| 96 | Полиграф уссурийский | Таёжинское | 0 | 0 | 0 | 0 | 0 | 0 | 275 | 355 |
| 97 | Усач черный еловый большой | Таёжинское | 0 | 2252.7 | 2135.2 | 1953.9 | 1310.4 | 1289.7 | 1281.4 | 1281.4 |
| 98 | Короед продолговатый | Терянское | 0 | 0 | 0 | 0 | 0 | 0 | 93.2 | 93.2 |
| 99 | Лубоед большой сосновый | Терянское | 0 | 0 | 0 | 0 | 0 | 0 | 70 | 70 |
| 100 | Усач черный еловый большой | Терянское | 0 | 759 | 759 | 759 | 759 | 759 | 654 | 654 |
| 101 | Короед продолговатый | Тунгуссо-Чунское | 48 | 30.7 | 30.7 | 30.7 | 30.7 | 0 | 30.7 | 30.7 |
| 102 | Короед шестизубый | Тунгуссо-Чунское | 74 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 103 | Лубоед малый сосновый | Тунгуссо-Чунское | 0 | 0 | 0 | 27.1 | 0 | 0 | 0 | 0 |
| 104 | Лубоед большой сосновый | Тунгуссо-Чунское | 0 | 28.6 | 27.1 | 0 | 27.1 | 27.1 | 27.1 | 27.1 |
| 105 | Усач черный еловый большой | Тунгуссо-Чунское | 0 | 0.6 | 0.6 | 0.6 | 0.6 | 0.6 | 0.6 | 0.6 |
| 106 | Усач черный еловый малый | Тюхтетское | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| 107 | Полиграф уссурийский | Тюхтетское | 0 | 0 | 0 | 0 | 0 | 38.3 | 306.3 | 278 |
| 108 | Усач черный еловый большой | Тюхтетское | 0 | 173 | 173 | 173 | 173 | 173 | 172.5 | 172.5 |
| 109 | Короед продолговатый | Ужурское | 7 | 7 | 7 | 7 | 7 | 7 | 7 | 7 |
| 110 | Усач черный еловый большой | Ужурское | 0 | 35 | 35 | 35 | 35 | 35 | 35 | 35 |
| 111 | Златка лиственничная | Усинское | 0 | 0 | 0 | 0 | 0 | 91 | 91 | 91 |
| 112 | Короед шестизубый | Усинское | 0 | 0 | 0 | 0 | 0 | 209 | 209 | 209 |
| 113 | Усач черный еловый большой | Усинское | 0 | 0 | 0 | 0 | 0 | 1499 | 1499 | 1499 |
| 114 | Усач черный еловый большой | Усольское | 0 | 1666 | 1666 | 39 | 39 | 39 | 39 | 39 |
| 115 | Лубоед малый сосновый | Уярское | 14 | 14 | 14 | 0 | 0 | 0 | 0 | 0 |
| 116 | Усач черный еловый большой | Уярское | 0 | 346.5 | 346.5 | 0 | 0 | 0 | 0 | 0 |
| 117 | Лубоед малый сосновый | Хребтовское | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 118 | Лубоед большой сосновый | Хребтовское | 0 | 84 | 84 | 84 | 84 | 84 | 84 | 84 |
| 119 | Усач черный еловый большой | Хребтовское | 0 | 768 | 768 | 768 | 717 | 717 | 717 | 717 |
| 120 | Лубоед малый сосновый | Чунское | 291 | 281 | 281 | 0 | 0 | 0 | 0 | 0 |
| 121 | Усач черный еловый малый | Чунское | 3 | 1.5 | 1.5 | 1.5 | 1.5 | 21.5 | 21.5 | 21.5 |
| 122 | Заболонник березовый | Чунское | 61 | 0 | 0 | 0 | 0 | 4 | 4 | 4 |
| 123 | Лубоед еловый большой | Чунское | 0 | 0 | 0 | 0 | 0 | 0 | 31 | 31 |
| 124 | Усач черный еловый большой | Чунское | 0 | 1552 | 0 | 0 | 956.1 | 956.1 | 17 | 17 |
Методы
В любом сообществе, состоящем из большого числа видов, маловероятно, чтобы какой-то экологический фактор оказывал влияние лишь на один вид из всего сообщества. Действие экологического фактора и одновременная реакция на него нескольких видов может говорить о наличии взаимосвязей между видами, о возможной соединенности в комплекс в сообществе, о наличии сопряженности популяционной динамики. Эта сопряженность выявляется в виде корреляции между количественными характеристиками популяций видов. Выделяют два основных типа сопряженности: временная сопряженность нескольких видов в одном местообитании и пространственная сопряженность популяционной динамики одного вида в разных местообитаниях. Временная сопряженность проявляется в том, что динамика численности разных видов комплекса может протекать синхронно в одном местообитании, как, например, подъемы численности сосновой пяденицы и комплекса взаимосвязанных с этим видом насекомых-филлофагов в Краснотуранском бору на территории Красноярского края (Пальникова и др., 2014). Пространственную сопряженность отдельного вида можно охарактеризовать, например, числом лесничеств, в которых наблюдаются вспышки его массового размножения в данный момент времени.
При изучении пространственной сопряженности некоторого вида k в N = 62 местообитаниях (лесничествах) в течение М = 8 лет использовалась для анализа матрица A(k)=||aijk|| размерности (N x M), в которой в ячейке (i, j) представлены данные о площади повреждения насаждения видом k в i-м лесничестве в j-м году. Такие матрицы строились для каждого из Р = 15 зарегистрированных видов ксилофагов.
При изучении временной сопряженности Р видов в местообитании m используется матрица B(m)=||bijm|| размерности (Р х М), в которой в ячейке (i, j) представлены данные по показателю повреждения насаждений в этом местообитании видом i в год j. Такие матрицы строились для каждого из N = 62 лесничеств.
15 матриц A(k) использовались для оценки пространственной сопряженности популяционной динамики отдельного вида насекомого в разных местообитаниях (лесничествах). Для расчетов пространственной сопряженности использовались значения коэффициентов корреляции между строками матриц А (k) для каждого из 15 изученных видов ксилофагов. В результате таких расчетов для всех 15 видов ксилофагов были получены корреляционные матрицы пространственной сопряженности размером (N x N), в которых отдельная ячейка (i, j) характеризовала пространственную сопряженность в течение 2007–2014 гг. развития вспышек массового размножения отдельного вида ксилофагов в лесничествах i и j. Если абсолютное значение коэффициента корреляции в ячейке было значимо согласно стандартным статистическим критериям, это указывало на пространственную сопряженность (в случае отрицательного значения коэффициента корреляции – антисопряженность) развития очагов отдельного вида в двух лесничествах i и j. Положительное значение коэффициента корреляции в ячейке корреляционной матрицы указывало на то, что при появлении очага массового размножения отдельного вида в одном лесничестве подобный же очаг появляется в другом лесничестве. При отрицательном значении коэффициента корреляции появление очага в одном лесничестве было связано с отсутствием очага изучаемого вида в другом лесничестве.
62 корреляционные матрицы B(m) использовались для оценки временной сопряженности массового размножения разных видов ксилофагов в одном лесничестве. Для расчетов использовались значения коэффициентов корреляции между строками матриц B(m). Если коэффициент корреляции был положительным, это указывало на то, что присутствие одного вида из комплекса ксилофагов являлось фактором, оказывающим влияние на появление другого вида, т. е. наличие временной сопряженности в динамике численности между разными видами ксилофагов в одном местообитании (лесничестве). Если коэффициент корреляции оказывался отрицательным, это указывало на то, что наличие в очаге одного вида не оказывало влияние на появление другого вида.
Для расчета корреляционных матриц использовался статистический пакет Statistica 6.0. При расчете оценивалась статистическая значимость каждого коэффициента корреляции в корреляционных матрицах на уровне р = 0.90. Если отдельный коэффициент корреляции в корреляционной матрице оказывался незначимым на выбранном уровне р, то из этого делался вывод об отсутствии пространственной синхронности повреждений насаждений вредителями одного вида в разных местообитаниях или временной синхронности повреждений насаждений вредителями нескольких видов в одном местообитании.
Результаты
Временная синхронизация характеризует ситуацию, когда на одной территории наблюдаются вспышки массового размножения нескольких видов. Для ксилофагов такой эффект может быть связан с изменением в состоянии кормовых древесных растений на территории. Различные лесничества на территории Красноярского края различаются по количеству зарегистрированных в них очагов массового размножения ксилофагов. «Рекордсменом» по числу очагов разных видов ксилофагов является Гремучинское лесничество, на территории которого в течение 2007–2014 гг. наблюдались очаги массового размножения 11 видов ксилофагов. Также большое число очагов разных видов стволовых вредителей наблюдалось на территориях Кодинского, Пировского, Манского и Невонского лесничеств. В 11 лесничествах наблюдались очаги массового размножения только одного вида; в 16 лесничествах вспышек массового размножения ксилофагов за 2007–2014 гг. не отмечалось.
Распределение лесничеств по количеству в них очагов массового размножения различных видов ксилофагов приведено на рис. 1. Почти в двух третях (65.6 %) от общего числа лесничеств (61) количество очагов ксилофагов не превосходит 2.
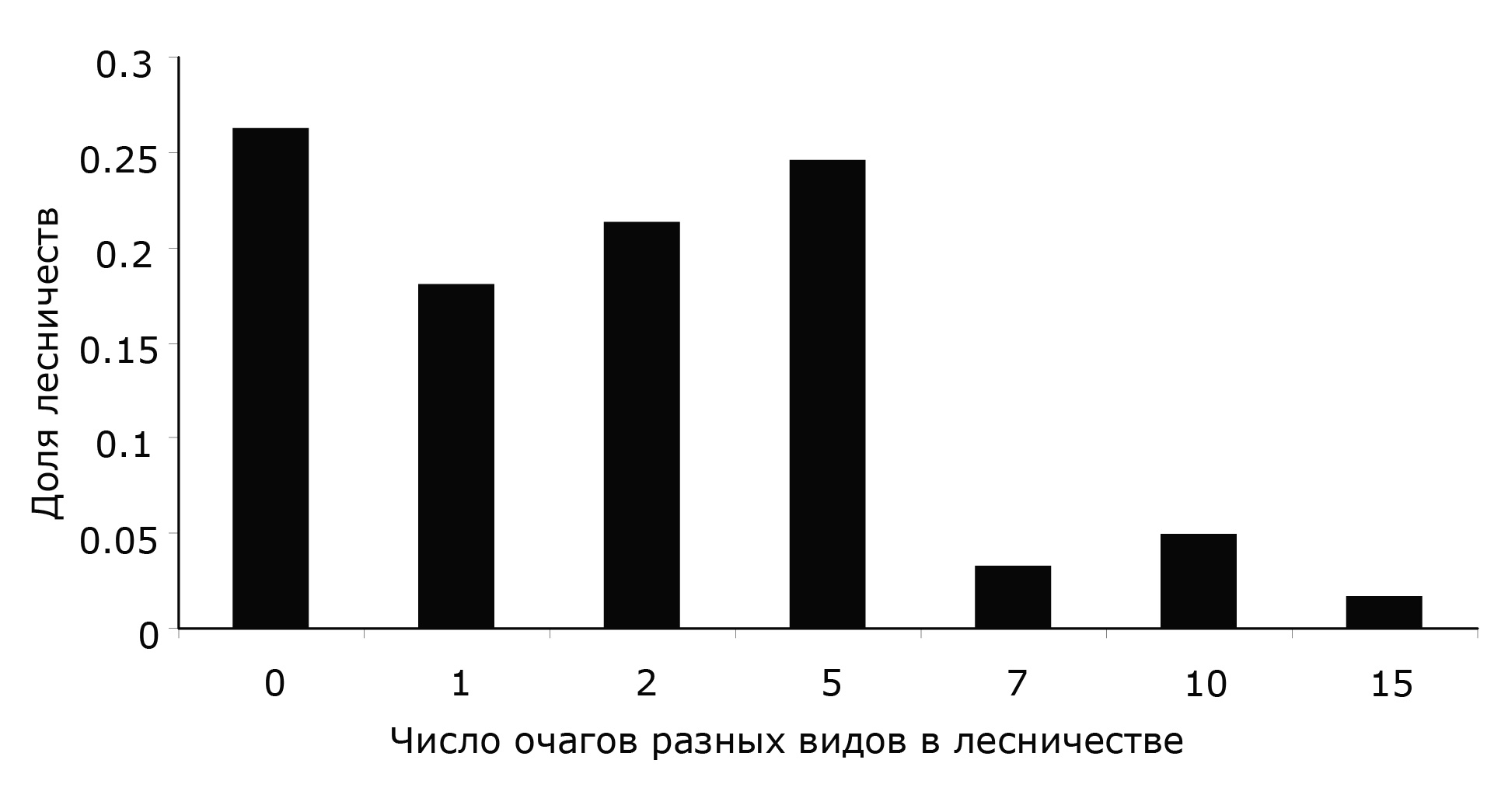
Рис. 1. Распределение лесничеств на территории Красноярского края по числу в них очагов массового размножения различных видов ксилофагов
Fig. 1. Distribution of forest areas in Krasnoyarsk region by the number of outbreaks of various xylophages species
Распределение очагов массового размножения ксилофагов на территории Красноярского края зависит от природно-климатических условий, в которых расположены леса. В табл. 2 приведены характеристики встречаемости очагов стволовых насекомых в зависимости от лесорастительной зоны, в которой расположено лесничество (Государственный доклад…, 2018).
Таблица 2. Встречаемость очагов стволовых насекомых в зависимости от лесорастительной зоны, в которой расположено лесничество
Table 2. Occurence of outbreaks of various types of xylophages depending on the zone in which the forest area is situated
| Лесорастительная зона | Всего лесничеств | Число лесничеств с зарегистрированными очагами массового размножения | Доля лесничеств с очагами массового размножения | Среднее число зарегистрированных очагов массового размножения на лесничество |
| Таежная зона | 17 | 12 | 0.71 | 4.75 |
| Лесостепная зона | 36 | 28 | 0.78 | 2.79 |
| Южносибирская горная зона | 9 | 5 | 0.55 | 2.00 |
Доля лесничеств с очагами массового размножения стволовых вредителей для Таежной зоны составляет 71 %; для Лесостепной зоны – 78 %, для Южносибирской горной зоны – 55 %.
Во всех трех лесорастительных зонах среднее число зарегистрированных очагов массового размножения ксилофагов не меньше 2. Для Таежной зоны это 4.75, для Лесостепной зоны – 2.79, для Южносибирской горной зоны – 2. Различные виды насекомых-ксилофагов очень сильно различаются по площади территорий, на которых возникали и действовали очаги их массового размножения (табл. 3).
Таблица 3. Распределение видов насекомых-ксилофагов по максимальной годичной площади (га) очагов массового размножения на территории Красноярского края
Table 3. Maximum annual area (ha) of xylophage foci in the territory of Krasnoyarsk Region
| Ранг вида | Вид насекомых-ксилофагов | Максимальная годичная площадь очагов, га |
| 1 | Усач черный еловый большой | 153085.0 |
| 2 | Полиграф уссурийский (белопихтовый) | 8437.6 |
| 3 | Усач черный сосновый | 7050.3 |
| 4 | Короед шестизубый | 3250.7 |
| 5 | Лубоед большой сосновый | 2031.5 |
| 6 | Короед-типограф | 1223.8 |
| 7 | Лубоед малый сосновый | 928.0 |
| 8 | Златка лиственничная | 809.1 |
| 9 | Короед большой лиственничный (продолговатый) | 314.0 |
| 10 | Заболонник березовый | 218.0 |
| 11 | Златка бронзовая | 184.0 |
| 12 | Усач черный еловый малый | 87.0 |
| 13 | Лубоед пальцеходный | 55.0 |
| 14 | Скрипун большой осиновый | 35.0 |
| 15 | Лубоед большой еловый | 31.0 |
Из данных табл. 3 видно, что размеры очагов массового размножения разных видов ксилофагов существенно различались. Так, максимальная годичная площадь очагов массового размножения Monochamus urussovi (153085 га) почти в 5000 (!) раз превосходит максимальную годичную площадь очагов Dendroctonus micans (31 га).
В качестве примера расчета временной сопряженности появления очагов массового размножения различных видов ксилофагов в одном лесничестве в табл. 4 приведены данные по площади поврежденных лесных насаждений (га) на территории Гремучинского лесничества (данные выделены из табл. 1). В ходе лесопатологических обследований лесных насаждений в течение 2007–2014 гг. в Гремучинском лесничестве отмечены 9 видов ксилофагов. Массовое размножение лубоеда пальцеходного отмечено один раз, при этом площадь поврежденных им насаждений составила чуть более 15 га. Скрипун осиновый в массе размножался в течение 2012–2014 гг., повреждая древостои на небольшой площади. Ежегодно в насаждениях Гремучинского лесничества отмечаются очаги короеда шестизубого, лубоеда малого соснового и усача черного елового большого (начиная с 2008 г.). Комплексные очаги массового размножения ксилофагов формируются в результате подъема численности одновременно 5–6 видов насекомых. Начиная с 2012 г. в очагах одновременно регистрируются 7 видов ксилофагов (см. табл. 4).
Таблица 4. Площади очагов (га) различных видов стволовых насекомых на территории Гремучинского лесничества
Table 4. Areas (ha) of outbreak foci of various species of insects-xylophages on the territory of the Gremuchinsky forestry
| Вид насекомых-ксилофагов | Площадь очагов (га) стволовых насекомых по годам | |||||||
| 2007 | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | |
| Златка бронзовая | 45 | 45 | 0 | 0 | 0 | 0 | 0 | 0 |
| Короед продолговатый | 41 | 41 | 41 | 5.4 | 5.4 | 188.1 | 78.4 | 30.2 |
| Короед шестизубый | 423 | 213.6 | 213.6 | 213.6 | 213.6 | 213.6 | 213.6 | 213.6 |
| Лубоед сосновый малый | 298 | 297.8 | 297.8 | 23.1 | 15.2 | 15.2 | 15.2 | 80 |
| Лубоед сосновый большой | 39 | 25.1 | 25.1 | 0 | 23.1 | 259 | 121.3 | 48.6 |
| Златка лиственничная | 0 | 0 | 45 | 45 | 45 | 45 | 45 | 96.3 |
| Скрипун осиновый | 0 | 0 | 0 | 0 | 0 | 35 | 35 | 12.8 |
| Лубоед пальцеходный | 0 | 0 | 0 | 15.2 | 0 | 0 | 0 | 0 |
| Усач черный еловый большой | 0 | 3287.4 | 3279.6 | 3241.2 | 3241.2 | 3241.2 | 3203.2 | 3203.2 |
По данным табл. 4, согласно приведенной выше методике расчетов, вычислялась корреляционная матрица временной сопряженности очагов разных видов ксилофагов на территории этого лесничества (табл. 5).
Таблица 5. Корреляционная матрица временной сопряженности очагов массового размножения ксилофагов в Гремячинском лесничестве
Table 5. Correlation matrix of the temporary conjugation of xylophage outbreaks in the Gremyachinsky forest area
| № вида | Вид насекомых-ксилофагов* | ||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
| 1 | 1.000 | -0.134 | 0.655** | 0.737* | -0.258 | -0.814* | -0.403 | -0.218 | -0.642 |
| 2 | 1.000 | -0.088 | -0.220 | 0.976* | -0.005 | 0.804* | -0.331 | 0.085 | |
| 3 | 1.000 | 0.483 | -0.136 | -0.533 | -0.264 | -0.143 | -1.000* | ||
| 4 | 1.000 | -0.405 | -0.590 | -0.559 | -0.308 | -0.464 | |||
| 5 | 1.000 | 0.123 | 0.881* | -0.321 | 0.129 | ||||
| 6 | 1.000 | 0.313 | 0.064 | 0.516 | |||||
| 7 | 1.000 | -0.264 | 0.249 | ||||||
| 8 | 1.000 | 0.142 | |||||||
| 9 | 1.000 | ||||||||
Примечание. * – виды: 1 – златка бронзовая, 2 – короед большой лиственничный (продолговатый), 3 – короед шестизубый, 4 – лубоед малый сосновый, 5 – лубоед большой еловый, 6 – златка лиственничная, 7 – скрипун большой осиновый, 8 – лубоед пальцеходный, 9 – усач черный еловый большой. ** – корреляции значимы на уровне р = 0.90.
Как известно, для видов i и j коэффициент корреляции r (i, j) = r (j, i), поэтому корреляционная матрица симметрична относительно главной диагонали, и данные ниже главной диагонали можно не приводить. Как показано в табл. 5, возможна положительная сопряженность встречаемости очагов двух видов в одном лесничестве (коэффициент корреляции r > 0 и значим на уровне р = 0.90) и отрицательная сопряженность очагов двух видов в одном лесничестве (r < 0 и значим на уровне р = 0.90). В первом случае при наличии в лесничестве очага одного вида большей частью встречается и очаг положительно сопряженного вида. Так, положительно сопряжены очаги массового размножения короеда большого лиственничного (продолговатого) (вид 2) и лубоеда соснового большого (вид 5). Для этих видов коэффициент корреляции r (2, 5) = +0.976. Положительно сопряжены очаги массового размножения короеда большого лиственничного (продолговатого) (вид 2) и скрипуна большого осинового (вид 7). Для этих видов коэффициент корреляции r (2, 7) = +0.804.
При отрицательной сопряженности в случае наличия очага одного вида очаг другого вида не встречается. Так происходит с очагами короеда шестизубого (вид 3) и усача черного елового большого (вид 9). Для этих видов коэффициент корреляции r (3, 9) = - 1.0.
На основе расчетов корреляционных матриц временной сопряженности очагов массового размножения комплекса насекомых-ксилофагов в различных лесничествах Красноярского края выявлена положительная сопряженность в динамике ряда видов ксилофагов (табл. 6).
Таблица 6. Положительная сопряженность очагов отдельных видов ксилофагов
Table 6. Positive temporary conjugation of foci of certain types of xylophages
| Вид насекомых-ксилофагов | Число сопряженных видов | Наиболее частый взаимосвязанный вид |
| Усач черный сосновый | 10 | Заболонник березовый |
| Лубоед большой сосновый | 8 | Лубоед малый сосновый |
| Короед шестизубый | 8 | Усач черный сосновый |
| Усач черный еловый большой | 8 | Усач черный сосновый |
| Заболонник березовый | 7 | Усач черный еловый большой |
| Златка лиственничная | 6 | Златка бронзовая |
| Полиграф уссурийский (белопихтовый) | 6 | Усач черный еловый большой |
| Златка бронзовая | 5 | Златка лиственничная |
| Короед большой лиственничный (продолговатый) | 5 | Усач черный сосновый |
| Короед-типограф | 4 | |
| Лубоед малый сосновый | 4 | Усач черный сосновый |
| Лубоед большой еловый | 3 | |
| Лубоед пальцеходный | 3 | Лубоед большой сосновый |
| Скрипун большой осиновый | 2 | |
| Усач черный еловый малый | 2 |
Усач черный сосновый M. galloprovincialis (см. табл. 6) является видом, с которым сопряжено наибольшее число очагов других видов-ксилофагов данного комплекса. Сопряженные очаги этот вид образует с 10 другими видами, из них наиболее часто взаимосвязаны очаги усача черного соснового и заболонника березового. Усач черный сосновый повреждает все хвойные, но наиболее сильно – сосну обыкновенную. При прохождении дополнительного питания в кронах деревьев жуки обгладывают кору тонких веток, чем при массовом размножении существенно ослабляют деревья. Помимо хвойных деревьев жуки могут повреждать ветви и лиственных пород, таких как береза и осина. Заболонник березовый широко распространен и повреждает, как правило, старые и ослабленные березы. Так как эти виды не конкурируют за кормовой ресурс, то сопряженность их очагов, по П. Морану (Moran, 1953), может быть связана с общностью требований к климатическим условиям.
Достаточно большая сопряженность с очагами других видов характерна для лубоеда большого соснового, короеда шестизубого и усача черного елового большого. Число взаимосвязанных видов стволовых насекомых с этими тремя видами равно 8. При этом максимально часто сопряженными видами являются лубоед малый сосновый и усач черный сосновый.
Лубоед малый сосновый, как и лубоед большой сосновый, встречается в ареале сосны обыкновенной повсеместно. Жуки питаются на ослабленных соснах. Район поселения – вершины, крупные ветви, центральная часть стволов деревьев различного возраста. Кроме этого, лубоед «стрижкой» ослабляет здоровые, еще не заселенные им сосны, тем самым подготавливая базу для дальнейшего расселения.
Жуки лубоеда большого соснового нападают на ослабленные сосны, часто образуют очаги на гарях, в очагах корневой губки, в условиях техногенной нагрузки. Эти виды относятся к весеннему фенологическому комплексу стволовых вредителей.
Черные хвойные усачи развиваются на всех хвойных породах. Жуки дополнительно питаются в кронах деревьев хвоей и лубом на побегах и ветвях живых деревьев. Большой черный еловый усач в лесах Сибири и Дальнего Востока размножается в огромных количествах в очагах сибирского шелкопряда, на гарях, в местах крупных лесозаготовок. Все хвойные усачи заселяют и стоящие, и поваленные деревья (Лесная энтомология, 2010). Эти виды могут выступать в качестве конкурентов, однако все же формируют сопряженные (комплексные) очаги. Это связано с тем, что лубоед большой сосновый, лубоед малый сосновый, черные хвойные усачи способны к дополнительному питанию на имагинальной фазе. Дополнительное питание жуки проводят на побегах и ветвях жизнеспособных деревьев, ослабляя их и расширяя свою кормовую базу. Кроме этого, совместное освоение древесных растений различными видами ксилофагов позволяет преодолеть сопротивление физиологических систем дерева (Исаев, Гирс, 1975).
Пространственную сопряженность очагов массового размножения отдельного вида можно охарактеризовать числом лесничеств, в которых наблюдаются вспышки его массового размножения. Так, очаги массового размножения усача черного елового большого встречались в 36 лесничествах, усача черного соснового – в 20, лубоеда малого соснового и полиграфа уссурийского – в 13 лесничествах. К числу часто встречающихся на территории края видов следует отнести лубоеда большого соснового, короеда-типографа (в 10 лесничествах). Скрипун большой осиновый с очагом площадью 35 га зафиксирован на территории одного лесничества (см. табл. 1). Причинами пространственной сопряженности могут выступать синхронизация популяционной динамики и состояния кормовых растений, сходство реакции популяций на изменение погоды в различных местообитаниях.
Риск вспышек массового размножения отдельного вида ксилофагов на территории края можно характеризовать, используя следующие показатели: доля лесничеств, в которых встречаются очаги этого вида, показатели сопряженности очагов этого вида в разных лесничествах, площадь очагов массового размножения данного вида по отношению к общей площади лесных насаждений. Чем больше эти значения, тем больше риск вспышек данного вида ксилофагов.
В качестве иллюстрации приведем оценку риска возникновения вспышки массового размножения усача черного елового большого в лесничествах на территории Красноярского края на протяжении 2007–2014 гг.
Имея данные по площади очагов Monochamus urussovi в пространстве лесничеств Красноярского края и времени (2008–2014 гг.) (табл. 7), рассчитаем корреляционную матрицу пространственной сопряженности очагов усача черного елового большого в различных лесничествах на территории Красноярского края. В табл. 8 приведен фрагмент этой корреляционной матрицы (привести целиком корреляционную матрицу затруднительно ввиду ее больших размеров).
Таблица 7. Площади очагов Monochamus urussovi Fisch. в лесничествах на территории Красноярского края на протяжении 2008–2014 гг.
Table 7. Areas of outbreak foci for Monochamus urussovi Fisch. on Krasnoyarsk Region territory for 2008–2014
| Лесничество | Площадь очагов Monochamus urussovi Fisch.(га) по годам | ||||||
| 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | |
| Ачинское | 13.7 | 13 | 13 | 13 | 13 | 13 | 13 |
| Балахтинское | 41.5 | 35.3 | 33.2 | 0 | 0 | 0 | 0 |
| Боготольское | 373.8 | 350.8 | 350.8 | 350.8 | 350.8 | 25 | 25 |
| В.-Манское | 659 | 659 | 609 | 609 | 609 | 609 | 609 |
| Гремучинское | 3287.4 | 3279.6 | 3241.2 | 3241.2 | 3241.2 | 3203.2 | 3203.2 |
| Мининское | 8.1 | 8.1 | 8.1 | 0 | 0 | 0 | 0 |
| Дзержинское | 0 | 0 | 0 | 0 | 2 | 2 | 2 |
| Д.-Мостовское | 95.3 | 95.3 | 205.3 | 110 | 110 | 110 | 110 |
| Енисейское | 10081 | 10144 | 1711 | 2227 | 2227 | 831 | 831 |
| Ермаковское | 3280.4 | 3280.4 | 3280.4 | 3280.4 | 3280.4 | 3140 | 3140 |
| Ирбейское | 35508 | 35508 | 35508 | 35508 | 27250 | 26520 | 0 |
| Казачинское | 7750 | 150 | 150 | 0 | 0 | 0 | 0 |
| Каратузское | 4065 | 4065 | 3434 | 3434 | 3567 | 1732 | 1732 |
| Кодинское | 2391 | 2391 | 2391 | 2391 | 2391 | 2293 | 2578.9 |
| Козульское | 3.9 | 3.9 | 3.9 | 3.9 | 3.9 | 0 | 0 |
| Красноярское | 89.5 | 84 | 84 | 0 | 0 | 0 | 0 |
| Кизирское | 798 | 2961.1 | 2669.3 | 2650.9 | 2496.2 | 2496.2 | 2048.2 |
| Маганское | 83 | 83 | 82.5 | 82.5 | 82.5 | 82.5 | 82.5 |
| Манзенское | 67.1 | 48.5 | 22 | 22 | 22 | 22 | 22 |
| Манское | 574.2 | 571.4 | 598.1 | 583.2 | 583.2 | 583.2 | 583.2 |
| Мотыгинское | 42143.6 | 40644.6 | 40361.6 | 40361.6 | 40361.6 | 35780.6 | 15342 |
| Невонское | 104.9 | 104.9 | 104.9 | 104.9 | 104.9 | 137.9 | 137.9 |
| Пировское | 5570.2 | 5532.7 | 5532.7 | 5532.7 | 5532.7 | 5343.2 | 3442 |
| С.-Шушенское | 956 | 956.1 | 956.1 | 0 | 0 | 956.1 | 956.1 |
| Саянское | 1.2 | 0 | 0 | 0 | 0 | 0 | 0 |
| С.-Енисейское | 330.5 | 330.5 | 309.2 | 25.9 | 25.9 | 25.9 | 5.9 |
| Таёжинское | 2252.7 | 2135.2 | 1953.9 | 1310.4 | 1289.7 | 1281.4 | 1281.4 |
| Терянское | 759 | 759 | 759 | 759 | 759 | 654 | 654 |
| Т.-Чунское | 0.6 | 0.6 | 0.6 | 0.6 | 0.6 | 0.6 | 0.6 |
| Тюхтетское | 173 | 173 | 173 | 173 | 173 | 172.5 | 172.5 |
| Ужурское | 35 | 35 | 35 | 35 | 35 | 35 | 35 |
| Усольское | 1666 | 1666 | 39 | 39 | 39 | 39 | 39 |
| Уярское | 346.5 | 346.5 | 0 | 0 | 0 | 0 | 0 |
| Хребтовское | 768 | 768 | 768 | 717 | 717 | 717 | 717 |
| Чунское | 1552 | 0 | 0 | 956.1 | 956.1 | 17 | 17 |
| Усинское | 0 | 0 | 0 | 0 | 1499 | 1499 | 1499 |
Таблица 8. Фрагмент корреляционной матрицы пространственной сопряженности очагов усача черного елового большого в различных лесничествах на территории Красноярского края
Table 8. A fragment of the correlation matrix of spatial contiguity of the foci of the Monochamus urussovi Fisch in various forestries in the Krasnoyarsk Territory
| Номер лесничества | Номер лесничества* | |||||||
| 1 | 5 | 7 | 9 | 16 | 17 | 20 | 32 | |
| 1 | 1.00 | 0.60** | -0.35 | 0.64* | 0.51 | -0.92* | -0.42 | 0.65* |
| 5 | 1.00 | -0.76* | 0.91* | 0.78* | -0.30 | -0.51 | 0.85* | |
| 7 | 1.00 | -0.60* | -0.75* | 0.06 | 0.09 | -0.55 | ||
| 9 | 1.00 | 0.75* | -0.37 | -0.75* | 0.99* | |||
| 16 | 1.00 | -0.25 | -0.14 | 0.74* | ||||
| 17 | 1.00 | 0.31 | -0.40 | |||||
| 20 | 1.00 | -0.76* | ||||||
| 32 | 1.00 | |||||||
Примечание. * – лесничества: 1 – Ачинское, 5 – Гремучинское, 7 – Дзержинское, 9 – Енисейское, 16 – Красноярское, 17 – Кизирское, 20 – Манское, 32 – Усольское. ** – коэффициент корреляции значим при р = 0.90.
Анализ табл. 8 позволяет выделить территории с синхронными в пространстве очагами массового размножения усача черного елового большого. Так, например, динамика развития очагов усача черного елового большого совпадает в Гремучинском, Енисейском и Усольском лесничествах (коэффициенты корреляции более 0.78), тогда как временная динамика развития очага этого вида антикоррелирует в Ачинском и Кизирском лесничествах. Таким образом, доля лесничеств, в которых отмечаются очаги данного вида, равна 0.581. По всему комплексу рассмотренных ксилофагов информация по площади очагов и доли лесничеств, в которых встречаются очаги, приведена в табл. 9.
Таблица 9. Встречаемость очагов массового размножения отдельных видов ксилофагов в лесничествах на территории Красноярского края на протяжении 2007–2014 гг.
Table 9. Occurrence of outbreaks foci of certain xylophages species in forest areas of Krasnoyarsk Region during 2007–2014
| Вид насекомых-ксилофагов | Максимальная годичная площадь очагов, га | Число лесничеств с очагами данного вида | Доля лесничеств с очагами данного вида | Отношение площадей очагов к площадям лесных насаждений |
| Усач черный еловый большой | 153085 | 36 | 0.581 | 0.00156 |
| Полиграф уссурийский (белопихтовый) | 8437.6 | 13 | 0.210 | 8.61*10-5 |
| Усач черный сосновый | 7050.3 | 20 | 0.323 | 7.19488*10-5 |
| Короед шестизубый | 3250.7 | 2 | 0.032 | 3.31736*10-5 |
| Лубоед малый сосновый | 2031.5 | 10 | 0.161 | 2.07316*10-5 |
| Короед-типограф | 1223.8 | 10 | 0.161 | 1.2489*10-5 |
| Лубоед малый сосновый | 928 | 13 | 0.210 | 9.47031*10-6 |
| Златка лиственничная | 809.1 | 6 | 0.097 | 8.25692*10-6 |
| Короед большой лиственничный (продолговатый) | 314 | 9 | 0.145 | 3.20439*10-6 |
| Заболонник березовый | 218 | 7 | 0.113 | 2.22471*10-6 |
| Златка бронзовая | 184 | 4 | 0.065 | 1.87773*10-6 |
| Усач черный еловый малый | 87 | 6 | 0.097 | 8.87841*10-7 |
| Лубоед пальцеходный | 55 | 5 | 0.081 | 5.61279*10-7 |
| Скрипун большой осиновый | 35 | 1 | 0.016 | 3.57177*10-7 |
| Лубоед большой еловый | 31 | 2 | 0.032 | 3.16357*10-7 |
Риски развития вспышек массового размножения различных видов ксилофагов на территориях разных лесничеств можно представить графически на рис. 2, где по оси абсцисс на этом графике – доля q лесничеств, в которых наблюдаются вспышки данного вида, а по оси ординат – логарифм доли S0 площади очагов данного вида от общей территории лесов края. Точка на плоскости характеризует отдельный вид. В качестве примера отмечены точки, характеризующие усача черного елового большого (точка 1), усача черного соснового (точка 2), короеда шестизубого (точка 3).
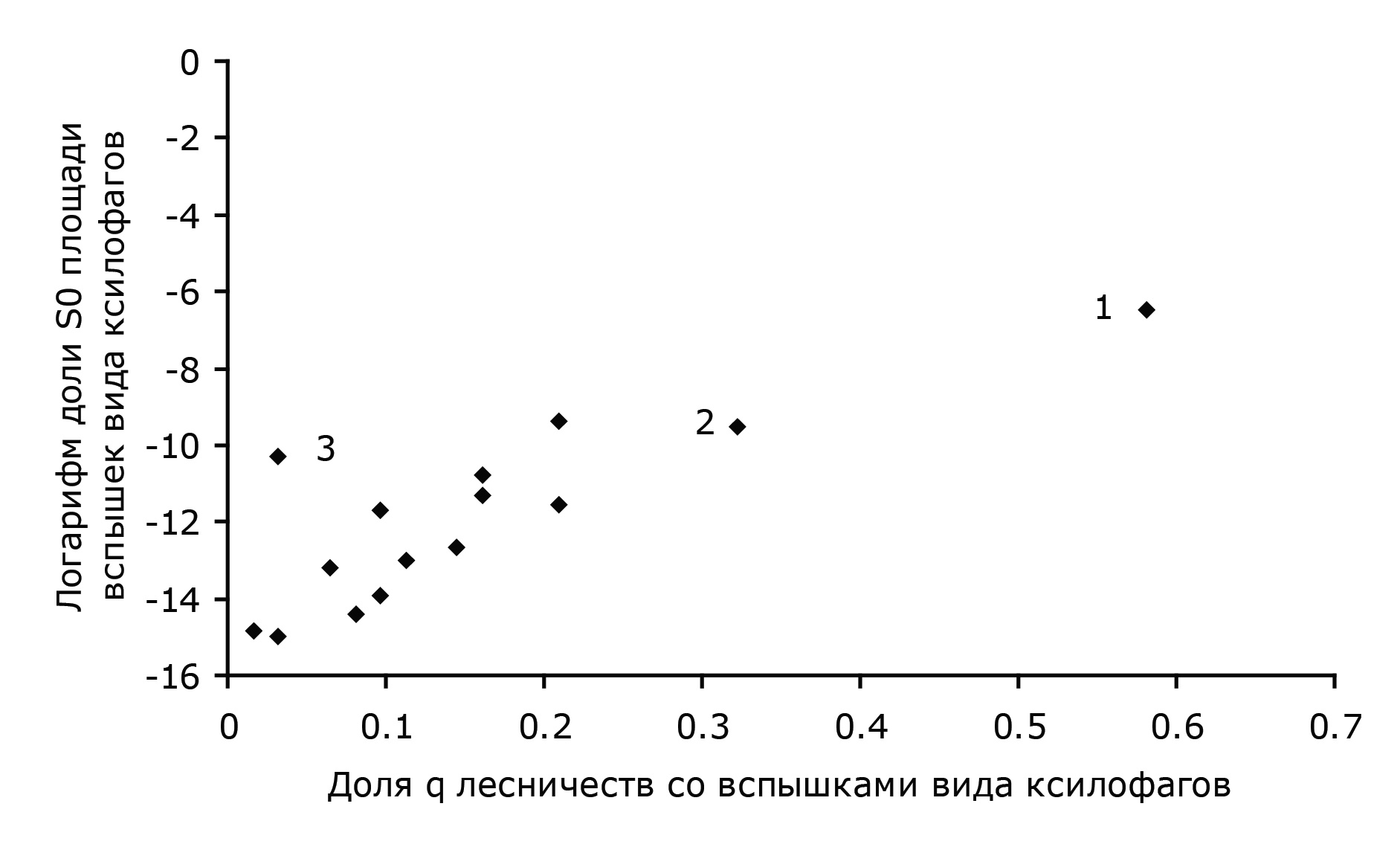
Рис. 2. Риски вспышек массового размножения различных видов ксилофагов
Fig. 2. The risks of outbreaks of various xylophages species
Точки в нижнем левом углу плоскости {q, ln S0} характеризуют виды с низким уровнем воздействия на лес (мала доля лесничеств, в которых встречаются очаги этих видов ксилофагов, и малы площади очагов). К этой группе принадлежат большинство видов из изученного энтомокомплекса: лубоед большой еловый, скрипун большой осиновый, лубоед пальцеходный, усач черный еловый малый, златка бронзовая и др. Точки в верхнем правом углу плоскости {q, ln S0} характеризуют виды ксилофагов с сильным воздействием на лес (очаги массового размножения этих видов встречаются в большинстве лесничеств на территории Красноярского края и площади очагов велики). К видам из этой группы принадлежат усач черный еловый большой, усач черный сосновый, лубоед малый сосновый, полиграф уссурийский (белопихтовый). В верхнем левом углу плоскости {q, ln S0} на рис. 2 должны находиться точки, характеризующие локально воздействующие виды (встречающиеся в малом числе лесничеств, но при этом площади очагов в этих лесничествах велики). В нижнем правом углу плоскости {q, ln S0} должны находиться точки, характеризующие диффузно воздействующие виды с очагами в большом числе лесничеств, но с малой площадью. Однако, как это видно из рис. 2, локально и диффузно воздействующих видов ксилофагов на территории лесничеств Красноярского края в течение времени исследований не встречалось.
Обсуждение
Проблема возникновения очагов и повторяемость вспышек массовых размножений дендрофильных насекомых в течение многих десятилетий занимает ведущее место в экологических исследованиях во всем мире. В середине двадцатых годов ХХ века экологами были выдвинуты теоретические представления о периодичности массовых размножений, их взаимодействии с циклами солнечной активности, климата и естественных врагов (энтомофагов). Главным же фактором динамики численности стволовых насекомых является количество и качество корма. Погода и другие экологические факторы оказывают опосредованное влияние на динамику популяций через состояние кормовых растений. В отечественной и зарубежной экологической литературе давно поднимается вопрос о связи популяционных циклов насекомых с климатическими факторами. Известно (Берриман, 1990), что циклы разных популяций одного вида, разделенных большими расстояниями, могут проходить синхронно друг с другом. Механизмы, обеспечивающие синхронное увеличение численности популяций насекомых на обширной территории, детально еще не известны. Предполагается, что причиной синхронизации вспышек массового размножения насекомых может быть некоторый внешний фактор, воздействие которого приводит к одновременному развитию локальных вспышек. В частности, такими синхронизирующими факторами могут быть изменение активности Солнца (Чижевский, 1973), летние засухи на обширной территории (Кондаков, 1974, 2002). Так как ритм солнечной активности определяет динамику воздействия солнечной радиации одновременно на всей планете, а вспышки численности различных видов насекомых не синхронизированы во времени и пространстве, то предполагается, что синхронизирующий фактор и есть сочетание ритма солнечной активности и локальных планетарных ритмов (Moran, 1953). По мнению разных авторов, причиной сопряженности популяционной динамики одного вида в разных местообитаниях может быть эффект П. Морана, связанный с однородностью климатических условий на значительной территории и сходством реакции популяций на изменение погоды в различных местообитаниях (Максимов, 1989; Bjornstad, Bascompte, 2001; Пальникова и др., 2014). Показано (Liebhold, Kamata, 2000; Liebhold et al., 2004), что степень сопряженности популяционной динамики одного вида в разных местообитаниях монотонно уменьшается с увеличением расстояния между этими местообитаниями. Если же уровень сопряженности динамики популяций не уменьшается при увеличении расстояния между местообитаниями, а расстояние между ними существенно превосходит радиус индивидуального перемещения особей изучаемого вида, то следует говорить о его глобальной пространственной когерентности, связанной с реакцией популяций на воздействие мощного модифицирующего фактора.
Динамика распространения очагов, синхронность их формирования в пространстве и степень синхронности формирования очагов разных видов в одном местообитании имеют практическое значение. Однако для этого требуются материалы по инвентаризации очагов популяций дендрофильных видов на территориях лесничеств, выполненные специалистами. Использование корреляционных матриц пространственной сопряженности для отдельного вида позволяет при обнаружении очагов массового размножения этого вида ксилофагов в одном лесничестве дать оценку рисков возникновения вспышек этого вида в других лесничествах. Использование корреляционной матрицы временной сопряженности динамики нескольких видов в отдельном лесничестве позволяет при обнаружении очага массового размножения одного вида ксилофагов в этом лесничестве дать оценки рисков развития вспышек других видов ксилофагов в этом лесничестве.
Заключение
1. Наиболее опасными видами насекомых-ксилофагов по риску воздействия на лесные насаждения на территории Красноярского края являются: Monochamus urussovi Fisch., Polygraphus proximus Blandf., Monochamus galloprovincialis Ol., Tomicus piniperda L.
2. Для оценки риска развития сопряженных очагов ряда видов в одном лесничестве можно использовать корреляционные матрицы временной сопряженности популяций ксилофагов. Так, например, для Гремучинского лесничества высок риск одновременного развития (временной сопряженности) нескольких видов ксилофагов: короеда большого лиственничного (продолговатого) и лубоеда большого соснового; короеда большого лиственничного (продолговатого) и скрипуна большого осинового; лубоеда большого соснового и скрипуна большого осинового; златки бронзовой и лубоеда малого соснового; златки бронзовой и короеда шестизубого.
3. Для оценки риска пространственной сопряженности массового размножения конкретного вида ксилофага в разных лесничествах можно использовать корреляционные матрицы пространственной сопряженности очагов данного вида. Наибольшая степень пространственной сопряженности очагов характерна для усача черного елового большого, усача черного соснового, полиграфа уссурийского (белопихтового). Это указывает на пространственную распространенность данных видов, следует ожидать одновременного появления очагов размножения этих видов на территории различных лесничеств.
Библиография
Берриман А. Защита леса от насекомых-вредителей . М.: ВО Агропромиздат, 1990. 288 с.
Государственный доклад «О состоянии и охране окружающей среды в Красноярском крае в 2017 году» . Красноярск, 2018. С. 171–172. URL: http://mpr.krskstate.ru/dat/File/3/doklad%202017. pdf.
Исаев А. С., Гирс Г. И. Взаимодействие дерева и насекомых-ксилофагов . Новосибирск: Наука, 1975. 348 с.
Лесная энтомология: учебник для студ. высш. учеб. заведений / Е. Г. Морозова, А. В. Селиховкин, С. С. Ижевский и др.; под ред. Е. Г. Мозолевской. М.: Издательский центр «Академия», 2010. 416 с.
Кондаков Ю. П. Закономерности массовых размножений сибирского шелкопряда . Новосибирск: Наука, 1974. С. 206–265.
Кондаков Ю. П. Массовые размножения сибирского шелкопряда в лесах Красноярского края // Энтомологические исследования в Сибири. Красноярск: КФ РЭО, 2002. Вып. 2. С. 25–74.
Максимов А. А Природные циклы: Причины повторяемости экологических процессов . Л.: Наука, 1989. 236 с.
Пальникова Е. Н., Суховольский В. Г., Тарасова О. В. Пространственно-временная когерентность популяционной динамики лесных насекомых-филлофагов // Евразиатский энтомологический журнал. 2014. № 13 (3). С. 228–236.
Чижевский А. Л. Земное эхо солнечных бурь . М.: Мысль, 1973. 349 с.
Bjornstad O., Bascompte J. Synchrony and second order spatial correlation in host–parasitoid system // Journal of Animal Ecology. 2001. Vol. 70. P. 924–933.
Moran P. A. P. The statistical analysis of the Canadian lynx cycle. II. Synchronization and meteorology // Australian Journal of Zoology. 1953. Vol. 1. P. 291–298. DOI: 10.1071/zo9530291.
Liebhold A., Kamata N. Are population cycles and spatial synchrony universal characteristics of forest insect population? // Population Ecology. 2000. Vol. 42. P. 205–209.
Liebhold A., Sork V., Peltonen M., Koenig W., Bjørnstad O. N., Westfall R., Elkinton J., Knops J. M. H. Within-population spatial synchrony in mast seeding of North American oaks // Oikos. 2004. Vol. 104. P. 156–164.

 © 2011 - 2026
© 2011 - 2026