



Исследования по термобиологии рептилий начались за рубежом в первой половине XIX в. Змеи, ящерицы, черепахи и крокодилы обильно населяли (и сейчас, конечно, тоже населяют) тропические страны (в XIX в. это чаще всего были колонии), в которых эти исследования изначально и проводились учеными и просто интересующимися любителями (и те и другие в основном – представители митрополий). Кроме того, среди рептилий тропиков многие животные представляют реальную опасность для человека (множество ядовитых змей, опасных крупных удавов и питонов, крокодилы и т. п.). Поэтому, особенно раньше, они играли серьезную роль в жизни людей и на них было обращено большое внимание. Это внимание, активированное, наверное, изначально в значительной степени страхом, в дальнейшем перерастало в любопытство, а затем и в научный интерес.
ИЗУЧЕНИЕ ТЕРМОБИОЛОГИИ РЕПТИЛИЙ В ЗАРУБЕЖНЫХ СТРАНАХ
Первая известная нам работа, касающаяся темы теплового баланса рептилий, была опубликована еще в первой половине XIX в. (Valensiennes, 1841). В ней говорилось о том, что насиживающие самки питонов заметно повышают температуру своего тела особенно в месте, где они покрывают кладку. Следующая публикация на эту же тему вышла уже через год (Lamarre-Pacquot, 1842). Но самое раннее устное сообщение этого автора Французской академии наук о том, что в Бенгалии насиживающие самки питонов повышают температуру тела на несколько градусов, состоялось еще в 1832 г.
Первое конкретное упоминание о температуре тела рептилий, которое нам известно, относится к середине XIX в. (Agassiz, Gould, 1856). В конце XIX в. для измерения температуры тела некоторые ученые стали применять термопары (Sutherlanad, 1896), что расширило возможности полевых и лабораторных исследований. Наблюдения за поведением, связанные с использованием тепла и с тепловой выносливостью рептилий, публиковались с конца XIX в. (Werner, 1891; Richet, 1889; Soetbeer, 1898).
Накопление первичного материала в том же направлении шло с самого начала XX в. (Gadow, 1901; Grinnell, 1908; Camp, 1916; Weese, 1917, 1919) и продолжалось вплоть до конца 30-х гг. (Buxton, 1923, 1924; Hall, 1922; Reese, 1923; Hesse, 1924; Parker, 1925; Kanitz, 1925; Baldwin, 1925a, b; Kammerer, 1926; Pearse, Hall, 1928; Krüger, 1924, 1929; Krüger, Kern, 1924; Weigmann, 1929; Franz, 1930; Pearse, 1931; Benedict, 1932; Blum, Spealman, 1933; Mosauer, Lazier, 1933; Mcilhenny, 1935; Parker, 1935; Mosauer, 1930, 1936; Lufti, 1936; Warden et al., 1936; Hesse et al., 1937; Atsatt, 1939; Bogert, 1939; Cowles, 1939; Klauber, 1939). При этом количество публикаций в 30-е гг. начало существенно возрастать. Если с середины XIX по конец 20-х гг. XX в. нам известна всего 31 публикация, то только за 30-е гг. – уже 39 (рисунок). В конце 30-х гг. в списке авторов появляются ученые, которые впоследствии станут классиками в области термобиологии, – это Р. Б. Коул (R. B. Cowles) и К. М. Богерт (C. M. Bogert).
Публикации, в той или иной мере касающиеся термобиологии рептилий, были связаны с описанием экологии этих животных, условий их обитания, поведенческих реакций, направленных на избегание температурой смерти от перегрева, на выбор наиболее адекватных и безопасных микробиотопов, времени активности и т. п. Но большая часть материалов касалась темы температурной выносливости рептилий. Наверное, это было связано с тем, что многие первые наблюдения над биологией пресмыкающихся проводились в местах, где их было больше всего и где они были визуально доступнее – в пустынях. Исследователей весьма впечатляло, что они встречали прежде всего ящериц и иногда змей а) при очень высоких температурах среды и/или б) в местах, где такие температуры периодически присутствовали. При этом пункт а часто ошибочно отождествляли с пунктом б. Тем не менее это первое, что бросалось в глаза, вызывало порой недоумение и, следовательно, научный интерес, а опыты по изучению температурной выносливости изначально могли быть поставлены даже на самом примитивном уровне, не требующем серьезной экипировки и научной подготовки (например: за сколько минут погибает змея, помещенная на почву с температурой 40°, 42° и т. д.).
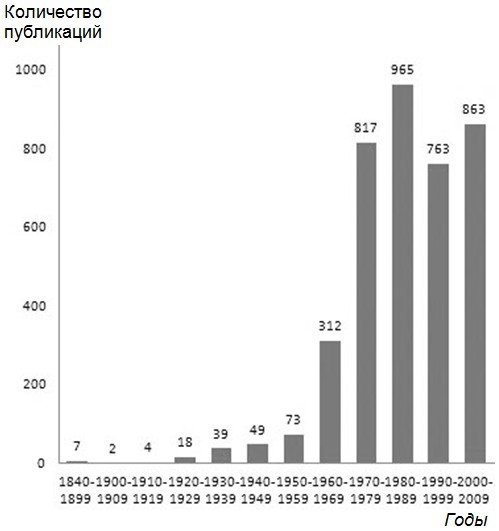
Количество известных нам зарубежных научных публикаций, касающихся термобиологии рептилий и ближайших смежных областей, по годам
Правда, уже в этот начальный период изучения термобиологии рептилий появлялись статьи, представляющие собой попытки осмысления значения температуры в жизни животных, соотношения экто- и эндотермии (Pearse, Hall, 1928; Benedict, 1932; Warden et al., 1936 и др.), но они были единичными.
Таким образом, с середины XIX в. по конец 30-х гг. XX в. в зарубежных исследованиях термобиологии рептилий происходил период первичного накопления научного материала по наиболее ярким, обращающим на себя внимание особенностям биологии пресмыкающихся, связанным с температурой.
Публикации 1940-х гг. (нам известно их 49, см. рисунок) продемонстрировали желание исследователей глубже проникнуть в суть биологии рептилий. Кроме уже ставших обычными тем экологии (Lueth, 1941; Lowe, 1942; Cowles, 1944; Bogert, 1949a, b и др.) и термальной выносливости (Cowles, 1940, 1942, 1945; Cole, 1943; Cowles, Burleson, 1945; Colbert et al., 1946; Woodbury, Hardy, 1948 и др.), встречались работы, посвященные другим важным аспектам: зимовкам рептилий и термальной выносливости при низких температурах (Cowles, 1941; Bailey, 1948, 1949), влиянию температуры на развитие морфологических признаков у змей (Klauber, 1941; Fox, 1948), общим проблемам терморегуляции (Herter, 1941; Cowles, 1942; Bogert, 1949а, b).
Но особое значение, наверное, имеет статья, посвященная общим вопросам термобиологии рептилий (Cowles, Bogert, 1944). Большинством герпетологов она считается первой серьезной работой по термобиологии рептилий, с которой можно исчислять начало развития этого направления науки. В ней авторы приводят результаты достаточно подробных исследований термобиологии двенадцати видов дневных и семи видов ночных рептилий (ящериц и змей) юго-западной части США. Они четко указали (правда, к сожалению, лишь вскользь) на ряд важнейших положений, которые их научными последователями, к сожалению, не были замечены. Так, они выделяли у ящериц состояние «нормальной активности» (normal activity), хотя на этом важном методическом аспекте, к сожалению, не сконцентрировали внимания. По сути, это состояние полностью соответствует тому, которое оказалось впервые описано как «полная активность[1]» годом ранее С. С. Либерман и Н. В. Покровской (1943), хотя с последней статьей Р. Б. Коул и К. М. Богерт явно еще не были знакомы. Кроме того, в своей работе они ясно продемонстрировали (хотя также, увы, не акцентировали на этом внимание), что данные по температурам активности и тем более по предпочитаемым температурам должны выделяться только из выборок температур тела при нормальной активности. Это чрезвычайно важно, поскольку если не учитывать указанные выше обстоятельства, то исследователи получат некорректные, ошибочные первичные данные, а это, в свою очередь, повлечет за собой ошибки в анализе материала и в выводах, касающихся даже самых общих (физиологических, эволюционных) проблем (Черлин, 2013, 2014а; Cherlin, 2015a).
В результате приведенные Р. Б. Коулом и К. М. Богертом в 1944 г. термобиологические показатели активности и термального предпочтения рептилий оказались методически намного более корректно определенными (как по смыслу, так и в числовом выражении), чем у большинства последовавших за ними исследователей. Другими словами, публикация Р. Б. Коула и К. М. Богерта (Cowles, Bogert, 1944) имеет намного большее научное, но в первую очередь – методическое, значение, чем это сейчас принято считать. Однако при этом она не является первой важной статьей по термобиологии рептилий. Это первенство по праву должно принадлежать работам И. Д. Стрельникова (1934 и др.), а также С. С. Либерман и Н. В. Покровской (1943), но об этом немного позже.
Публикации 50-х гг. (нам известно их 73, см. рисунок) имели разную научную направленность: были исследования по экологии и терморегуляционному поведению рептилий (Carpenter, 1952; Norris, 1953; Pearson, 1954; Fitch, 1954; Cowles, 1956; Inger, 1959 и др.), по температуре их тела в природе и ее динамике (Bogert, 1953; Carpenter, 1956; Fitch, 1956), по термофизиологии и термальной выносливости (Lowe, Vance, 1955; Parker, 1956; Reichling, 1957; Cowles, 1958b; Evans, Cowles, 1959; Musacchia, 1959), по терморегуляции (Saint-Girons, Saint-Girons, 1956; Reichling, 1957; Cowles, 1958а; Bogert, 1959).
Таким образом, для 40-х и 50-х гг. XX в. в зарубежных странах характерно постепенное, медленное возрастание интереса и количества публикаций по термобиологии рептилий, усиление инструментальной составляющей в исследованиях, особенно в 50-е гг. В этот период также начали формироваться пока еще самые общие представления о связи физиологии и экологии рептилий с температурой. В зарубежной науке это было время заложения фундамента термобиологии рептилий как новой, развивающейся науки, в основном определился круг проблем, набор понятий, терминов, методических приемов работы, появились первые сводки и попытки обобщений.
C начала 1960-х, а особенно в 1970-х гг. количество публикаций по термобиологии рептилий стало увеличиваться лавинообразно (нам известно за 60-е годы 312, а за 70-е – 817 публикаций в зарубежной научной прессе, см. рисунок). Для этого времени характерно проявление максимального интереса к проблемам термофизиологии (метаболизм, энергетический обмен, иммунология, солевой и водный балансы, переваривание пищи, значение циркуляции крови в терморегуляции и т. п.) (Bennett, 1972; Bennett, Ruben, 1979; Bennett et al., 2000; Bligh, Johnson, 1973; Pearson, Brandford, 1976 и др.), а затем по убывающей – к терморегуляции (стали появляться первые серьезные работы, посвященные терморегуляции у рептилий, – Cowles, 1962; Brattstrom, 1965; McGinnis, Moore, 1969 и др., а также принципам организации терморегуляции, которые иногда описываются разными математическими моделями – Hardy, 1965; Heath, 1970; Berk, Heath, 1975; Barber, 1977, 1979; Barber, Crawford, 1977), и терморегуляционному поведению, температурам тела в природе и приспособительной экологии разных видов рептилий, размножению, температурной выносливости и методологии исследований. Именно на этот период пришелся пик научной активности замечательного американского ученого Пауля Лихта (Paul Licht). Он изучал влияние температуры на различные физиологические системы рептилий, ответы эндокринной системы на температуры при регуляции разных функций организма, взаимодействие разных внешних факторов на активность, переваривание корма, репродуктивные функции и т. д. и т. п. Пауль Лихт внес неоценимый вклад в развитие нашего знания и понимания экологической физиологии, физиологической экологии, экологии и других сторон жизнедеятельности рептилий, их адаптаций к среде обитания и т. п.
Таким образом, 1960-е и 1970-е гг. в зарубежных странах стали, в первую очередь, временем углубления знаний о связи внутренних и внешних, физиологических и экологических составляющих термобиологии рептилий.
В 1980–1990-е гг. у зарубежных ученых проявился очень высокий интерес к термобиологии рептилий (за 80-е гг. нам известно 965, за 90-е – 763 публикации в зарубежной научной прессе, см. рисунок). Во-первых, за это время было накоплено большое количество конкретных материалов полевых и лабораторных исследований в области термоэкологии отдельных видов. Скорее всего, это связано с тем, что у большого количества исследователей в западных странах появилась возможность совершать многочисленные экспедиции в тропические регионы. Кроме того, появилось много новых технических средств и электронных приборов, позволяющих более оперативно, просто и эффективно изучать температуру тела животных, использовать дистанционные методы регистрации активности и температуры тела и т. п. Это позволило увеличить скорость накопления первичных данных по термоэкологии и термофизиологии отдельных видов рептилий, быстрее собирать материалы для статей и диссертаций и стало привлекательным для герпетологов.
Во-вторых, были накоплены обширные материалы по термофизиологии, терморегуляции и термобиологическим показателям (Nelson et al., 1984 и др.). Большое и серьезное внимание было уделено метаболизму и энергетике, различным проблемам термофизиологии (влиянию температуры на разные физиологические функции и др.), термоэкологии и терморегуляции (начали активно использоваться электронные технические средства слежения и контроля), влиянию температуры инкубации на пол детенышей и т. п. Здесь также, наверное, большую роль сыграло появление нового, современного технического инструментария. Кроме того, в связи с накоплением большого количества результатов конкретных исследований, в данный период появилась возможность начать реальный анализ этих материалов: сравнения термобиологических показателей рептилий из разных природно-климатических регионов, термофизиологических характеристик, структур активности, термоэкологических особенностей, вариантов организации терморегуляции, обобщений уровня эволюции термобиологической сферы. За эти годы термобиология рептилий в целом сформировалась, укрепила свой научный базис, более или менее определилась с понятийным аппаратом и методическими подходами к исследованиям (к сожалению, лишь «более или менее», о чем подробнее – далее).
В нулевых годах XXI в. происходило продолжение накопления данных полевых исследований термоэкологии отдельных видов рептилий и их связей с условиями среды, изучение и сравнение термобиологических показателей, разных аспектов термофизиологии, а главное – появлялось все большее количество аналитических публикаций, иногда касающихся проблем и моделей терморегуляции, эволюции термобиологической сферы (за это время нам известно 863 публикации в зарубежной научной прессе, см. рисунок).
Однако эти аналитические статьи имели не всегда объективно высокую научную ценность из-за ряда стандартных методических некорректностей, а также недостаточной разработанности методологической базы – стандартизации понятийного аппарата и т. п. Создается впечатление, что основные достижения, соответствующие уровню развития науки, в этой области были получены еще в 1960–1990-х гг., а теперь требовалось их глубокое осмысление и развитие, появление «прорывных» идей и разработок, но с этим дело обстояло не так хорошо, как хотелось бы.
С начала 10-х гг. XXI в. количество публикаций по термобиологии рептилий в зарубежной прессе стало сокращаться. Интерес к этой области понемногу затухал. Скорее всего, это можно объяснить тем, что за все предыдущие годы был накоплен огромный материал по термобиологии отдельных видов рептилий, но адекватного общего взгляда, общей концепции этой области науки, выходящей на общебиологический уровень, так и не было предложено. В этих условиях дальнейшее накопление данных понемногу теряло свой смысл и свое значение.
Таким образом, 1980-е, 1990-е и нулевые годы XXI в. стали в зарубежных странах золотым веком термобиологии рептилий. Основные базы первичных данных по термобиологии рептилий и более или менее обобщающие идеи были накоплены именно в это время.
В исследования по термобиологии рептилий внесли свой огромный вклад многие зарубежные исследователи, изучавшие экологию, физиологию, экологическую физиологию, физиологическую экологию и другие направления термобиологии рептилий в разные годы. Об этих ученых здесь просто нельзя не упомянуть: S. С. Anderson, R. A. Anderson, R. A. Avery, R. M. Andrews, F. H. Pough, R. B. Cowles, C. M. Bogert, P. Licht, G. A. Bartholomew, A. F. Bennett, S. D. Bradshaw, В. H. Brattstrom, T. J. Case, J. L. Cloudsley-Thompson, D. Crews, P. M. C. Davies, W. R. Dawson, B. T. Firth, E. Frankenberg, C. Gans, B. Green, O. E. Greenwald, P. T. Gregory, G. C. Grigg, L. Gvozdik, A. Hailey, J. J. Harlow, J. E. Heath, H. Heatwole, P. E. Hertz, R. B. Huey, V. H. Hutchison, H. B. John-Alder, W. H. Karasov, D. King, C. B. De Witt, J. C. Lee, H. Lillywhite, W. W. Mayhew, S. M. McGinnis, R. Meek, K. A. Nagy, G. Naulleau, K. S. Norris, J. W. Patterson, E. R. Pianka, F. H. Pough, C. L. Prosser, P. J. Regal, H. Saint-Girons, K. Schmidt-Nielsen, F. Seebacher, R. Shine, V. H. Shoemaker, I. F. Spellerberg, R. D. Stevenson, J. R. Templeton, C. R. Tracy, H. Underwood, S. Waldschmidt, G. J. W.Webb, Y. L. Werner, J. M. Whittier и многие другие.
СОСТОЯНИЕ ИЗУЧЕННОСТИ И ПРОБЛЕМЫ В ИССЛЕДОВАНИЯХ ТЕРМОБИОЛОГИИ РЕПТИЛИЙ В ЗАРУБЕЖНЫХ СТРАНАХ
Более чем за 180 лет изучения термобиологии рептилий был накоплен гигантский фактический материал полевых и лабораторных наблюдений, специальных лабораторных опытов. Изучены экология, географическое распространение, биотопическое размещение, термобиология, особенности физиологии и физиологической экологии многих видов пресмыкающихся. На эти темы было опубликовано огромное количество статей и выпущено много книг. Очень многое в биологии рептилий стало более понятным и научно обоснованным. Описать все достижения в этой области просто невозможно. Они очень разнообразны и многоплановы. Но с сожалением вынуждены отметить, что в работах зарубежных исследователей термобиологии рептилий за последнее время, несмотря на обилие публикаций, присутствуют некоторые негативные, на наш взгляд, аспекты.
Одна из серьезных проблем экологических, физиолого-экологических и других исследований заключается в широком применении стандартных, привычных еще с середины прошлого века методов исследований, которые, как показали недавние работы, содержат определенные методологические некорректности, о части которых мы упомянули в предыдущем разделе. Они, к сожалению, искажают результаты и приводят к ошибочным частным и общебиологическим выводам (Черлин, 2010, 2013, 2014а).
Объективность наличия этих проблем отчасти доказывается материалами подробных экологических, натуралистических наблюдений и исследований, отчасти тем, что именно и только при методически корректном варианте работы термобиологические показатели демонстрируют вполне определенные закономерности. Так, диапазоны температур тела при полной активности и температур термостабилизации[2] при методически корректно выполненной работе оказываются внутри вида стабильными, одинаковыми весной, летом и осенью, и даже в биотопах с различными климатическими условиями (прохладными и жаркими, если животные данного вида их населяют). По факту, во все сезоны года рептилии одного вида в периоды времени, когда они температуру тела реально избирают, стараются удерживать ее в одном и том же диапазоне (обзор у Черлин, 2014а).
Но чаще всего в работе герпетологов встречается другой вариант: данные собираются просто при «активности», т. е. когда животное обнаруживается исследователем в зоне активности и двигается: например, когда дневную ящерицу встречают в светлое время дня на поверхности, исследователь не выделяет у нее фазу полной активности и не учитывает то, как она ведет себя по отношению к теплу (старается как можно быстрее нагреться, избирает конкретный диапазон высоких температур тела, не избирает температуру, стремится избежать перегрева или максимально продлить время нахождения при высоких температурах тела и т. п.). Другими словами, в таком варианте работа выполняется методически некорректно. В этом случае диапазон температур тела при такой активности оказывается существенно шире диапазона температур тела при полной активности, а предпочитаемые температуры (средние арифметические из такой выборки) не несут никакой серьезной смысловой нагрузки и проявят изменчивость: будут выше в более теплых климатических и погодных условиях (например, летом, в низинах, в южных регионах) и ниже – в более прохладных (например, весной и осенью, в горах или на севере). При этом разница между одними и теми же показателями, полученными корректно или некорректно (например, температурами термостабилизации и предпочитаемыми температурами тела), может оказаться не в 1–2°, а в прохладные периоды может доходить до 10° (!). Так, для одного и того же вида разные исследователи могут, например, весной на основании своих полевых изысканий при некорректном и корректном вариантах сбора первичной информации показать предпочитаемую температуру тела 22 или 32° (не хочу ссылаться на первоисточник, дабы никого не обижать)! А это уже принципиальные, фатальные различия!
Таким образом, налицо путаница в понятиях, в методике исследований, в показателях активности и термальных предпочтений (оптимальные, предпочитаемые, добровольные, средние), в методах получения первичного материала и вариантах обработки данных и т. п. Эти факты являются вполне объективными показателями необходимости стандартизации понятийного аппарата, учета статусов активности в термобиологии рептилий, коррекции исследовательских методик и т. п. И, несмотря на утверждение многих известных исследователей о том, что «не нужно все усложнять, и так все понятно», ситуация совершенно очевидная: работа по приведению в порядок понятийного аппарата, методологической и методической частей термобиологических исследований первостепенно необходима!
Часть общих проблем термобиологических исследований рептилий связана также с тем, что до сих пор не принята унифицированная, научно обоснованная система термобиологических терминов и понятий. К сожалению, значительная часть герпетологов этого обстоятельства не признает, считая, что все и так понятно без специального обращения на это внимания. Но по факту же одинаковыми терминами разные авторы часто называют термобиологические показатели, полученные по различным методикам. В результате такие одинаковые термины несут разные смысловые нагрузки, т. е. они попросту отражают разные биологические феномены, хотя исследователи пытаются их при этом сравнивать и делают из этого анализа далекоидущие общебиологические выводы. Не нужно быть гением, чтобы понять, что в этом случае все получающиеся таким образом научные построения оказываются далекими от истины.
Несмотря на кажущуюся очевидность ситуации, подавляющая часть герпетологов, занимающихся исследованиями термобиологии рептилий, особенно в поле, не выделяют статусов и форм активности со всеми вытекающими из этого некорректностями и ошибками. Хуже того, если в какой-либо статье приведены таблицы, в которые сведено огромное количество измерений температуры тела разных видов рептилий, но не описана методика измерений, т. е. не указано, в какой фазе активности эти измерения произведены, или хотя бы словесно это подробно не описано. В результате все эти данные теряют свою научную ценность, поскольку невозможно понять, что и как было получено и обработано, были допущены методические ошибки или нет, и даже что вообще означают приведенные данные. Тем не менее получающиеся ошибочные выводы используются исследователями в частных и общебиологических построениях, которые в результате также закономерно получаются некорректными.
К сожалению, этой «болезнью» болеют и зарубежные, и отечественные исследователи, а наши попытки объяснить ситуацию воспринимаются чаще всего, мягко говоря, «без энтузиазма», с обидой или даже «в штыки», как «наглое посягательство» на «непререкаемые авторитеты» и «безусловные и уже сложившиеся аксиомы».
В связи с этим пока неописанными и неосознанными научной общественностью наверняка остаются важнейшие закономерности, связанные с организацией и регуляцией жизнедеятельности рептилий.
Так же неблагополучно обстоят дела и с пониманием такого важнейшего аспекта жизни рептилий, как их термальные адаптации. Им посвящено много научных публикаций, сравнительно недавно даже вышла отдельная книга по этой теме (Angilletta, 2009). Однако на деле получается, что в них рассматриваются отдельные стороны биологии рептилий (экологической физиологии, физиологии, экологии, общей термобиологии рептилий и т. п.), но к самим адаптациям они имеют очень мало отношения. Здесь, на наш взгляд, важно понять, что множество закономерностей, зависимостей и связей различных явлений и феноменов в биохимии, физиологии, экологии и других областях биологии рептилий с температурой реально существуют и не вызывают сомнений. Проблема не в том, что их нет и что мы их отвергаем. Проблема в том, что далеко не все эти зависимости и связи могут быть квалифицированы как термальные адаптации (Черлин, 2015). Скорее всего, исследуя термальные адаптации, нужно было бы заниматься, с одной стороны, биохимическими и физиологическими термальными компенсациями, а с другой – двумя важнейшими аспектами явления адаптации: 1) комплексом стабильных, неизменных, видоспецифичных характеристик термального гомеостаза и 2) набором и принципами поведенческих реакций, тонко «подстраивающих» пространственно-временную структуру суточной и сезонной активности в направлении сохранения неизменности комплекса характеристик термального гомеостаза в разных условиях среды.
Было выполнено и опубликовано некоторое количество работ, посвященных принципам организации у рептилий механизмов терморегуляции, которые иногда описываются разными математическими моделями (Brattstrom, 1965; Heath, 1970; Berk, Heath, 1975; Barber, 1977; Barber, Crawford, 1977; Barber, 1979; Huey, 1982; Adolph, 1990; Adolph, Porter, 1993 и др.). Эти и другие публикации действительно слегка приподнимают завесу над данной темой, описывают некоторые внешние проявления механизмов терморегуляции. Но, несмотря на всю их серьезность, к сожалению, они не вскрывают самих механизмов этого феномена. Мало того, вследствие отсутствия общей концепции термобиологии рептилий, у специалистов до сих пор не сложилось представления о том, какую, собственно, температуру рептилии регулируют, для чего, в каких случаях и т. п. Совершенно очевидно, что температуру тела эти животные регулируют не всегда. Да и само явление регуляции температуры тела на самом деле оказывается неоднородным: иногда рептилии поддерживают температуру тела в некотором диапазоне, ограниченном сверху и снизу опасными (или неблагоприятными, «отталкивающими») границами; иногда они поддерживают некоторый уровень температур (оптимальный, предпочитаемый, «eccritic» и т. п., а точнее – уровень температур термостабилизации (Черлин, 2014а; Cherlin, 2015b)), но этот тип регуляции проявляется не всегда; поддерживают они и определенную температуру тела в состояниях неактивности (в периоды сна, гибернации, эстивации). Но до сих пор неясно: одинаков ли уровень поддерживаемых животными температур в разных физиологических состояниях и других случаях? Нет даже единого, общепринятого списка тех термобиологических параметров, которые затронуты механизмами регуляции; не проясняется также и то, одними и теми же механизмами поддерживаются у рептилий разные термобиологические параметры или нет... Читая многие даже очень серьезные публикации, становится очевидно, что этими важнейшими вопросами ученые чаще всего даже не задаются и, следовательно, не пытаются на них ответить. А без этого не будет понята и вся система отношений жизнедеятельности рептилий с температурным фактором.
Также непросто обстоят дела и с проблемами эволюции термобиологической сферы. Такие важнейшие и вроде бы соответствующие современному пониманию эволюции утверждения, как, в частности, то, что изменение термальной среды влечет за собой соответствующий адаптивный сдвиг термобиологической нормы животных в популяции, на самом деле неочевидны, и, если оно действительно существует, то требует специальных, экспериментальных или других доказательств. Наши исследования как раз показывают спорность такого утверждения (Черлин, 2015). Совершенно очевидно, что в разных географических регионах и климатических условиях могут существовать рептилии с очень сходными или просто одинаковыми основными термофизиологическими характеристиками активности, а в одних и тех же географических регионах и климатических условиях – рептилии с сильно различающимися характеристиками. И это – благодаря использованию разных пространственно-временных структур суточной и сезонной активности. Кроме того, мы четко показали, как система поведенческой модификации пространственно-временной структуры активности чрезвычайно эффективно препятствует любым изменениям термофизиологической видовой нормы. Собственно, эти поведенческие реакции на то и направлены, чтобы сохранять неизменными характеристики термофизиологической видовой нормы. В такой ситуации сложно предположить возможность адаптивного изменения видовой термофизиологической нормы, неизменность которой так надежно защищена. Но изменение этой нормы все-таки происходит. На наш взгляд, это может быть следствием, например, каких-то неадаптивных чисто биохимических, физиологических, морфологических, генетических и пр. изменений в нервном механизме регуляции температур и т. п., о которых мы еще не знаем. Вместе с тем система поведенческих модификаций, изменяя время и место активности животных, «подгоняет», адаптирует новую физиологическую норму к конкретным климатическим условиям региона или заставляет животных с этой новой нормой сменить биотопическое распределение и/или даже географическое распространение (т. е. изменить временное и пространственное размещение). Тогда отбор способствует сохранению удачных сочетаний новой видоспецифичной нормы, комплекса поведенческих реакций и внешних условий. В таком случае новая видовая норма оказывается действительно хорошо адаптированной к климатическим условиям региона, который животное занимает, т. е. процесс адаптации происходит прежде всего за счет поведенческих реакций. Но при этом вряд ли можно утверждать, что новая видовая норма появилась в результате адаптации к данным климатическим условиям.
Кроме того, к сожалению, у исследователей, герпетологов, ослабевает здоровый и очень важный для зоологов натурализм, т. е. они перестают быть настоящими натуралистами. Порой неумеренное стремление завести полученные первичные данные в различные модели уводит обсуждение важнейших сторон жизнедеятельности животных в плоскость математических формул. Эти модели становятся часто самоцелью, а сравнение моделей превращается в «игры разума», слабо соприкасающиеся с реальной жизнедеятельностью рептилий. Обращаю внимание на то, что мы категорически не выступаем против математической обработки данных и применения математических моделей. Просто математическая обработка должна быть подспорьем, иногда – очень важным, но все-таки именно подспорьем для ученого на пути понимания того, как и почему животные живут именно так, а не иначе. Но порой бывает важнее и ценнее увидеть какой-то мелкий элемент поведения животного (что может заметить только хороший натуралист), открывающий целый пласт его бытия, чем, например, сравнивать теоретические модели регуляции каких-нибудь физиологических параметров, плохо представляя, по каким реальным законам организована физиологическая система и экология этих животных и в чем эти законы проявляются.
В качестве примера важности тонких натуралистических наблюдений можно привести описание слабо, неявно проявляющихся поведенческих особенностей, позволивших выявить у рептилий состояние полной активности и вскрывших, вслед за этим, целый пласт предельно важных термофизиологических процессов, феноменов, характеристик, закономерностей (Либерман, Покровская, 1943; Cowles, Bogert, 1944; Cherlin, 2015b; Черлин, 2016). А в качестве примера очень удачного применения математического моделирования, не оторванного от объекта исследования, можно привести работы А. В. Коросова (2008, 2010) по моделированию терморегуляции обыкновенной гадюки, Pelias berus.
***
Несмотря на то, что начиная с 1940-х гг. было опубликовано несколько книг и сводок по термобиологии рептилий или касающихся этой темы (Cowles, Bogert, 1944; Brattstrom, 1965; Tempelton, 1970; Cloudsley-Thompson, 1971; Comparative…, 1973; Schmidt-Nielsen, 1964, 1997 и др.) и на обилие более частных публикаций, к сожалению, общепринятого системного понимания этой области науки до сих пор не существует. Пока исследования, анализ материалов и выводы в значительной мере определяются давно установившимися методами и понятиями и идут по традиционной, накатанной колее (с использованием порой красивых и современных технических средств и методов обработки материала, «автоматически» обеспечивающих для многих исследователей безусловное принятие научной общественностью полученных данных и публикаций). Чтобы из этого положения, наконец, выйти, необходимо внести в исследования «свежую струю».
Таким образом, можно заключить, что само по себе обилие публикаций по термобиологии рептилий не обеспечивает наличия объективной картины этого научного направления. Необходимо сформулировать, наконец, общую концепцию в термобиологии рептилий, стандартизировать и научно обосновать систему понятий, терминов и показателей, методов сбора и обработки материалов и т. п., чему научное «термобиологическое сообщество» пока активно сопротивляется, явно ошибочно считая, что либо это все уже есть, либо что можно обойтись и без этого.
Нам кажется, что в данном случае именно описанные нами проблемы этой области науки лучше и четче характеризуют состояние изученности термобиологии рептилий, чем перечисление отдельных достижений, не связанных между собой и не объединенных пока единой идеей.
[1] Полная активность – животные находятся в физиологическом состоянии, в полной мере, без существенных физиологических ограничений, обеспечивающем эффективное протекание всех основных поведенческих актов (локомоция, терморегуляция, питание, сексуальность, территориальность, защита, коммуникации и т. п.). В этом состоянии движения рептилий активны, быстры (адекватно видовым особенностям) и точны (Либерман, Покровская, 1943; Dawson, 1975; Cowles, Bogert, 1944; Черлин, 2014б; Cherlin, 2015a).
[2] Температуры (диапазон) термостабилизации – довольно узкий диапазон высоких температур тела, который рептилии, находясь в состоянии полной активности, направленно и оперативно поддерживают у себя в течение более или менее продолжительного времени. В цифровом выражении он почти соответствует тому, что ранее назвали в литературе «оптимальными» или «предпочитаемыми» температурами, определенным методически корректным способом (что встречалось, увы, нечасто), но по сути этот диапазон чаще всего не является ни оптимальным ни для каких физиологических процессов в организме, ни предпочитаемым, поскольку в основном это компромисс между желаемым и возможным уровнями температур. Просто рептилии по ряду причин (пока не до конца понимаемых) стабилизируют в нем температуру своего тела. Поэтому мы нашли разумным и адекватным изменить его название на «диапазон термостабилизации», убрав из него оценку причин этой стабилизации (Черлин, 2014а).
Библиография
Коросов А. В. Простая модель баскинга обыкновенной гадюки (Vipera berus L.) // Современная герпетология. 2008. Т. 8. Вып. 2. С. 118–136.
Коросов А. В. Экология обыкновенной гадюки на Севере . Петрозаводск: Изд-во ПетрГУ, 2010. 264 с.
Либерман С. С., Покровская И. В. Материалы по экологии прыткой ящерицы // Зоологический журнал. 1943. Т. 22. № 2. С. 247–256.
Cтрельников И. Д. Свет как фактор в экологии животных. Действие солнечной радиации на температуру тела некоторых пойкилотермных животных . // Известия Научного института имени П. Ф. Лесгафта. 1934. Т. 17–18. С. 313–372.
Черлин В. А. Термобиология рептилий. Общие сведения и методы исследований (руководство) . СПб.: Русско-Балтийский информационный центр «БЛИЦ», 2010. 124 с.
Черлин В. А. Сложности и возможные ошибки при полевых исследованиях по термобиологии рептилий // Современная герпетология: проблемы и пути их решения: Статьи по материалам докладов Первой международной молодежной конференции герпетологов России и сопредельных стран (Санкт-Петербург, Россия, 25–27 ноября 2013 г.) / Зоологический институт РАН. СПб., 2013. С. 32–39.
Черлин В. А. Рептилии: температура и экология . Saarbrücken: Lambert Academic Publishing, 2014а. 442 с.
Черлин В. А. О статье С. С. Либерман и Н. В. Покровской по термобиологии прыткой ящерицы (1943 г.) // Принципы экологии. 2014б. Т. 3. № 3. С. 25–32.
Черлин В. А. Тепловые адаптации рептилий и механизмы их формирования // Принципы экологии. 2015. T. 4. № 1. С. 17–76.
Черлин В. А. Современный взгляд на термобиологию с позиции изучения рептилий // Биосфера. 2016. Т. 8. № 1. С. 47–67.
Adolph S. C. Influence of behavorial thermoregulation on microhabitat use by two Sceloporus lizards // Ecology. 1990. No 71. P. 315–327.
Adolph S. C., Porter W. P. Temperature, activity, and life histories // The American Naturalist. 1993. Vol. 142. No 2. P. 273–295.
Agassiz L., Gould A. A. Principles of zoology: Touching the structure, development, distribution and natural arrangement of the races of animals, living and extinct. Part I. Comparative physiology. Sheldon, Blakeman and Co., New York, 1856. 250 p.
Angilletta M. J. Thermal Adaptation: A Theoretical and Empirical Synthesis. Oxford: Oxford University Press, 2009. 289 р.
Atsatt S. R. Color changes as controlled by temperature and light in the lizards of the desert regions of southern California // Univ. Calif. Los Angeles Publ. Biol. Sci. 1939. No 1. P. 237–276.
Bailey R. M. Winter mortality in the snake Storeria dekay // Copeia. 1948. No 3.
Bailey R. M. Temperature toleration of garter snakes in hibernation // Ecology. 1949. Vol. 30. P. 238–242.
Baldwin F. M. Body temperature changes in turtles and their physiological interpretations (Chrysemys marginata belli, C. Gray and Chelydra serpentina, Lin.) // Amer. Jour. Physiol. 1925а. No 72. P. 210–211.
Baldwin F. M. The relation of body to environmental temperatures in turtles Chrysemys marginata belli (Gray) and Chelydra serpentina (Lin.) // Biol. Bull. 1925b. No 48. P. 432–445.
Barber B. J. A theoretical and experimental analysis of behavioral temperature regulation in the lizard, Dipsosaurus dorsalis // Dissertation Abst. Int. 37. 1977. Р. 4330.
Barber B. J. Dual threshold control of peripheral temperature in the lizard Dipsosaurus dorsalis // Physiol. Zool. 1979. Vol. 52. P. 250–263.
Barber B. J., Crawford E. C. Jr. A stochastic dual-limit hypothesis for behavioral thermoregulation in lizards // Physiol. Zool. 1977. Vol. 50. No 1. P. 53–60.
Benedict F. G. The physiology of large reptiles with special reference to the heat production of snakes, tortoises, lizards and alligators. Washington: Carnegie Institution, 1932. No 425. P. 539.
Bennett A. F. A comparison of activities of metabolic enzymes in lizards and rats // Comp Biochem. Physiol. 1972. 42B. P. 637–647.
Bennett A. F., Hicks J. W., Cullum A. J. An experimental test of the thermoregulatory hypothesis for the evolution of endothermy // Evolution. 2000. Vol. 54. No 5. P. 1768–1773.
Bennett A. F., Ruben J. A. Endothermy and activity in vertebrates // Science. 1979. Vol. 206. P. 649–654.
Berk M. L., Heath J. E. An analysis of behavioral thermoregulation in the lizard, Dipsosaurus dorsalis // J. Therm. Biology. 1975. No 1. P. 15–22.
Bligh J., Johnson K. G. Glossary of terms for thermal physiology // J. Appl. Physiol. 1973. No 35. P. 941–961.
Blum H. F., Spealman C. R. Note on the killing of rattlesnakes by "sunlight" // Copeia. 1933. P. 150–151.
Bogert C. M. Reptiles under the sun // Nat. Hist. 1939. Vol. 24. P. 26–37.
Bogert C. M. Thermoregulation in reptiles, a factor in evolution // Evolution. 1949а. No 3. P. 195–211.
Bogert C. M. Thermoregulation and eccritic body temperatures in Mexican lizards of the genus Sceloporus // Ann. Inst. Biol. Mexico. 1949b. No 20. P. 415–426.
Bogert C. M. Body temperatures of the tuatara under natural conditions // Zoologica. 1953. No 38. P. 63–64.
Bogert C. M. How reptiles regulate their body temperature // Scientific American. 1959. No 200. P. 105–120.
Brattstrom В. Н. Body temperatures of reptiles // Amer. Midl. Naturalist. 1965. Vol. 73. No 2. P. 376–492.
Buxton P. A. Animal life in deserts: A study of the fauna in relation to environment. London: Edward Arnold and Co., 1923. 176 p.
Buxton P. A. Heat, moisture and animal life in deserts // Proc. Roy. 1924. Soc. B 96. P. 123–131.
Camp C. L. Notes on the local distribution and habits of the amphibians and reptiles of southeastern California in the vicinity of the Turtle Mountains // Univ. Calif. Publ. Zool. 1916. Vol. 12. P. 503–544.
Carpenter C. C. Comparative ecology of the common garter snake (Thamnophis s. sirtalis), the ribbon snake (Thamnophis s. sauritus), and Butler's garter snake (Thamnophis butleri) in mixed populations // Ecol. Monogr. 1952. Vol. 22. P. 235–258.
Carpenter C. C. Body temperatures of three species of Thamnophis // Ecology. 1956. Vol. 37. P. 732–735.
Cherlin V. A. The activity statuses and their significance in physiological ecology of reptiles and other ectothermic animals // Some important aspects of thermal biology of reptiles. Saarbrücken, Lambert Academic Publishing, 2015a. P. 11–54.
Cherlin V. A. Why the complex of thermobiological characters is so and not another? // Some important aspects of thermal biology of reptiles. Saarbrücken, Lambert Academic Publishing, 2015b. P. 55–116.
Cloudsley-Thompson J. L. The Temperature and Water Relations of Reptiles. Merrow Publishing Co., Ltd., Watford, Herts., England, 1971. 159 p.
Colbert E. H., Cowles R. B., Bogert C. M. Temperature tolerances in the American alligator and their bearing on the habits, evolution and extinction of the dinosaurs // Bull. Am. Mus. Nat. Hist. 1946. No 86. P. 329–373.
Comparative animal physiology. (L. Prosser, Ed.). Philadelphia, London, Toronto: W. B. Saunders Company, 1973. 888 p.
Cowles R. B. Possible implications of reptilian thermal tolerance // Science. 1939. Vol. 90. P. 465–466.
Cowles R. B. Additional implications of reptilian sensitivity to high temperature // Amer. Nat. 1940. Vol. 75. P. 542–561.
Cowles R. B. Winter activities of desert reptiles // Ecology. 1941. Vol. 22. P. 125–140.
Cowles R. B. Critical thermal levels and thermal regulation in desert reptiles // 25th Ann. Meeting Amer. Soc. Ichthyologists and Herpetologists, Amer. Mus. Nat. Hist. Abstr. 1942. No 10 (mimeographed).
Cowles R. B. Parturition in the yucca night-lizard, Xantusia vigilis // Copeia. 1944. No 1. P. 98–100.
Cowles R. B. Heat-induced sterility and its possible bearing on evolution // Ibid. 1945. Vol. 79. P. 160–175.
Cowles R. B. Sidewinding locomotion in snakes // Copeia. 1956. No 4. P. 211–214.
Cowles R. B. Possible origin of dermal temperature regulation // Evolution. 1958а. No 12. P. 347–357.
Cowles R. B. The effect of temperature on thyroid histology and survival in the lizard, Sceloporus occidentalis // Copeia. 1958b. P. 265–276.
Cowles R. B. Semantics in biothermal studies // Science. 1962. No 135. P. 670.
Cowles R. B., Bogert C. M. Preliminary study of the thermal requirements of desert reptiles // Bulletin of the American Museum of Natural History. 1944. No 83. P. 261–296.
Cowles R. B., Burleson G. L. The sterilizing effect of high temperature on the male germ-plasm of the yucca night lizard, Xantusia vigilis // Am. Nat. 1945. Vol. 79. P. 417–435.
Dawson W. R. On the physiological significance of the preferred body temperatures of reptiles // Perspectives of biophysical ecology. Ecological studies. Vol. 12. (D. M. Gates and R. B. Schmerl, Eds.). New York, 1975. P. 443–473
Evans E. E., Cowles R. B. Effect of temperature on antibody synthesis in the reptile Dipsosaurus dorsalis // Proc. Soc. Exp. Biol. Med. 1959. No 101. P. 482–483.
Fitch H. S. Life history and ecology of the five-lined skink, Eumeces fasciatus // Univ. Kansas. Publ. Mus. Nat. Hist. 1954. No 8. P. 1–156.
Fitch H. S. Temperature responses in free-living amphibians and reptiles of northeastern Kansas // Univ. Kansas Publ. Mus. Nat. Hist. 1956. No 8. P. 417–476.
Fox W. Effect of temperature on development of scutellation in gater snake, Thamnophis elegans atratus // Copeia. 1948. No 4. P. 252–262.
Franz Н. Untersuchungen über den Wärmebaushalt der Poikilothermen // Biolog. Zentralblatt. 1930. Bd. 50. S. 158–181.
Gadow H. Amphibia and reptiles // The Cambridge Natural History (C. S. F. Harmer and A. E. Shipley, Eds.). London: Mc-Millan and Co, 1901. Vol. 8. P. 1–668.
Grinnell J. The biota of the San Bernardino Mountains // Univ. Calif. Publ. Zool. 1908. No 5. P. 1–170.
Hall F. G. The vital limit of exsiccation of certain animals // Biol. Bull. 1922. Vol. 42. P. 31–51.
Hardy J. D. The "set-point" concept in physiological temperature regulation // Physiological controls and regulations. Philadelphia, 1965. P. 98–116.
Heath J. E. Behavioral regulation of body temperature in poikilotherms // The Physiologist. 1970. Vol. 13. P. 339–410.
Herter K. Die Vorzungstemperaturen bel Landtieren // Naturwissenschaften. Berlin, 1941. Vol. 29. P. 155–164.
Hesse R. Tiergeographie auf oekologischer grundlage. Jena: Gustav Fischer, 1924. 613 p.
Hesse R., Allee W. C., Schmidt K. P. Ecological animal geography. New York: J. Wiley and Sons, 1937. 597 p.
Huey R. Temperature, physiology, and the ecology of reptiles // Biology of the Reptilia. Physiology (C). Vol. 12. (C. Gans and F. H. Pough, Eds.). London: Academic Press, 1982. P. 25–91.
Inger R. F. Temperature responses and ecological relations of two Bornean lizards // Ecology. 1959. Vol. 40. No 1. P. 127–136.
Kammerer P. Der Artenwandel auf Inseln. Vienna; Leipzig: Franz Deuticke, 1926. 324 p.
Kanitz A. Temperatur und Lebens-Vorgange im Allgemeinen // Tabulae Biol. 1925. Vol. 2. P. 9–25.
Klauber L. M. Studies of reptile life in the and southwest // Bull. Zool. Soc. San Diego. 1939. Vol. 14. P. 1–100.
Klauber L. M. Four papers on the application of the statistical methods to herpetolo-gical problems. III. The correlation between scalation and life zones in San Diego County snakes // Bull. Zool. Soc. San Diego. 1941. Vol. 17. P. 73–79.
Krüger P., Kern H. // Arch. Physiol. 1924. No 202. S. 119–138.
Krüger P. Uber die Bedeutung der ultraroten Strahlen für den Wärmehaushalt der Poikilothermen // Biol. Zbl. 1929. Vol. 49. P. 65–82.
Lamarre-Picquot P. Troisieme memoirs sur l’incubation et qutre phenomenes observes ches les ophidiens // Comp. Rend., Acad. Sci. Paris, 1842. Vol. 14. P. 164.
Lowe C. Notes on the mating of desert rattlesnakes // Copeia. 1942. P. 261–262.
Lowe C. H., Vance V. J. Acclimation of the critical thermal maximum of the reptile Urosaurus ornatus // Science. 1955. No 122. P. 73–75.
Lueth F. X. Effects of temperature on snakes // Copeia. 1941. P. 125–132.
Lufti M. Das termotaktische Verhalten einiger Reptilien: Inaugural-Diss. Munchen, 1936.
McGinnis S. M., Moore R. G. Thermoregulation in the Boa constrictor // Herpetologica. 1969. No 25. P. 38.
Mcilhenny E. A. The alligator's life history. Boston: The Cristopher Publishing House, 1935. 117 p.
Mosauer W. A note on the sidewinding locomotion of snakes // Amer. Nat. 1930. Vol. 64. P. 178-183.
Mosauer W. The toleration of solar heat in desert reptiles // Ecology. 1936. Vol. 17. P. 56–66.
Mosauer W., Lazier E. L. Death from insolation in desert snakes // Copeia. 1933. P. 149.
Musacchia X. J. The viability of Chrysemys picta submerged at various temperatures // Physiological Zoology. 1959. No 32. P. 47–50.
Nelson D. O., Heath J. E., Prosser C. L. Evolution of temperature regulatory mechanisms // Amer. Zool. 1984. No 24. P. 791–807.
Norris K. S. The ecology of the desert iguana Dipsosaurus dorsalis // Ecology. 1953. No 34. P. 256–287.
Parker G. H. The time of submergence necessary to drown alligators and turtles // Occas. Papers Boston Soc. Nat. Hist. 1925. Vol. 5. P. 157–159.
Parker H. W. A new melanic lizard from Transjordania, and some speculations concerning melanism // Proc. Zool. Soc. London, 1935. P. 137–142.
Parker H. W. The lizard genus Aprasia; its taxonomic and temperature correlated variation // Bulletin of the British Museum of Natural History. 1956. No 3. P. 363–385
Pearse A. S. Animal ecology. New York: McGraw-Hill, 1931. 642 p.
Pearse A. S., Hall F. G. Homoiothermism: The origins of warm-blooded vertebrates. New York: J. Wiley and Sons, 1928. 119 p.
Pearson O. P. Habits of the lizard, Liolaemus multiformis multiformis at high altitudes in southern Peru // Copeia. 1954. No 2. P. 111–116.
Pearson O. P., Brandford D. F. Thermoregulation of lizards and toad at high altitudes in Peru // Copeia. 1976. No 1. P. 155–170.
Reese A. M. Some reactions of Alligator mississippiensis // Jour. Comp. Psychol. 1923. Vol. 3. No 1. P. 51–59.
Reichling H. Transpiration und Vorzugstemperatur mitteleuropiiischer Reptilien und Amphibien // Zool. Jb. (Syst.). 1957. Vol. 67. P. 1–64.
Richet Ch. La chaleur animale. (F. Alcan, Ed.). Paris, 1889. 307 p.
Saint-Girons H., Saint-Girons M. C. Cycle d'activite et thermoregulation chez les reptiles (lezards et serpents) // Vie Milieu. 1956. No 7. P. 133–226.
Schmidt-Nielsen K. Desert animals: Physiological problems of heat and water. Oxford: Clarendon Press, 1964. 277 p.
Schmidt-Nielsen K. Animal physiology. New York: Cambridge University Press, 1997. 607 p.
Soetbeer F. Ueber die Korperwarme der poikilothermen Wirbelthiere // Arch. Exp. Pathol. Pharmakol. 1898. Vol. 40. P. 53–80.
Sutherlanad A. The temperature of reptiles, monotremes and marsupials // Proceedings of the Royal Society of Victoria. 1896. No 9. P. 57–67.
Templeton J. R. Reptiles // The Comparative Physiology of Thermoregulation. (G. C. Whittow, Ed.). New York; London: Academic Press, 1970. Vol. 1. P. 167–221.
Valensiennes A. Obsrevations faites pendant l’incubation d’une femelle du python a deux raies (Pythin bivittatus, Kuhl.) pendant les mois de mai et de juin 1841 // Comp. Rend., Acad. Sci. Paris, 1841. Vol. 13. P. 126–133.
Warden C. J., Jenkins T. N., Warner L. H. Comparative psychology. Vertebrates. New York: The Ronald Press, 1936. 560 p.
Weese A. O. An experimental study of the reactions of the horned lizard, Phrynosoma modestum Gir., a reptile of the semi-arid desert // Biol. Bull. 1917. No 32. P. 98–116.
Weese A. O. Environmental reactions of Phrynosoma // Amer. Naturalist. 1919. No 53. P. 33–54.
Weigmann R. Die wirkung starker abkuhlung auf amphibien und reptilian // Zeitschrift fiir Wissenschaffliche Zoologie. 1929. P. 1–692.
Werner F. Beiträge zur Kenntnis der Reptilien und Amphibien von Istrien und Dalmatien // Verh. zool.-bot. Ges. Wien. 1891. No 41. S. 751–768.
Woodbury A. M., Hardy R. Studies of the desert tortoise Gopherus agassizii // Ecol. Monogr. 1948. Vol. 18. P. 145–200.

 © 2011 - 2026
© 2011 - 2026