



Введение
У большинства видов птиц в населенных пунктах наблюдаются заметные отличия от природных популяций тех же видов в самых разных аспектах биологии. Некоторые из них представляют собой высокоэффективные адаптации к обитанию в антропогенном ландшафте и присутствию человека, другие являются реакциями на определенные особенности антропогенного ландшафта, но не носят адаптивного характера. Различия этих категорий зачастую трудновыявляемы, поскольку неадаптивные формы поведения могут сопровождаться повышением репродуктивного успеха вследствие не связанных с ними факторов, столь же обычной является и обратная ситуация. Поэтому обнаружение подобных различий в естественных и синантропных популяциях, хотя и свидетельствует о формировании особых городских популяций и начале процесса синантропизации вида, не всегда является предпосылкой успешной синантропизации. Поэтому целью нашего исследования было изучение репродуктивных отличий природных и синантропных популяций и определение адаптивности наблюдаемых изменений гнездования птиц. Модельным объектом послужила зарянка (Euithacus rubecula) – обычный в Рязанской области представитель семейства дроздовых (Turdidae). Вид обитает во многих населенных пунктах, в первую очередь, на территориях, занятых зелеными насаждениями (Птицы городов, 2001; Ильичев и др., 1987; Прокофьева, 2006; Симкин, 1990; Saemann, 1970, 1973). Даже на территории небольшого парка, при наличии подходящих микростаций, зарянка может успешно гнездиться. Пригодные для гнездования участки существуют и в черте застройки. Задачи исследования включали сравнительный анализ особенностей гнездования и репродуктивного успеха, выявление специфики гнездования городских популяций и их адаптивности.
Материалы
Репродуктивную биологию зарянки мы изучали в 2000–2017 годах на базе Рязанского государственного университета в естественных и антропогенных стациях Рязанской области. Изучение гнездовой биологии зарянок производили по общепринятым методикам (Нумеров и др., 2010). Описывали тип расположения гнезда, высоту над землей, способ, которым оно было найдено. Регистрировали количество и размеры яиц в кладке. Массу яиц определяли только в гнездах с ненасиженными кладками – не позднее двух суток после начала насиживания. Если в момент нахождения гнезда стадию насиживания определить не было возможности, его начало рассчитывали по срокам вылупления птенцов. При дальнейших наблюдениях определяли редукцию кладки и выводка, по возможности устанавливали причины гибели. Обнаружено и обследовано 149 гнезд, из них судьбу 136 гнезд удалось проследить со стадии откладки яиц или насиживания до вылета птенцов.
Методы
Все полученные данные мы условно подразделили на материалы по естественным биотопам (сюда включили удаленные от города и пригородные леса, а также лесопарк – крупный пригородный лесной массив) и антропогенным стациям (мелкие внутригородские парки и лесопосадки, дачные поселки, селитебный ландшафт). Материалы по каждой группе стаций обрабатывались отдельно. Рассчитывали следующие показатели: размер полной кладки и лимиты, долю разоренных гнезд, вероятность разорения гнезда за сутки для каждой стадии репродуктивного цикла, репродуктивный успех (отношение числа слетков к количеству отложенных яиц). Смертность потомства рассчитывали (для стадий насиживания и выкармливания гнездовых птенцов) от количества особей, доживших до начала данной стадии репродуктивного цикла и от начального количества отложенных яиц. Сравнительный анализ производили при помощи коэффициента корреляции Пирсона и Т-критерия, кроме того, рассчитывали стандартное отклонение и ошибку средней (m).
Результаты
Зарянки демонстрируют чрезвычайную пластичность в расположении гнезд, которые могут находиться в укрытиях, в том числе антропогенного происхождения, или быть совершенно открытыми, располагаться на высоте от 3–4 метров до уровня почвы и ниже. В различных регионах все показатели репродуктивной биологии зарянок широко варьируют (Благосклонов, 2017; Головань, 1997; Зимин, 2009; Кладки и размеры..., 1995; Лебедева, Ломадзе, 2007; Симкин, 1990; Сотников, 2008; Шепель, Зиновьев, 1999).
Тип расположения гнезда
Зарянка – почти исключительно закрытогнездящийся вид. Расположенные выше уровня земли гнезда почти всегда находятся в каких-либо нишах растительного или антропогенного субстрата. Отступлением от этого правила были только два случая гнездования в густом пучке пристволовой поросли (в одном и том же месте два года подряд, что позволяет предположить повторное гнездование тех же особей). Однако и в этом случае гнезда были укрыты с трех сторон и хорошо замаскированы. Все остальные надземные гнезда располагались в дуплах и полудуплах, в углублениях в торцах пней, в антропогенных укрытиях. В дуплах и нишах стволов располагалось 53.0 % гнезд, что позволяет считать этот способ гнездования основным для рязанских зарянок. Избирательности в отношении древесных пород у зарянки не наблюдается. При наличии удобного дупла на подходящей высоте птицы поселяются на любых древесных породах. Около четверти всех гнезд было расположено на уровне почвы или ниже. Их можно также разделить на несколько групп. Наиболее обычно гнездование в стенках крутых склонов и обрывчиков (16 гнезд), при этом гнездо укрыто с трех сторон, сверху и снизу. Реже (12 случаев) гнезда располагаются на сравнительно пологих склонах или ровных участках, обычно в основании куста или ствола, под кучкой веток, пучком травы или лежащим предметом, нередко в ямках глубиной 10–35 см. В антропогенных стациях 10 гнезд было найдено в антропогенных укрытиях на уровне земли (в основном старые банки из-под краски). Среди антропогенных укрытий, приподнятых над землей, зарянки использовали горизонтальные железные трубы, дуплянку и скворечник, открытые сбоку железный и бетонный фонарные столбы, располагали гнезда между стеной здания и кустом, между деревянным забором и кустом, под крышей и между рамами разбитого окна дачного домика. В населенных пунктах в укрытиях антропогенного происхождения (в том числе наземных) располагались более половины гнезд.
Высота гнездования
Антропогенная нагрузка на высоту расположения гнезд влияла очень слабо. Вероятно, отсутствие связи высоты расположения гнезда со степенью антропогенной трансформации местообитаний связано у зарянки с преобладанием гнездования в дуплах, когда птицы вынуждены занимать имеющиеся укрытия, невзирая на их высоту. По нашим наблюдениям, у зарянки не происходит изменения стереотипа гнездования в связи с антропогенной нагрузкой (за исключением использования антропогенных укрытий). Среди 44 гнезд, найденных в населенных пунктах, 13 было расположено на земле, что даже несколько выше частоты наземного гнездования в естественных стациях.
Кладки и размеры яиц
В полных кладках зарянки мы находили от 3 до 9 яиц. В среднем на гнездо с полной кладкой пришлось 6.0 ± 1.03 яйца. Три яйца было зарегистрировано только в кладках, найденных в конце репродуктивного периода. Вероятно, это вторые кладки, или отложенные взамен погибших. Как правило, полная кладка включает 6 яиц, реже 7, 5, или 8. В двух кладках оказалось 9 яиц. Значимых отличий по средним значениям в естественных и антропогенных ландшафтах не обнаружено. Так, в населенных пунктах кладки содержат 5.9 ± 1.08 яица, а в естественных лесах – 6.1 ± 0.99. Анализ распределения показывает, что в населенных пунктах относительно выше доля больших и маленьких кладок, преобладание кладок среднего размера не столь выражено, как в естественных стациях (рис. 1). Полученные нами данные в целом соответствуют таковым для зарянки в других частях ареала, где в полных кладках также содержится от 3 до 9 яиц, чаще 6–7 (Болотников и др., 1969; Зимин, 2009; Ильичев и др., 1987; Лебедева, Ломадзе, 2007; Прокофьева, 2006; Сапетина, 2009; Сотников, 2008; Фуфаев, 1982; Хазиева и др., 1975).
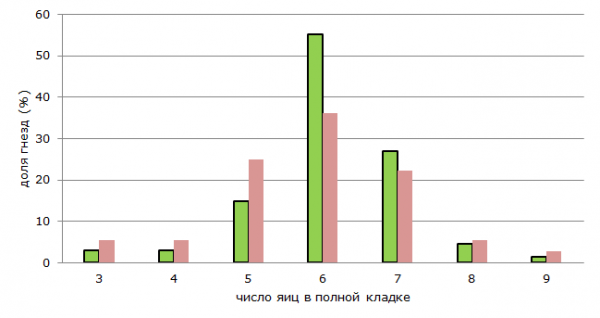
 – естественные стации (natural stations), n = 73;
– естественные стации (natural stations), n = 73;  – антропогенные стации (anthropogenic stations), n = 37
– антропогенные стации (anthropogenic stations), n = 37
Рис. 1. Распределение кладок зарянки с разным количеством яиц в естественных и антропогенных стациях
Fig. 1. Distribution of robin's clutches with different number of eggs in natural and anthropogenic stations
Параметры яиц в изученных гнездах оказались следующими: В антропогенном ландшафте длина 19.5 ± 0.37 мм, ширина – 15.3 ± 0.48 мм, масса – 2.6 ± 0.08 г (n = 21). В естественных лесах эти же показатели составили соответственно 18.6 ± 0.34, 14.3 ± 0,42 и 2.1 ± 0,06 (n = 26). На уровне тенденции в первом случае характерен более крупный размер, хотя отличия статистически не значимы (Т критерий 1.7 и 1.5 соответственно). Для яиц из естественных стаций характерна и меньшая вариация размеров.
Сроки размножения
Анализ сроков размножения зарянок показывает, что как в естественных, так и в антропогенных стациях для значительного числа пар характерны два цикла размножения. Попытки повторного гнездования после потери первых гнезд несколько сглаживают четкое распределение кладок по двум циклам, поскольку у таких пар бывает лишь одна успешная кладка в году. В случае повторного неуспеха может быть и три попытки гнездования за сезон. В антропогенных стациях первые кладки появляются раньше, а последние – позже, чем в естественных. Поздние случаи гнездования в этом случае нельзя объяснить большим, чем в естественных лесах, числом повторных кладок, поскольку общий репродуктивный успех популяции существенно не отличается, а график, наоборот, показывает, что вторые кладки более распространены в естественных стациях.
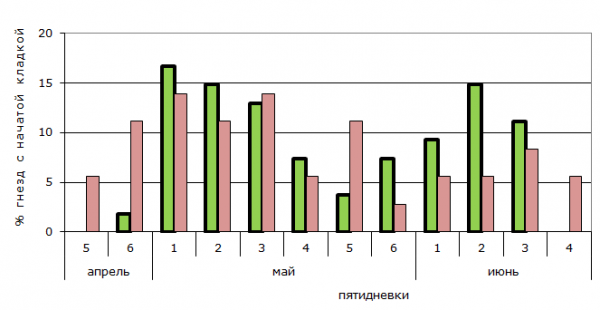
 – естественные стации (natural stations);
– естественные стации (natural stations);  – антропогенные стации (anthropogenic stations)
– антропогенные стации (anthropogenic stations)
Рис. 2. Сроки размножения зарянок в естественных и антропогенных стациях
Fig. 2. Periods of reproduction of the robin in natural and anthropogenic stations
Репродуктивные потери
Репродуктивные показатели зарянок в естественных и антропогенных стациях представлены в табл. 1.
Таблица 1. Некоторые репродуктивные показатели зарянок (2000–2017 гг.)
| Показатель | Естественные стации (n = 93) | Населенные пункты (n = 44) |
| Средний размер кладки (x ± σ) | 6.1 ± 0.99 | 5.9 ± 1.08 |
| ошибка средней (m) | 0.10 | 0.16 |
| лимиты | 3–9 | 3–9 |
| Отход яиц на стадии откладки, % | 6.86 | 1.17 |
| Смертность потомства при насиживании (от числа доживших до его начала яиц), % | 20.92 | 19.47 |
| Смертность потомства при насиживании (от начального числа яиц), % | 19.48 | 18.26 |
| В т. ч. эмбриональная смертность, % | 3.15 | 3.73 |
| Смертность гнездовых птенцов (от числа вылупившихся), % | 15.62 | 18.80 |
| в т. ч. частичный отход птенцов в успешных гнездах, % | 2.3 | 1.1 |
| Смертность гнездовых птенцов (от начального числа яиц), % | 11.50 | 14.11 |
| в т. ч. частичный отход птенцов в успешных гнездах, % | 1.7 | 0.8 |
| Репродуктивный успех (отношение числа слетков к числу отложенных яиц), % | 62.15 | 61.41 |
| Кол-во слетков на гнездо (x ± σ) | 3.6 ± 2.93 | 3.4 ± 2.74 |
| ошибка средней (m) | 0.30 | 0.51 |
| Кол-во слетков на успешное гнездо (x ± σ) | 5.8 ± 1.27 | 5.3 ± 1.15 |
| ошибка средней (m) | 0.17 | 0.22 |
| Вероятность гибели гнезда за сутки на стадиях кладки, насиживания, выкармливания гнездовых птенцов, % | 1.93; 1.24; 0.69 | 1.53; 0.87; 0.81 |
| Средняя высота (м) | 1.2 ± 1.13 | 1.3 ± 1.23 |
| ошибка средней (m) | 0.12 | 0,22 |
| лимиты | -0.12…4.5 | -0.35…5.5 |
| Соотношение легко-, средне-, труднодоступных и недоступных для человека гнезд, % | 90.9; 2.0; 7.1; 0.0 | 85.7; 8.2; 2.0; 4.1 |
Как для естественных, так и для антропогенных стаций характерна большая смертность кладок, чем гнездовых птенцов. В пригородных лесах отмечена также гибель потомства при исчезновении (вероятно, гибели) самок на всех стадиях репродуктивного цикла, преимущественно при кладке яиц. Это 8.5 % от всех приступивших к размножению самок. В предыдущих исследованиях показано, что именно на этой стадии самки оказываются повышенно уязвимыми перед хищниками (Зимин, 2009).
Отсутствие таких наблюдений в антропогенных стациях свидетельствует о меньшем давлении неспециализированных хищников на взрослых птиц. Вероятность гибели гнезда за сутки в период кладки составила в естественных стациях 1.93 %, при насиживании – 1.24 %, во время выкармливания гнездовых птенцов – 0.69 %. В антропогенных стациях соответственно 1.53, 0.87 и 0.81 %. Очевидно, что в обоих типах стаций проявляется сходная закономерность. По нашему мнению, она указывает на роль хищников в структуре репродуктивных потерь зарянки.
Снижение вероятности разорения гнезда по мере протекания репродуктивного цикла можно объяснить первоочередной гибелью наиболее заметных для хищников гнезд. Во время выкармливания птенцов вероятность разорения оказалась самой низкой, вероятно, потому, что наиболее заметные гнезда к этой стадии элиминировались уже в течение более чем 3 недель, а взрослые птицы у гнезд ведут себя крайне осторожно, что в большей степени характерно не для дуплогнездников, а для открытогнездящихся птиц.
Показатели репродуктивного успеха птиц существенно изменяются в зависимости от особенностей биотопа, условий конкретного года, антропогенной нагрузки и т. д. Изучение репродуктивных показателей зарянки в различных частях ареала показало, что для них характерно высокое разнообразие. По данным разных авторов, успех размножения составляет обычно 50–75 % (Зимин, 2009; Прокофьева, 2006; Сотников, 2008; Фуфаев, 1982).
Взаимосвязь различных гнездовых показателей, адаптивность антропогенных изменений гнездования
Исследование закономерностей расположения гнезд показывает, что некоторые типы гнездования распространены более, чем другие. Логично предположить, что именно они наиболее благоприятны для зарянки. Однако, поскольку стереотип гнездования (даже в случае его антропогенной трансформации) представляет собой относительно стабильное свойство вида, а репродуктивный успех в значительной мере является функцией изменчивой среды, полного их соответствия не может быть.
В табл. 2 показано, что некоторые репродуктивные показатели зарянок существенно отличаются в разных типах укрытий.
Таблица 2. Специфика репродуктивных показателей зарянок в естественных (А) и антропогенных (Б) стациях
(2000–2017 гг.)
| Тип расположения гнезда | Доля, % | Размер кладки (x ± σ) | Доля разоренных гнезд, % | Успех (отношение числа слетков к числу отложенных яиц), % | ||||
| А | Б | А | Б | А | Б | А | Б | |
| В выворотне | 1.1 | – | 7.0 | – | 0.0 | – | 57.0 | – |
| В искусственных гнездовьях | 2.1 | 2.0 | 6.0 | 6.0 | 0.0 | 100.0 | 62.5 | 0.0 |
| На деревьях открыто | 2.1 | – | 7.0 | – | 50.0 | – | 36.4 | – |
| В пнях | 5.3 | 2.0 | 5.0 ± 1.22 | 5.0 | 80.0 | 100.0 | 24.0 | 0.0 |
| На обрывчиках | 10.6 | – | 6.3 ± 0.71 | – | 30.0 | – | 52.6 | – |
| В дуплах и полудуплах | 63.8 | 24.5 | 6.2 ± 1.01 | 6.2 ± 0.83 | 28.3 | 25.0 | 68.2 | 74.6 |
| На земле | 14.9 | 8.2 | 6.4 ± 0.81 | 6.0 ± 0.00 | 42.9 | 25.0 | 58.5 | 66.7 |
| В антропогенных укрытиях на земле | – | 20.4 | – | 5.5 ± 1.38 | – | 30.0 | – | 60.0 |
| В антропогенных укрытиях выше уровня земли | – | 42.9 | – | 6.1 ± 1.21 | – | 28.6 | – | 59.8 |
По нашему мнению, в отношении наиболее редко используемых укрытий такие отличия могут оказаться случайными, поскольку по ним собрано небольшое количество данных именно вследствие редкого использования. Однако в отношении наиболее предпочитаемых птицами укрытий, по которым объем материала значительно больше, такие отличия также отмечены.
Обсуждение
Анализ специфики репродуктивных показателей зарянок в различных типах естественных и антропогенных укрытий показывает, что наиболее интересная связь наблюдается между типом расположения гнезда и величиной кладки. Существует тенденция более крупных кладок в наиболее часто используемых типах укрытий. Здесь же наблюдается и наименьшая вариабельность кладок. По нашему мнению, это свидетельствует как о наличии у популяции в целом определенных предпочтений, так и о том, что наиболее предпочитаемые укрытия чаще занимают физиологически более полноценные особи.
Данная закономерность прослеживается именно на уровне тенденции, поскольку корреляция (коэффициент Пирсона) показателей, хотя и положительная, оказалась недостоверной (для естественных стаций r = 0.35, p > 0.05, для антропогенных стаций r = 0.22, p > 0.05). Поскольку для изучаемого вида характерна территориальность, всегда имеется некоторый резерв гнездопригодных укрытий, и конкуренция между соседними парами за оптимальное место размещения гнезда не может быть большой.
Более заметная связь данных показателей в естественных стациях, по нашему мнению, также определяется более высокой численностью зарянок, тогда как в антропогенном ландшафте гнездовые участки отдельных пар часто не смыкаются.
Второй неожиданный факт – больший разброс кладок по количеству содержащихся в них яиц в антропогенных укрытиях, чем в естественных, в одних и тех же стациях (r = 0.69, p < 0.01, сравнивалась доля гнезд в каждом типе укрытия и средний для него размер кладки). По нашему мнению, это может свидетельствовать о разнокачественности особей селитебных стаций, когда часть (гнездящиеся в естественных укрытиях) не отличается от «диких» птиц, а другие представляют собой зарождающуюся синантропную популяцию.
Как известно, у синантропных птиц более выражена вариабельность и других аспектов биологии (Барановский, Иванов, 2014а; Ильенко, 1976). Косвенным подтверждением вышесказанного были результаты эксперимента, проведенного в 2016–2017 гг. После размещения в подходящих местах городских насаждений старых банок из-под краски (на земле) часть из них оказалась заселена зарянками (по 1–2 в год), тогда как в пригородных лесах при таком же эксперименте ни одна банка не была занята. Вероятно, в антропогенных стациях уже произошло устойчивое изменение стереотипа гнездования, что предполагает и определенную стабильность состава обитающих там особей.
Возникает вопрос – насколько гнездовые предпочтения зарянок адаптивны? Анализ данных позволил обнаружить тенденцию более редкого разорения гнезд, расположенных в предпочитаемых птицами местах, и, соответственно, более высокого репродуктивного успеха в таких гнездах. В естественных стациях эта связь в обоих случаях оказалась достоверной (r = -0.56, p < 0,05; r = -0.71, p < 0.01). В населенных пунктах она выражена на уровне тенденции, поскольку более сильной оказывается другая закономерность – большей успешности гнездования в дуплах деревьев. Смешение этих тенденций приводит к статистической недостоверности обеих связей (r = -0.45, p > 0.05; r = 0.34, p > 0.05), поскольку они оказываются направлены взаимно противоположным образом. Это свидетельствует о том, что использование антропогенных укрытий в городах может служить для зарянок своеобразной экологической ловушкой, поскольку демаскирует гнезда и снижает репродуктивный успех.
Заключение
Антропогенный, особенно селитебный, ландшафт в целом является неблагоприятным для зарянки, несмотря на то, что отдельные пары гнездятся даже в центральных частях больших городов. Индекс синантропизации этого вида составляет 21.55 (Барановский, Иванов, 2016). Однако у зарянки существует ряд антропогенных трансформаций всех основных аспектов биологии – не только гнездования, но и питания, кормового и оборонительного поведения, антропотолерантности (Барановский, Иванов, 2013), некоторые из которых, вероятно, адаптивны.
Мы предполагаем адаптивность в первую очередь некоторых аспектов поведения, таких как высокая антропотолерантность, трофические ассоциации с человеком и техникой (Барановский, Иванов, 2013). Изменения в питании зарянок, как и у других насекомоядных птиц, свидетельствуют о неблагоприятной для них трофической ситуации в населенных пунктах (Барановский, Иванов, 2013, 2014б).
В плане гнездового поведения главнейшей антропогенной трансформацией является переход на использование антропогенных укрытий, тем более выраженный, чем интенсивнее антропогенная трансформация среды.
Вероятно, отличия в выборе мест раздмещения гнезд отражают более глубокую разнокачественность городских особей, что предполагает существование особой синантропной популяции зарянок, более или менее устойчивой и обособленной от «диких» птиц.
Репродуктивный успех особей, гнездящихся в антропогенных укрытиях, несколько ниже, что позволяет рассматривать их переход на новый тип гнездования как одну из экологических ловушек антропогенного ландшафта.
Библиография
Барановский А. В., Иванов Е. С. Гнездящиеся птицы города Рязани (Атлас распространения и особенности биологии) . Рязань: ПервопечатникЪ, 2016. 367 с.
Барановский А. В., Иванов Е. С. Некоторые аспекты экологии зарянки в природных и антропогенных стациях Рязанской области // Вестник Балтийского федерального университета им. И. Канта. 2013. Вып. 7. Естественные науки. С. 86–94.
Барановский А. В., Иванов Е. С. Окрасочный полиморфизм и аберрации окраски оперения синантропных птиц в г. Рязани // Евразийский союз ученых. 2014а. № 9. С. 34–36.
Барановский А. В., Иванов Е. С. Специфика трофической стратегии насекомоядных птиц в естественных и антропогенных ландшафтах (на примере двух видов мухоловок) // Проблемы региональной экологии. 2014б. № 6. С. 87–91.
Благосклонов К. Н. Гнездование зарянки Erithacus rubecula в искусственных гнездовьях // Русский орнитологический журнал. 2017. Т. 26. № 1433. С. 1596–1597.
Болотников А. М., Хазиева С. М., Шураков А. И., Фуфаев А. А. Заметки о плодовитости и сроках гнездовой жизни некоторых птиц Предуралья // Сборник статей по птицеводству и орнитологии. Т. 69. Пермь, 1969. С. 67–72.
Головань В. И. Случаи нетипичного расположения гнезд воробьиных на юго-востоке Псковской области // Русский орнитологический журнал. Экспресс-выпуск № 23. 1997. С. 21.
Зимин В. Б. Зарянка на севере ареала. Т. 1. Распространение. Численность. Размножение . Петрозаводск: Карельский научный центр РАН, 2009. 443 с.
Ильенко А. И. Экология домовых воробьев и их эктопаразитов . М.: Наука, 1976. 120 с.
Ильичев В. Д., Бутьев В. Т., Константинов В. М. Птицы Москвы и Подмосковья . М.: Наука, 1987. 220 с.
Кладки и размеры яиц птиц юго-востока Мещерской низменности (Труды ОБГЗ. Вып. 18) / А. Д. Нумеров, С. Г. Приклонский, В. П. Иванчев, Ю. В. Котюков, Т. А. Кашенцева, Ю. М. Маркин, А. В. Постельных. М.: ЦНИЛ охотничьего хозяйства и заповедников, 1995. 168 с.
Лебедева Н. В., Ломадзе Н. Х. Зарянка Erithacus Rubecula на Северо-Западном Кавказе // Труды Южного научного центра Российской академии наук. Т. 3: Биоразнообразие и трансформация горных экосистем Кавказа. Ростов-на-Дону: ЮНЦ РАН, 2007. С. 252–277.
Нумеров А. Д., Климов А. С., Труфанова Е. И. Полевые исследования наземных позвоночных . Воронеж: ВГУ, 2010. 301 с.
Прокофьева И. В. О гнездовании зарянки Erithacus rubecula на юге Ленинградской области // Русский орнитологический журнал. Экспресс-выпуск 308. 2006. С. 100–105.
Птицы городов Среднего Поволжья и Предуралья / Казанский гос. пед. ун-т; Отв. ред. И. И. Рахимов. Казань: Мастер Лайн, 2001. 272 с.
Сапетина И. М. Птицы Окского заповедника и сопредельных территорий (биология, численность, охрана). Т. 2. Воробьиные птицы . М.: КМК, 2009. 172 с.
Симкин Г. Н. Певчие птицы . М.: Лесная промышленность, 1990. 400 с.
Сотников В. Н. Птицы Кировской области и сопредельных территорий. Т. 2. Воробьинообразные. Ч. 2 . Киров: ООО «Триада плюс», 2008. 432 с.
Фуфаев А. А. Величина кладки и успех размножения у воробьиных птиц Камского Предуралья // Гнездовая жизнь птиц. Пермь, 1982. С. 38–47.
Хазиева С. М., Болотников А. М., Каменский Ю. Н., Никольская В. И. Материалы о гнездящихся птицах правобережья реки Камы Добрянского района Пермской области // Гнездовая жизнь птиц. Пермь, 1975. С. 117–127.
Шепель А. И., Зиновьев Е. А. Животный мир заказника «Предуралье» (позвоночные) . Пермь: Изд-во Пермского ун-та, 1999. 144 с.
Saemann D. Die Brutvogelfauna einer sächsischen Grossstadt // Veröff. Mus. Naturkde. Karl-Marx-Stadt. 1970. № 5. Р. 21–85.
Saemann D. Untersuchungen zur Siedlungsdichte der Vögel in verschiedenen Grossstadthabitaten // Mitt. IG Avifauna DDR. 1973. № 6. Р. 3–24.

 © 2011 - 2026
© 2011 - 2026