



Введение
На большую роль различных форм активности (реактивности) организмов, их группировок и сообществ в организации, функционировании и развитии экосистем и биосферы указывают современные исследования (Васильев, 2005; Biosphere…, 2008; Чайковский, 2008; Иорданский, 2011; Grime, Pierce, 2012; Левченко, 2012; Проблемы…, 2013; Савинов, 2016б). В том числе показано, что растения как важнейшие продуценты характеризуются специфичными вещественными, энергетическими и информационными взаимодействиями с разнообразными абиотическими и биотическими факторами экосистем (Каратыгин, 1993; Photosynthesis…, 1996; Gurevitch et al., 2002; Тихонович, Проворов, 2009; Савинов, 2012). Это многообразие связей и взаимодействий отражается в размерах и географическом положении ареалов растений, обилии их групп, степени и формах внутривидовой дифференциации (Жизнь растений, 1974; Gurevitch et al., 2002). Одни группы растений, биологически прогрессируя, активно распространяются, увеличивают численность и усиливают адаптивную радиацию, представители же других групп, напротив, оказываются менее активными в освоении новых биотопов, поддержании численности, внутривидовой дифференциации, регрессируют, нередко оказываясь на грани исчезновения, или вымирают (Мелихова, 2014).
Важную роль таких градаций активности видов растений в формировании фитоценозов исследователи стали осознавать постепенно.
Так, еще в 30-х годах прошлого века Л. Г. Раменский (1971) отметил (тезисно), что виды растений разных семейств активно и конвергентно осуществляют в фитоценозах специализацию, приводящую к формированию трех ценотипов: 1) виоленты (силовики), являющиеся, как правило, эдификаторами в мезофильных биотопах, энергично развивающиеся и захватывающие территорию, подавляющие полнотой использования ресурсов среды другие виды растений; 2) патиенты (выносливцы), успех которых в борьбе за существование достигается не энергией жизнедеятельности, а выносливостью к неблагоприятным условиям биотопов (засолению, закислению почвы, аридности климата и т. п.); 3) эксплеренты (выполняющие), не обладающие качествами виолентов и патиентов, но способные очень быстро временно захватывать появляющиеся (пионерные) биотопы, еще не занятые виолентами или освобожденные от них естественным или искусственным путем. Л. Г. Раменский (1971) отметил, что предложенная им классификация ценотипов растений весьма условна, поскольку большинство растений совмещают признаки разных ценотипов, являются переходными формами или проявляют изменчивость метаболизма в разных биотопах. Впоследствии Т. А. Работнов (1974, с. 208) предложил расширить классификацию ценотипов Л. Г. Раменского и дополнил ее новыми группами (субвиолентами, субпатиентами, флюктуационными и сукцессионными эксплерентами).
Однако в дальнейшем ценотипическая концепция Л. Г. Раменского отечественными фитоценологами мало использовалась и не получила широкого освещения в отечественной и зарубежной литературе. Вследствие этого, независимо от Л. Г. Раменского, английским фитоценологом Дж. Граймом (Grime, 1974, 1977, 1979; Grime et al., 1988; Grime, Pierce, 2012) в 70-х годах прошлого века была предложена и развивается до сих пор теория адаптивных (экологических) стратегий растений, аналогичная ценотипической концепции Раменского, что отметил и сам Дж. Грайм (Grime, Pierce, 2012). Как и Л. Г. Раменский, Дж. Грайм (Grime, 1974) выделил те же самые функциональные группы растений (в соответствии с их адаптивными стратегиями), хотя неудачно (Работнов, 1985) назвал их: 1) конкуренты (С), 2) стресс-толеранты (S) и 3) рудералы (R). В связи с этим зарубежными фитоценологами и рядом отечественных ученых исследования адаптивных стратегий растений ведутся в рамках CSR-теории Грайма (Миркин и др., 1999; Пьянков, Иванов, 2000; Пьянков и др., 2001; Franzaring et al., 2007; Pierce et al., 2007, 2013, 2014, 2017; Kosakivska, 2008; Иванова, Юмагулова, 2009; Бетехтина, Веселкин, 2011; Магомедмирзаев и др., 2013; Сибгатуллина, Валеева, 2013; Магомедмирзаев, Гусейнова, 2015; Li, Shipley, 2017). При этом важно, что и выделяемые ценотические (ценобиотические) типы по Раменскому, и соответствующие им основные типы стратегии по Грайму характеризуют взаимоотношения между видами растений в соответствующих биотопах (Работнов, 1985), и этот подход в фитоценологии можно условно назвать ценотипическим.
В 60-х годах прошлого века Б. А. Юрцевым (1966, 1968) была сформулирована концепция об активности видов растений как мере их преуспевания в соответствующих ландшафтно-климатических условиях, как мере их фитоценотической значимости. Подход, сформировавшийся на основе указанной концепции, можно условно назвать статусным (от лат. status – положение, состояние). Статусный подход обусловил спектр исследований, при выполнении которых с помощью ряда сходных методов был получен массив данных по активности видов растений в различных наземных и водных экосистемах (Дидух, 1982; Свириденко, 2000; Налимова, 2006; Чепинога, Росбах, 2008; Бабич, Ушакова, 2012; Коновалова и др., 2014; Панкова, 2014). По мнению исследователей, эти данные позволили глубже отразить структуру фитоценозов, оценить их устойчивость к воздействию негативных природных и антропогенных экологических факторов, а также прогнозировать динамику структурно-функциональных параметров сообществ (Бабич, Ушакова, 2012; Коновалова и др., 2014). Параллельно проводятся исследования ценотической активности кормовых трав в агроэкосистемах (Рыженко, 2012; Минвалиев, Павлова, 2015), развиваются представления о неоднородности (квантованности) растительного покрова (Ипатов, 2016; Лебедева и др., 2016), не связанные со статусным подходом, но близкие к нему.
Иной подход, который условно можно обозначить как эргонтический (от греч. ergon – действие, работа), предложен и развивается нами (Савинов, 2015а, б, 2016а, б; Савинов, Никитин, 2016). Он основан на определении активности прежде всего как интенсивности (скорости) взаимодействия биологической системы с другими внешними биотическими и абиотическими системами в вещественном, энергетическом и информационном аспектах. Формализация этого определения позволила разработать и начать апробировать метод количественной оценки активности групп растений и животных на популяционном уровне (Савинов, 2015а, 2016а, б; Савинов, Никитин, 2016). Представляется, что предложенный метод позволяет вести в новом аспекте теоретические и прикладные исследования, в частности в плане биоиндикации и биомониторинга экосистем (Савинов, Никитин, 2016), а также в области эволюционной экологии (Савинов, 2015б, 2016а, б).
Целью данной статьи является сравнительный анализ ценотипического, статусного и эргонтического подходов к оценке активности растений как особого экологического фактора в фитоценозах и экосистемах.
Аналитический обзор
Теория экологических стратегий растений Раменского – Грайма (ценотипический подход)
Выделенные Л. Г. Раменским (1971) ценотипы (виоленты – силовики, патиенты – выносливцы, эксплеренты – выполняющие) «по существу характеризуют три основных направления приспособления растений, возникших в процессе их сопряженной эволюции при совместном произрастании и в процессе формирования фитоценозов (фитоценогенеза)» (Работнов, 1985, с. 3). При этом Л. Г. Раменский (1971, с. 232) специально отметил, что выделенные им основные ценобиологические типы – результат схематизации, поскольку, во-первых, «в действительности большинство растений имеет характеристику переходную или смешанную, совмещающую черты двух типов», во-вторых, ценотипы видов растений могут меняться в различных биотопах. Поэтому «в большинстве растительных группировок имеются виды, приближающиеся по своим свойствам к трем намеченным типам: силовиков, выносливцев и выполняющих; их соотношение отражает условия среды и внутренний ценобиотический строй группировки» (Раменский, 1971, с. 232).
К аналогичным выводам пришел и Дж. Грайм (Grime, 1974, 1977, 1979; Grime et al., 1988) и потому, кроме выделенных им типов конкурентов (С), стресс-толерантов (S) и рудералов (R), дополнительно указал возникающие в зависимости от биотопических условий промежуточные типы (CR, SC, RS, CSR), неудачно назвав их типами вторичной стратегии (Работнов, 1985). В действительности же, как отметил Т. А. Работнов (1985), во-первых, следует говорить о патиентности и виолентности как естественных качествах растений, поскольку, например, патиентность является универсальным свойством растений, в частности, виоленты в градиенте условий произрастания нередко переходят в патиентное состояние, также патиентами они являются в составе сомкнутых фитоценозов в начальный период онтогенеза и иногда в фазе старения. Во-вторых, по мнению Т. А. Работнова (1985, с. 8), «эксплеренты целесообразно трактовать несколько шире, чем это принято Л. Г. Раменским и Граймом, включив в этот тип стратегии растения, способные быстро и положительно реагировать на ослабление конкуренции не только в результате различных форм нарушения растительности». Тогда эксплерентами являются и весенние эфемероиды некоторых типов листопадных лесов в период ослабления конкуренции со стороны деревьев-виолентов (Работнов, 1985). В-третьих, Т. А. Работнов (1985) предположил, что помимо трех основных типов стратегии по Раменскому – Грайму следует выделить четвертый тип пионерных растений, свойственных первым этапам первичных сукцессий, а также пятый тип, отмеченный Р. Уиттекером (1980) в отношении некоторых растений (пустынных однолетников, однолетних луговых и степных вик), развивающихся лишь эпизодически, при наступлении благоприятных условий для них (при прорастании их семян, годами покоящихся в почве, после ее обильного увлажнения атмосферными осадками).
Т. А. Работнов (1974, с. 208) указал, что «в понятие ценотипов следует вкладывать динамическое содержание», поскольку они являются группами видов растений «со сходным изменением их ценотической значимости в зависимости от условий произрастания или особенностей их жизненного цикла». Исходя из этого, Т. А. Работнов (1974, с. 208) предложил расширенную классификацию ценотипов, выделив наряду с виолентами, патиентами и эксплерентами дополнительно новые группы: субвиоленты – «многолетние растения, способные в благоприятных условиях создавать монодоминантные сообщества, но недостаточно конкурентоспособные»; субпатиенты – виды, сходные с патиентами в отношении биотопов, способные создавать монодоминантные сообщества, но иногда лишь на короткий срок; флюктуационные эксплеренты – многолетние (реже – малолетние) растения, принимающие малое участие в формировании фитоценозов, но способные быстро и резко положительно реагировать на ослабление конкуренции и недолго доминировать при массовом отмирании или ослаблении доминантов; сукцессионные эксплеренты – растения, длительно доминирующие при непрерывном воздействии фактора, вызвавшего нарушение прежнего фитоценоза.
В процессе непрерывного взаимодействия видов растений с биотическими и абиотическими факторами внешней среды происходит фитоценогенез – коадаптация данных видов, в результате чего исторически формируются фитоценозы (Работнов, 1985). В соответствии с этим под «стратегией вида» Т. А. Работнов (1975, 1985) предложил понимать совокупность приспособлений, обеспечивающих виду возможность совместно обитать с другими организмами и занимать определенное положение в биоценозе. Все это позволяет говорить о взаимосвязи между представлениями о ценотипах Раменского – Работнова (Раменский, 1971; Работнов, 1975, 1985) и адаптивными стратегиями Раменского – Грайма (Grime et al., 1988; Grime, Pierce, 2012), статусным (Юрцев, 1966, 1968) и эргонтическим (Савинов, 2015а, 2016а, б, 2017) подходами к оценке активности растений в экосистемах.
В теории экологических стратегий не используется понятие «активность растений», но при расчетах по системе Раменского – Грайма (CSR-теории) обычно применяется комплекс показателей, характеризующих энергию жизнедеятельности видов растений разных семейств, степень и скорость освоения ими ресурсов экосистемной среды в сравнении с конкурентами (Grime, 1974, 1977, 1979; Grime et al., 1988; Grime, Pierce, 2012; Работнов, 1985; Миркин и др., 1999; Пьянков, Иванов, 2000; Пьянков и др., 2001; Усманов и др., 2001; Franzaring et al., 2007; Pierce et al., 2007, 2013, 2014, 2017; Kosakivska, 2008; Иванова, Юмагулова, 2009; Магомедмирзаев и др., 2013; Сибгатуллина, Валеева, 2013; Магомедмирзаев, Гусейнова, 2015; Li, Shipley, 2017); наряду с этим предпринимаются попытки связать типы экологических стратегий со способностью растений к микоризообразованию и его интенсивностью (Grime et al., 1988; Cornelissen et al., 2001; Бетехтина, Веселкин, 2011).
Дж. Грайм (Grime, 1979; Grime et al., 1988) при выделении типов экологических стратегий использовал известный продукционный показатель – относительную скорость роста растений (RGR) и морфологический индекс (М):
RGR = (lnW2 – lnW1) / (t2 – t1), (4)
М = (a + b + c) / 2, (5)
где RGR – относительная скорость роста (relative growth rate) (г/г . сут), W2, W1 – сухая масса растения (г), А1, A2 – площадь листьев (см2) в моменты времени t1 и t2 соответственно; a – максимальная высота растения, b – величина горизонтального разрастания растения и c – максимальная величина опада (в баллах).
Впоследствии для установления адаптивных стратегий стали использовать отношение суммарной площади листьев к массе растения – LAR (leaf area ratio) (cм2/г), скорость чистой ассимиляции единицы листовой поверхности – NAR (net assimilation rate) (г/см2 . сут), а также составляющие LAR (Пьянков, Иванов, 2000; Craine, 2005, 2007; Carlyle, Fraser, 2006; Franzaring et al., 2007; Иванова, Юмагулова, 2009; Cerabolini et al., 2010; Pierce et al., 2013, 2014, 2017):
RGR = NAR × LAR, (6)
LAR = SLA × LMR, (7)
где SLA – площадь единицы массы листа (мм2/мг), LMR – листовой индекс (отношение биомассы листьев к общей биомассе растения, мг/мг) (Angert et al., 2007).
Однако указанные функциональные показатели не всегда однозначно указывают на принадлежность данного вида растений к группе с той или иной экологической стратегией. Например, у 35 видов дикорастущих растений разных стратегий (C, S, R) в условиях бореальной зоны Среднего Урала не было обнаружено значимых различий по LAR и SLA, а R-стратеги не имели существенных отличий по LMR от C- и S-стратегов (Пьянков, Иванов, 2000). Показано также, что в сообществе верхового болота в условиях западно-сибирской тайги разнонаправленно изменялись функциональные параметры R-, C- и S-стратегов: величины интенсивности транспирации листьев увеличивались в ряду R→C→S, интенсивности фотосинтеза – в ряду S→C→R, интенсивности дыхания – в ряду C→R→S (Иванова, Юмагулова, 2009).
Несомненно, у того или иного вида растения тип основной экологической стратегии генетически запрограммирован, хотя и может меняться в онтогенезе; при этом функциональные и морфологические параметры колеблются в пределах нормы реакции, отражая виолентность, патиентность и эксплерентность, проявляющиеся в соответствующих экологических условиях у всех видов растений, но в разной степени (Работнов, 1985; Пьянков и др., 2001; Усманов и др., 2001). Показано, что тип экологической стратегии связан с направленностью метаболических процессов у растений, особенностями их химического состава (Пьянков, Иванов, 2000; Пьянков и др., 2001), степенью микоризации (Бетехтина, Веселкин, 2011). Имеющиеся данные (Работнов, 1985; Pierce et al., 2017) позволяют полагать, что тип определенной экологической стратегии конвергентно сформировался в ходе адаптациогенеза у растений разных семейств в пределах соответствующих природно-климатических зон, экосистем и биомов. Несомненно, благодаря разным типам экологических стратегий (C, S, R, CR, SC, RS, CSR) наземных и водных растений формируются адекватные биотопам биоценозы, происходят сукцессии экосистем.
Можно согласиться с мнениями о том, что хотя теория Раменского – Грайма подвергается критике (Craine, 2005, 2007; Grime, 2007; Tilman, 2007), предельно обобщенно характеризует направления адаптациогенеза растений и способы этой адаптации, но указанная теория открыта для дополнения новыми положениями и данными, вследствие чего обладает операциональной и прогностической ценностью (Работнов, 1985; Миркин и др., 1999; Бетехтина, Веселкин, 2011). В отличие от статусного (Юрцев, 1966, 1968) и эргонтического подходов (Савинов, 2015а, б, 2016а, б, 2017), теория экологических стратегий Раменского – Грайма не оперирует понятием «активность» и не ставит задачи количественного определения активности растений, но вопросы адаптации, разрабатываемые в теории экологических стратегий, напрямую связаны с аспектами активности ценопопуляций и видов.
Статусный подход к оценке активности растений в экосистемах
Основоположник статусного подхода Б. А. Юрцев (1966, с. 5) предложил активными видами растений считать «преуспевающий элемент флоры, противостоящий реликтовым ее элементам»; при этом Б. А. Юрцев полагал, что «эколого-биологические свойства активных видов соответствуют общей ландшафтно-климатической обстановке данной территории, и это находит свое выражение в повышенной численности таких видов, значительной широте их экологической амплитуды, более равномерном распределении их по территории, – то есть в более интенсивном освоении этими видами данного ландшафта». Впоследствии Б. А. Юрцев (1968) увеличил число критериев активных видов и конкретизировал высказанные положения. В частности, исследуя фитоценозы высокогорий северо-востока Сибири (Юрцев, 1968), он учитывал, что активность вида в пределах ландшафта тем выше, во-первых, чем больше экологическая амплитуда вида (т. е. чем больше разнообразие заселенных им экотопов); во-вторых, чем равномернее вид распределен по территории и, в-третьих, чем выше общий уровень численности вида в занимаемых им экотопах. На основании соотношений этих показателей Б. А. Юрцев в фитоценозах выделял 5 ступеней активности видов растений: 1) особо активные (I ступень активности), 2) высокоактивные (II ступень), 3) среднеактивные (III ступень), 4) малоактивные (IV ступень), 5) неактивные (V ступень). Но поскольку у видов одной и той же ступени активности соотношение широты экологической амплитуды, равномерности распределения по территории и уровня численности может быть неодинаковым, то виды каждой ступени, в свою очередь, были подразделены на подгруппы (табл. 1). Наряду с этими критериями Б. А. Юрцев (1968) предлагал по возможности определять дополнительные важные характеристики активности вида: его жизненность в основных экотопах, фенотипический (в том числе возрастной) и генотипический составы ценопопуляций; но полагал, что эти характеристики можно оценить только при длительных стационарных исследованиях.
В отношении растений наземных экосистем в настоящее время используются различные модификации метода Б. А. Юрцева в форме степенных (балльных) измерений внутриландшафтной (парциальной) активности видов (Налимова, 2003, 2006; Чепинога, Росбах, 2008; Бабич, Ушакова, 2012; Коновалова и др., 2014). Одна из модификаций метода представлена в табл. 2.
Таблица 1. Критерии выделения ступеней (степеней) активности видов растений (по Юрцеву, 1968)
| Группы видов по характерному для них уровню численности | Виды | |||||||||
| стенотопные | ||||||||||
| ЭТ | ГЭТ | ГСТ | обычных экотопов | редких экотопов | ||||||
| ПМ | СП | ПМ | СП | ПМ | СП | П | М | П | М | |
| Массовые | Iа | Iб | IIа | IIб | IIIа | IVа | IIIб | IVб | IVв | Vа |
| Малочисленные | IIв | IIг | IIIв | IIIг | IIIд | IVг | IIIе | Vб | Vв | Vг |
| Единично встречающиеся | IIIж | IVд | IVе | Vд | IVж | Vе | IVз | Vж | Vз | Vи |
Примечание. ЭТ – эвритопные, ГЭТ – гемиэвритопные, ГСТ – гемистенотопные, ПМ – повсеместные, СП – спорадические, П – постоянные, М – малопостоянные, I – особо активные, II – высокоактивные, III – среднеактивные, IV – малоактивные, V – неактивные.
Таблица 2. Количественные критерии степеней активности видов растений (по Налимовой, 2003, 2006, с коррекциями)
| Степени активности | Показатели активности, в долях | ||
| амплитуда экологического пространства | встречаемость | обилие | |
| Высокая (I) | >0.4 | >0.4 | 0.3–1.0 |
| Средняя (II) | 0.2–0.4 | 0.2–0.4 | 0.04 |
| Низкая (III) | 0.1–0.2 | 0.1–0.2 | 0.03 |
| Очень низкая (IV) | <0.1 | <0.1 | 0.01 |
С помощью этой и аналогичных методик (Бабич, Ушакова, 2012; Коновалова и др., 2014) были получены данные, показывающие сходство в соотношении долей активных и неактивных видов растений для фитоценозов разных типов наземных и водных экосистем. В частности, в луговых фитоценозах заповедника «Присурский» (Чувашская Республика) 68.2–97.1 % видов растений характеризовались низкой активностью, из них до 60 % видов – очень низкой активностью; средней активностью обладали 2.9–31.8 % видов и только 6 % – высокой активностью (Налимова, 2006). В лесных питомниках южнотаежной зоны Вологодской области сорные растения также представлены преимущественно неактивными стенотопными (56.8 %) и малоактивными гемиэвритопными, гемистенотопными и стенотопными (30.9 %) видами, а среднеактивные, высокоактивные и особоактивные виды составляют 5.6, 4.2 и 2.5 % соответственно (Бабич, Ушакова, 2012). При этом видами, проявляющими наибольшую ценотическую активность на полях питомников, являются типичные убиквисты: Achillea millefolium L., Chenopodium album L., Elytrigia repens L., Rumex acetosella L., Taraxacum officinale Wigg. Сходная ситуация отмечена и в фитоценозах лесных питомников Архангельской области (Коновалова и др., 2014). При этом соотношение групп видов с разной активностью рассматривается как отражение структуры фитоценозов.
Развивая идеи Б. А. Юрцева (1966, 1968), Я. П. Дидух (1982) предложил рассматривать активность вида в нескольких аспектах: 1) региональном, 2) временном, 3) фитоценотическом и 4) локальном. Этот подход был апробирован при изучении фитоценозов Ялтинского заповедника (Дидух, 1982). В частности, при изучении активности видов в первом аспекте выявлялись видовые оптимумы и тенденции (прогнозы) изменения видовых ареалов. При этом были выделены категории экспансивных, преуспевающих, угасающих и реликтовых видов растений. Изучение активности видов во временном аспекте позволило определить период оптимума (максимума, минимума) активности, изменений ее в различные исторические периоды, что помогает изучению активности в первом, региональном аспекте. Исследованиями активности видов в фитоценотическом аспекте выявлялась, во-первых, приуроченность оптимума активности к определенному синтаксону (или флороценотическому комплексу) и, во-вторых, изменения активности в других синтаксонах (или флороценотических комплексах) данного региона. Изучение активности видов в локальном аспекте позволило определить степень активности видов на небольших (меньше площади элементарной флоры) территориях, включающих только те ценозы, в которых встречался данный вид.
Наряду с расширением спектра показателей активности видов растений на базе методологии Б. А. Юрцева предлагались и упрощенные методики. Например, Л. И. Малышев (1973) считал, что для оценки активности (А) вида растения достаточно интегрального выражения лишь его встречаемости (F) и обилия (D):
A = (F · D)1/2 (1)
Б. Ф. Свириденко (2000) парциальную активность (ПА) в отношении видов гидромакрофитов также по существу рассматривает как функцию двух переменных величин – встречаемости и обилия в пределах одного класса экотопов и определяет активность так:
ПА = (ПП / 100 %) · (В / 100 %), (2)
где ПП – среднее проективное покрытие вида в популяциях (в %), В – экотопическая встречаемость популяций (%). При этом максимальная величина парциальной активности может быть равна 1 (при ПП = 100 %, В = 100 %), но ее реальные значения всегда ниже.
По предложению Б. Ф. Свириденко (2000), виды растений, у которых ПА < 0.01, следует считать неактивными, с величинами ПА > 0.01 – активными с подразделением на высокоактивные (ПА > 0.10), среднеактивные (0.05 < ПА < 0.10) и низкоактивные (0.01 < ПА < 0.05) виды.
В соответствии с этой методикой, например, в водоемах Северного Казахстана высокоактивными (ПА > 0.10) являются виды-убиквисты: Phragmites australis (Cav.) Trin. ex Steud. (ПА = 0.46–0.54), Potamogeton pectinatus L. (ПА = 0.18–0.35), Typha angustifolia L. (ПА = 0.15–0.20); низкоактивными (0.01 < ПА < 0.05) – Potamogeton pusillus L. (ПА = 0.01–0.04), Eleocharis palustris (L.) R. Br (ПА = 0.01–0.04); неактивными (ПА < 0.01) – Chara kirghisorum Lessing emend. Hollerbach (вид редкий и типичный олиготрофный, олигосапробный представитель), Elodea сanadensis Michx. (инвазионный вид, но еще относительно слабо распространившийся по водоемам Казахстана); при этом доля активных видов в экосистемах невелика – 18–28 % (Свириденко, 2000; Свириденко и др., 2013). Аналогичное распределение активных и неактивных видов гидромакрофитов наблюдается и в водоемах европейской части России: здесь во флоре доля активных видов составляет всего 4–7 %, а неактивных – 56–66% (Панкова, 2014), хотя в группу активных входят другие виды растений: Carex acuta L., Nuphar lutea (L.) Smith, Potamogeton natans L., Rorippa amphibia (L.) Bess., Sagittaria sagittifolia L., Sparganium erectum L. (Панкова, 2014). Активные виды как убиквисты нередко создают до 95 % общей биомассы макрофитов в лимносистемах (Грищенкова, Власов, 2008).
Проводятся методологически сходные исследования прикладного характера (но не связанные с теорией Б. А. Юрцева) в отношении ценотической активности кормовых трав в агроэкосистемах (Рыженко, 2012; Минвалиев, Павлова, 2015). В частности, изучаются конкурентные взаимоотношения (ценотическая активность) злаковых и бобовых трав в составе различных травосмесей и при разных уровнях минерального питания (с целью разработки технологий повышения урожайности травосмесей и качества их биомассы). При этом установлено, что ценотическая активность изучаемых видов многолетних злаковых и бобовых трав (доля каждого вида в составе агрофитоценозов) нестабильна и зависит от влияния многих экологических факторов: 1) фона минерального питания, 2) стадии вегетации, 3) видового состава травосмесей, 4) климатических условий года, 5) долголетия, темпа развития и отавности компонентов травосмеси (Минвалиев, Павлова, 2015).
Ко всем рассмотренным работам (с термином «активность») примыкают исследования неоднородности растительного покрова (Ипатов, 2016; Лебедева и др., 2016), не основанные на концепции статусного подхода, но методологически близкие к нему. В данном случае изучается многолетняя динамика видовой неоднородности (квантованности) фитоценозов с использованием коэффициента участия вида (КУ). Этот коэффициент определяется либо по массе вида в процентах от общей массы растений на учетной площади, либо по доле проективного покрытия вида (ППвида) в общем проективном покрытии видов (ППвидов) фитоценоза:
КУ = (ППвида / ∑ ППвидов ) . 100 %. (3)
Исследования с вычислением коэффициента участия (КУ) показали, что в одних фитоценозах сохраняется многолетнее доминирование некоторых видов растений на фоне большинства слабо представленных видов; в других же сообществах в отдельные годы или под влиянием микрорельефной неоднородности виды-доминанты значительно снижают КУ (Ипатов, 2016; Лебедева и др., 2016).
Эргонтический подход к оценке активности растений в экосистемах
Этот подход основан на предложенном нами определении: активность материальной системы, в том числе живой, есть характер и интенсивность ее взаимодействия с окружающей средой (т. е. с другими системами) в единицу времени, и чем сильнее и разнообразнее взаимодействует система с окружающей средой (другими системами), тем она активнее (Савинов, 2015а, 2016а, б). В отношении адаптирующихся, живых систем, это взаимодействие, т. е. активность (А), по существу отражает скорость аккумуляции или генерирования вещества, энергии и информации адаптирующейся системой, например биосистемой организменного, популяционного или биоценотического уровней (Савинов, 2015а):
А = М / t (8), А = Q / t (9), А = Н / t (10),
где М – масса потребленного и/или синтезированного вещества (г), Q – количество потребленной и/или выделенной энергии (Дж), Н – количество информации (бит), экзогенно полученное или эндогенно созданное биосистемой (ее структурно-функциональным матриксом) за единицу времени t (например, в секунду).
В физиологическом и экологическом аспектах жизнедеятельность растений традиционно характеризуется скоростью аккумуляции или продукции вещества или энергии (Angert et al., 2007; Susiluoto, Berninger, 2007). Таковыми «скоростными» показателями являются, например, 1) интенсивность фотосинтеза, выражаемая в мг поглощенного СО2, отнесенного к единице площади (дм2) листьев растения в единицу времени (мин, час); 2) продуктивность растений, определяемая количеством произведенного ими вещества (энергии) на единицу площади листьев в единицу времени (г/м2 . сутки, ккал/м2 . сутки). Очевидно, эти и подобные показатели характеризуют активность растений, однако в силу специфики они не будут сопоставимы с характеристиками активности других членов биоценоза – консументов и редуцентов, что затруднит исследования экосистем в новом аспекте. Если же попытаться выражать активность, в частности, как скорость информационного взаимодействия биосистем с окружающей средой, то указанных выше ограничений не возникает, и можно использовать универсальные единицы информации (бит, байт).
В предложенной формуле (10) величина Н определяется известной формулой Шеннона, ей принято выражать количество информации, накопленное или генерируемое биосистемами организменного и надорганизменного уровней (Емельянов, 1994; Алимов и др., 2013):
H = – ∑pi log2 pi. (11).
В формуле (11) pi есть частота какого-либо события; например, для исследований активности растений мы использовали: а) частоту проявления (доли) в ценопопуляции дискретных фенотипических признаков – фенов, б) доли (частоты) химических элементов, аккумулированных в растениях разных видов, в) частоты посещения насекомыми-опылителями видов энтомофильных растений. При перечисленных формализациях для растительных организмов будут определены разные формы (составляющие) активности: условно 1) морфогенетической активности (при использовании частот параметров фенотипической изменчивости), 2) аккумулятивной активности (например, на основе соотношения величин долей аккумуляции разных металлов), 3) консортивной (в частности, энтомофильной активности, при учете частот взаимодействия видов растений с видами антофильных насекомых) и т. п. При возможности определения нескольких подобных форм активности для одного и того же вида растения эти формы можно интегрировать в показатель обобщенной активности (Аd) по формуле (Савинов, 2017):
Ad = (A1 · A2 · … · An)1/n (12)
Очевидно, приоритет в определении той или иной формы активности будет обусловлен, например, задачами, объектами экологического исследования и возможностями исследователя.
Важно отметить, что, с одной стороны, известна взаимосвязь энергетических и массовых характеристик биосистем организменного и популяционного уровней (Большаков и др., 1998). С другой стороны, потоки энергии через указанные системы взаимосвязаны с потоками информации через них (Алимов и др., 2013). Количество этой информации является мерой организованности систем (Винер, 1958; Камшилов, 1970; Емельянов, 1994): выше организованные системы содержат больше информации, и наоборот. В свою очередь количество заключенной в системе информации адекватно разнообразию (неоднородности) элементов системы (Камшилов, 1970). Информационно значимые исследования фенотипического разнообразия на популяционном уровне актуальны и перспективны (Емельянов, 1994; Васильев, 2005). Учитывая это, мы в своих работах (Савинов, 2016а,б; Савинов, Никитин, 2016) использовали формулу (10), выражающую скорость информационного взаимодействия биосистемы популяционного уровня со средой обитания (Савинов, 2015а, 2016а, б).
Примеры использования эргонтического подхода
Апробирование эргонтического подхода проведено нами с использованием формулы (10) на основе собственных и литературных данных.
Определение морфогенетической активности растений
Были использованы многолетние данные о частотах встречаемости (долях) фенов листьев в ценопопуляциях сныти Aegopodium podagraria L. и подорожника Plantago major L. (Савинов и др., 2015; Савинов, Никитин, 2016) из городских парковых зон, служащих полигонами для наших регулярных биоиндикационных исследований (Савинов и др., 2007; Савинов и др., 2015) (рис.). Указанные ценопопуляции произрастали в 4 биотопах с разными уровнями токсической нагрузки (Безель и др., 1998) на почвы, загрязняемые тяжелыми металлами (ТМ): свинцом, цинком, медью и хромом. В частности, для биотопов № 1–4 (табл. 3) токсическая нагрузка составляла 3.42; 2.16; 1.94; 1.00 (условный контроль) относительных единиц соответственно. Время от начала вегетации растений до сбора листьев для выявления их фенов (рисунок) составляло 25–40 суток. По частотам встречаемости фенов листьев в ценопопуляциях сныти и подорожника (Савинов и др., 2015; Савинов, Никитин, 2016) нами рассчитаны величины Н ценопопуляций этих растений по формуле (11) для каждого сезона исследований. По величинам Н и учтенного времени вегетации с помощью формулы (10) были определены ежегодные величины морфогенетической активности растений и средняя величина этой активности за 5 сезонов исследований (Савинов, Никитин, 2016).
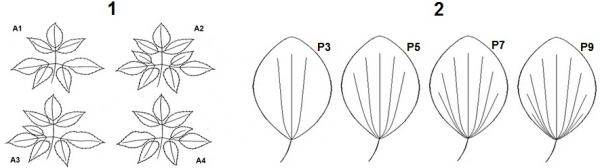
Рис. Фены листьев сныти (1) и подорожника (2) (Савинов и др., 2015; Савинов, Никитин, 2016)
Fig. Phens of leaves of goutweed (1) and plantain (2) (Savinov et al., 2015; Savinov, Nikitin, 2016)
Таблица 3. Динамика морфогенетической активности (10-7 бит/сек) ценопопуляций сныти и подорожника в биотопах по градиенту загрязнения ТМ (Савинов, Никитин, 2016)
| Ценопопуляции/ | Годы | Средняя активность | ||||
| биотопы | 2011 | 2012 | 2013 | 2014 | 2015 | в течение 5 сезонов |
| 1 | 5.13 | 2.15 | 5.9 | 4.91 | 5.13 | 4.64±0.58** |
| 4.05 | 4.42 | 3.6 | 4.17 | 3.19 | 3.89±0.49** | |
| 2 | 6.26 | 2.23 | 6.71 | 4.85 | 5.41 | 5.09±0.70** |
| 4.33 | 4.74 | 4.17 | 4.72 | 3.07 | 4.21±0.68** | |
| 3 | 6.9 | 2.87 | 7.11 | 5.68 | 5.97 | 5.71±0.68* |
| 5.32 | 5.41 | 4.91 | 5.53 | 3.84 | 5.00±0.69* | |
| 4 (контроль) | 7.73 | 4.92 | 7.89 | 6.03 | 6.98 | 6.71±0.50 |
| 6.98 | 6.19 | 5.32 | 5.85 | 4.18 | 5.70±0.42 | |
Примечание. Над чертой – величины активности ценопопуляций сныти, под чертой – то же для подорожника (пояснения в тексте); * – p < 0.05; ** – p < 0.01.
Расчеты показали, что морфогенетическая активность сныти и подорожника в их ценопопуляциях уменьшается с повышением токсической нагрузки ТМ на почвы, и эти различия сохраняются в течение нескольких лет (см. табл. 3). Эти результаты согласуются с данными о том, что наличие в почве комплекса ТМ (Pb, Zn, Cu, Cd) в дозах 1–5 ПДК ухудшает биометрические параметры растений, площадь фотосинтезирующей поверхности и может многократно снижать первичную продукцию (Щелчкова и др., 2010). С усилением загрязнения почв ТМ тормозятся морфогенетические процессы и снижается жизнеспособность аборигенных видов растений (Воробейчик, Хантемирова, 1994; Дуля и др., 2013).
Определение аккумулятивной активности растений
В отечественной и зарубежной литературе имеются многочисленные данные о количестве (концентрациях) химических элементов, в частности металлов, накопленных различными видами растений за определенные периоды вегетации. Аккумуляционную активность видов мы определяли (Савинов, 2016а, б) на основе таких литературных данных (Kozanecka et al., 2002; Кулагина, 2009; Шубина, Юрьев, 2009; Копылова, 2011; Великанова и др., 2012) о концентрациях металлов (Cu, Zn, Mn, Fe, Ni, Cr) в листьях споровых (Pleurozium schreberi (Willd. ex Brid.) Mitt., Lycopodium clavatum L., Dryopteris filix-mas (L.) Schott, Pteridium aquilinum (L.) Kuhn), покрытосеменных растений (Convallaria majalis L., Vaccinium myrtillus L., Polygonum aviculаre L., Plantаgo mаjor L., Populus balsamifera L., Ulmus pumila L., Malus baccata (L.) Borkh.) и хвое Pinus sylvestris L., сформировавшихся за время их вегетации (до сбора растительного материала). По концентрациям металлов в листьях мы рассчитывали доли каждого элемента в их пуле для каждого вида растений. Данные о долях металлов позволили рассчитать величины Н по формуле (11) для указанных видов растений. По величине Н и времени, за которое были накоплены металлы, по формуле (10) определены величины аккумулятивной активности растений.
Наши расчеты показали, что в экосистемах минимальную аккумуляционную активность в отношении металлов проявляют вечнозеленые растения (Pinus sylvestris, Vaccinium myrtillus) – 0.08–0.11 . 10-7 бит/сек, наибольшую – покрытосеменные с непродолжительным периодом вегетации (Convallaria majalis, Polygonum aviculаre, Plantаgo mаjor, Populus balsamifera, Ulmus pumila, Malus baccata) – 0.41–0.81 . 10-7 бит/сек, а споровые (Pleurozium schreberi, Lycopodium clavatum, Dryopteris filix-mas, Pteridium aquilinum) занимают промежуточное положение – 0.28–0.61 . 10-7 бит/сек (Савинов, 2016а, б).
Полученные данные согласуются с мнением Б. А. Юрцева (1966) о том, что вечнозеленость является архаичным признаком, эволюционным приспособлением, возникшим у предков вечнозеленых растений в связи с историческим развитием их в экосистемах с пониженными летними температурами и бедными почвами (с дефицитом элементов питания). Вечнозеленые растения (в отличие от летнезеленых) обладают пониженной интенсивностью многих физиологических процессов, малым приростом и длительностью функционирования осевых органов; поэтому развитие вечнозеленых растений в условиях дефицита элементов питания (нитратов, фосфатов, микроэлементов) происходит лишь благодаря активному симбиозу с микоризными грибами, обеспечивающими растения необходимыми минеральными соединениями (Юрцев, 1966; Suzuki et al., 2001; Smith, Read, 2008).
Определение энтомофильной активности растений
Известно, что взаимодействие цветковых растений и антофильных насекомых обусловливает их коэволюцию, при этом для насекомых привлекательность цветков растений обусловлена многими их признаками: цветом, запахом, количеством нектара, морфологическими особенностями и др. (Гринфельд, 1978).
Для оценки энтомофильной активности нами были использованы данные многочасовых учетов посещения антофильными насекомыми цветковых растений разных семейств в условиях Южного Зауралья (Сорокина, 2003). По долям (частотам встречаемости) особей каждого вида насекомых в учетах на 4 видах растений нами рассчитаны величины Н для этих растений по формуле (11). По величинам Н и времени пребывания насекомых на растениях с помощью формулы (10) были определены величины энтомофильной активности растений.
По нашим расчетам, энтомофильная активность Sisymbrium loeselii L. (сем. Brassicaceae), Heracleum sibiricum L. (сем. Umbelliferae), Taraxacum officinale Wigg. (сем. Asteraceae), Kadenia dubia (Schkuhr) Lavrova et V.N. Tichom. (сем. Umbelliferae) составила 5.8 . 10-4; 1.5 . 10-4; 1.1 . 10-4; 0.5 . 10-4 бит/сек соответственно. Эти величины энтомофильной активности, по-видимому, соответствуют сформировавшимся биоэкологическим особенностям видов (в жизнедеятельности которых энтомофилия занимает разное место и проявляется специфично): S. loeselii – рудеральный, эврибионтный вид, активный энтомофил и очень продуктивный нектаронос, размножающийся семенами (Никитин, 1983; Denisow, 2005); H. sibiricum – R-эксплерент, инвазивный вид с перекрестным, энтомофильным опылением и семенным размножением, широко распространившийся на территории Средней России, а также Западной Сибири и Центральной Европы (Демьянова и др., 2007; Дозорова, Опарина, 2016); T. officinale – широко распространенный эврибионт, сочетающий перекрестное энтомофильное опыление с сильно выраженной способностью к самоопылению (автогамия, гейтеногамия) и апомиксису, способный размножаться вегетативно (Ермакова, 1990; Демьянова, 2010); K. dubia входит в российские и западно-европейские Красные книги, в фитоценозах встречается изредка и в небольшом обилии (Еленевский, Буланый, 2008; Ершова, 2012).
Заключение
Впервые предпринятый нами анализ позволяет заключить, что активность видов растений играет важную роль в организации, функционировании и развитии фитоценозов и экосистем, хотя понимается по-разному и потому в настоящее время изучается на основе независимо возникших подходов; их можно условно подразделить на ценотипический, статусный и эргонтический.
При ценотипическом подходе, т. е. в теории адаптивных (экологических) стратегий растений Раменского – Грайма (Раменский, 1971; Grime, 1974, 1977, 1979; Grime et al., 1988; Grime, Pierce, 2012; Работнов, 1985; Пьянков, Иванов, 2000; Пьянков и др., 2001; Pierce et al., 2007, 2013, 2014, 2017; Бетехтина, Веселкин, 2011), не используется понятие «активность растений», но при разграничении ценотипов (растений с разными стратегиями) обычно используется комплекс показателей, характеризующих энергию жизнедеятельности видов растений разных семейств, степень и скорость освоения ими ресурсов экосистемной среды в сравнении с конкурентами.
Статусный подход основан на положении о том, что активность вида растения есть мера его преуспевания в соответствующих ландшафтно-климатических условиях, т. е. выражается в результатах освоения видами экосистем, что адекватно степени экологической амплитуды, площади и характеру распространения (повсеместности и равномерности) основных типов местообитания вида внутри ландшафта, общего уровня численности вида (постоянства присутствия) в основных экотопах (Юрцев, 1966, 1968). Вследствие этого при статусном подходе активность каждого вида растений определяется ценотической значимостью, «местом», которое данный вид занимает среди других в системе сложившихся связей и отношений в растительном сообществе (Коновалова и др., 2014), т. е. по существу его экологической нишей.
Эргонтический подход (Савинов, 2015а, 2016а б, 2017), в отличие от статусного, позволяет оценивать величины разных форм происходящей, наблюдаемой активности видов (ценопопуляций), в частности морфогенетической, аккумуляционной, энтомофильной и других форм активности видов (ценопопуляций) растений. Величины таких форм активности могут быть интегрированы в обобщенном показателе активности. Таким образом, эргонтический подход, во-первых, дает возможность оценить вклад различных форм активности видов (ценопопуляций) в их наблюдаемое жизненное состояние в экосистеме, во-вторых, такой подход позволяет количественно обобщать данные о разных формах активности видов растений в едином показателе.
Есть основания полагать, что ценотипический, статусный и эргонтический подходы имеют точки соприкосновения, могут взаимно дополнять друг друга. На это определенно указывают следующие факты. Во-первых, под «стратегией вида» предложено понимать совокупность приспособлений, обеспечивающих виду возможность совместно обитать с другими организмами и занимать определенное положение в биоценозе (Работнов, 1975, 1985). Во-вторых, определено, что «экологическая стратегия» выражается в непрерывном адаптациогенезе растений в пределах соответствующей экосистемы, т. е. в их приспособительном взаимодействии с разнообразными биотическими и абиотическими факторами и объектами внешней среды (Работнов, 1985; Grime et al., 1988; Grime, Pierce, 2012). В-третьих, еще родоначальник статусного подхода Б. А. Юрцев (1968) предлагал по возможности определять дополнительные важные характеристики активности вида, в частности фенотипический и генотипический составы ценопопуляций. Наши исследования подтверждают это, в частности, показывают закономерное снижение морфогенетической активности ценопопуляций (определяемой с использованием фенов) в градиенте загрязнения тяжелыми металлами почв урбоэкосистем (Савинов и др., 2015; Савинов, Никитин, 2016). В-четвертых, исследования взаимосвязи типов экологических стратегий растений с их способностью к микоризообразованию и его интенсивностью (Grime et al., 1988; Cornelissen et al., 2001; Бетехтина, Веселкин, 2011) по существу характеризуют симбиотическую активность организмов (Савинов, 2016в).
Таким образом, проблема активности растений как фактора их развития в соответствующих экосистемах позволяет интегрировать данные из различных областей биологии и экологии и в новых, перспективных аспектах рассматривать вопросы организации и развития фитоценозов в экосистемах (Савинов, 2015б, 2016а, б, 2017). Наряду с этим нам представляется, что предпринятый анализ в отношении продуцентов укрепляет теоретическую базу и для исследований роли активности консументов и редуцентов в организации и функционировании различных типов экосистем и биосферы в целом.
Библиография
Алимов А. Ф., Богатов В. В., Голубков С. М. Продукционная гидробиология . СПб.: Наука, 2013. 343 с.
Бабич H. A., Ушакова С. Н. Активность видов сегетальной флоры питомников южнотаежной подзоны Вологодской области // Вестн. Северн. (Арктич.) федеральн. ун-та. Сер. Естественные науки. 2012. Вып. 2. С. 5–10.
Безель В. С., Жуйкова Т. В., Позолотина В. Н. Структура ценопопуляций одуванчика и специфика накопления тяжелых металлов // Экология. 1998. № 5. С. 376–382.
Большаков В. Н., Корытин Н. С., Кряжимский Ф. В., Шишмарев В. М. Новый подход к оценке стоимости биотических компонентов экосистем // Экология. 1998. № 5. С. 339–348.
Бернштейн Н. А. Очерки по физиологии движений и физиологии активности . М.: Медицина, 1966. 349 с.
Бетехтина А. А., Веселкин Д. В. Распространенность и интенсивность микоризообразования у травянистых растений Среднего Урала с разными типами экологических стратегий // Экология. 2011. № 3. С. 176–183.
Биологический энциклопедический словарь . М.: Сов. энциклопедия, 1986. 831 с.
Васильев А. Г. Эпигенетические основы фенетики: на пути к популяционной мерономии . Екатеринбург: Академкнига, 2005. 640 с.
Великанова Н. А., Гапонов С. П., Сливкин А. И. Оценка экологического состояния почв и лекарственного сырья (травы горца птичьего и листьев подорожника большого) по содержанию тяжелых металлов в городе Воронеже и его окрестностях // Вестник ВГУ. Сер. Химия. Биология. Фармация. 2012. № 2. С. 238–244.
Винер Н. Кибернетика, или управление и связь в животном и машине . М.: Советское радио, 1958. 215 с.
Воробейчик Е. Л., Хантемирова Е. В. Реакция лесных фитоценозов на техногенное загрязнение: зависимости доза – эффект // Экология. 1994. № 3. С. 31–43.
Гринфельд Э. К. Происхождение и развитие антофилии у насекомых . Л.: Изд-во ЛГУ, 1978. 208 с.
Грищенкова Н. Д., Власов Б. П. Антропогенное воздействие на видовой состав и количественное развитие высшей водной растительности Беларуси // Водные экосистемы: трофические уровни и проблемы поддержания биоразнообразия: Материалы Всерос. конф. Вологда, 2008. С. 43–45.
Демьянова Е. И. Антэкология . Пермь: Перм. гос. ун-т, 2010. 116 с.
Демьянова Е. И., Квиткина А. К., Лыков В. А. Особенности опыления Heracleum sibiricum L. и Seseli libanotis (L.) Koch (Аpiaceae) в Приуралье // Вестн. Перм. ун-та. 2007. Вып. 5. С. 6–13.
Дидух Я. П. Проблемы активности видов растений // Бот. журн. 1982. Т. 67. № 7. C. 925–935.
Дозорова С. В., Опарина С. Н. Изучение особенностей репродуктивной биологии Heracleum sibiricum L. и возможные причины его уязвимости // Молодежный научный форум: Естественные и медицинские науки. № 6 (34). М.: МЦНО, 2016. С. 13–18 . URL: http://nauchforum.ru/archive/MNF_nature/6(34).pdf.
Дуля О. В., Микрюков В. С., Воробейчик Е. Л. Стратегии адаптации Deschampsia caespitosa и Lychnis flos-cuculi к обитанию в условиях промышленного загрязнения: анализ с использованием зависимостей доза – эффект // Экология. 2013. № 4. С. 243–253.
Еленевский А. Г., Буланый Ю. И. Зонтичные (Apiaceae, Umbelliferae) Саратовской области // Бюл. Бот. сада Саратов. гос. ун-та. Саратов: Изд-во Сарат. ун-та, 2008. Вып. 7. С. 35–44.
Емельянов И. Г. Разнообразие и устойчивость биосистем // Успехи соврем. биологии. 1994. Т. 114. Вып. 3. С. 304–318.
Ермакова И. М. Одуванчик лекарственный // Биологическая флора Московской области. М.: Изд-во МГУ, 1990. С. 210–229.
Ершова Э. А. Антропогенная трансформация в пригородных сообществах с участием в травостое папоротника орляка (Pteridium pinetorum) // Растительный мир Азиатской России. 2012. № 2 (10). С. 132–138.
Жизнь растений . Т. 1. М.: Просвещение, 1974. 487 с.
Иванова Н. А., Юмагулова Э. Р. Эколого-физиологические механизмы адаптации и типы стратегии сосудистых растений верховых болот . Нижневартовск: Изд-во Нижневарт. гуманит. ун-та, 2009. 186 с.
Иорданский Н. Н. Организмы, виды и эволюция . М.: ЛИБРОКОМ, 2011. 176 с.
Ипатов В. С. Квантованность динамических рядов растительных сообществ // Бот. журн. 2016. Т. 101. № 4. С. 345–357.
Камшилов М. М. Организованность и эволюция // Журн. общ. биологии. 1970. Т. 31. № 2. С. 157–178.
Каратыгин И. В. Коэволюция грибов и растений . СПб.: Гидрометеоиздат, 1993. 118 с.
Коновалова И. С., Бабич Н. А., Марич С. Н. Фитоценотическая значимость сорных растений лесных питомников // Известия вузов. Лесной журн. 2014. № 1. С. 37–43.
Копылова Л. В. Особенности поглощения некоторых тяжелых металлов древесными растениями в условиях городской среды // Вестник ИрГСХА. 2011. Вып. 44. С. 91–99.
Кулагина Л. С. Оценка роли сосны обыкновенной (Pinus sylvestris L.) в оптимизации техногенной среды Башкирского Зауралья // Современные проблемы контроля качества природной и техногенной сред. Тамбов: Изд. дом ТГУ, 2009. С. 33–38.
Лебедева В. Х., Тиходеева М. Ю., Ипатов В. С. О неоднородности растительного покрова лугов и лесов // Бот. журн. 2016. Т. 101. № 4. С. 358–376.
Левченко В. Ф. Биосфера: этапы жизни (эволюция частей и целого) . СПб.: Свое издательство, 2012. 264 с.
Магомедмирзаев М. М., Гусейнова З. А., Алибегова А. Н., Магомедова С. М. Проблема адаптивных стратегий растений . Махачкала: Наука–Дагестан, 2013. 300 с.
Магомедмирзаев М. М., Гусейнова З. А. Репродуктивные адаптивные стратегии в эволюционно-морфологических преобразованиях семенных растений // Теоретические проблемы экологии и эволюции. Тольятти: Кассандра, 2015. С. 199–203.
Малышев Л. И. Флористическое районирование на основе количественных признаков // Бот. журн. 1973. Т. 58. № 11. С. 1581–1588.
Мелихова Г. И. Красная книга. Растения мира . М.: Эксмо, 2014. 96 с.
Минвалиев С. В., Павлова О. В. Ценотическая активность многолетних трав в условиях Приморского края // Вестн. Алтайского ГАУ. 2015. № 9 (131). С. 26–30.
Миркин Б. М., Усманов И. Ю., Наумова Л. Г. Типы стратегий растений: место в системах видовых классификаций и тенденции развития // Журн. общ. биологии. 1999. Т. 60. № 6. С. 581–595.
Налимова Н. В. Флористическое разнообразие и проблемы сохранения популяций редких видов растений ГПЗ «Присурский»: Автореф. дис. … канд. биол. наук . Сыктывкар, 2003. 23 с.
Налимова Н. В. Оценка внутриландшафтной активности популяций растений // Поливариантность развития организмов, популяций и сообществ. Йошкар-Ола: Мар. гос. ун-т. 2006. С. 118–123.
Никитин В. В. Сорные растения флоры СССР . Л.: Наука, 1983. 454 с.
Панкова Н. Л. Структура и динамика растительного покрова водоемов Окского заповедника / Тр. Окского гос. прир. биосф. заповедника. Вып. 31. Рязань: НП «Голос губернии», 2014. 166 с.
Проблемы эволюции биосферы / Ред. С. В. Рожнов. М.: ПИН РАН, 2013. 340 c.
Пьянков В. И., Иванов Л. А. Структура биомассы у растений бореальной зоны с разными типами экологических стратегий // Экология. 2000. № 1. С. 3–10.
Пьянков В. И., Иванов Л. А., Ламберс Х. Характеристика химического состава листьев растений бореальной зоны с разными типами экологических стратегий // Экология. 2001. № 4. С. 243–251.
Раменский Л. Г. Избранные работы. Проблемы и методы изучения растительного покрова . Л.: Наука, 1971. 334 с.
Работнов Т. А. Луговедение . М.: Изд-во МГУ, 1974. 384 с.
Работнов Т. А. Изучение ценотических популяций в целях выяснения «стратегии жизни» видов растений // Бюлл. МОИП. Отд. биол. 1975. Т. 80. № 2. С. 5–17.
Работнов Т. А. О типах стратегий растений // Экология. 1985. № 3. С. 3–12.
Рыженко О. В. Ценотическая активность многолетних трав на второй год жизни в зависимости от нормы азотных удобрений в условиях Приморского края // Вестн. КрасГАУ. 2012. Вып. 8. С. 48–51.
Савинов А. Б., Курганова Л. Н., Шекунов Ю. И. Интенсивность перекисного окисления липидов у Taraxacum officinale Wigg. и Vicia cracca L. в биотопах с разными уровнями загрязнения почв тяжелыми металлами // Экология. 2007. № 3. С. 191–197.
Савинов А. Б. Аутоценоз и демоценоз как симбиотические системы и биологические категории // Журн. общ. биологии. Т. 73. № 4. 2012. С. 284–301.
Савинов А. Б. Активность материальных систем, ее информационно-эволюционная роль, количественная и качественная оценка // ХXIХ Любищевские чтения. Современные проблемы эволюции и экологии. Ульяновск: УлГПУ, 2015а. С. 104–111.
Савинов А. Б. Активность растений как фактор их эволюции в древней и современной биосфере // Палеоботанический временник. Вып. 2. М.: ГЕОС, 2015б. С. 155–160.
Савинов А. Б., Никитин Ю. Д., Тарасова М. П. Особенности изменчивости листьев растений в городских фитоценозах разного состава (информационный аспект активности сообществ) // ХХIX Любищевские чтения. Современные проблемы эволюции и экологии. Ульяновск: УлГПУ, 2015. C. 479–484.
Савинов А. Б. Эволюционная теория активности систем // ХXХ Любищевские чтения. Современные проблемы эволюции и экологии. Ульяновск: УлГПУ, 2016а. С. 44–51.
Савинов А. Б. Теория активности систем и познание эволюции глобальных процессов // Эволюция: срезы, правила, прогнозы. Волгоград: Учитель, 2016б. С. 138–154.
Савинов А. Б. Активность компонентных сообществ паразитов наземных и водных животных // Тр. Центра паразитологии ИПЭЭ РАН. Т. 49: Фауна и экология паразитов. М.: КМК, 2016в. С. 145–147.
Савинов А. Б. Активность живых организмов как фактор их онтогенеза и эволюции // ХXХI Любищевские чтения. Современные проблемы экологии и эволюции. Ульяновск: УлГПУ, 2017. С. 66–73.
Савинов А. Б., Никитин Ю. Д. Фенотипическая изменчивость как проявление активности растений в сообществах (на примере урбоценозов) // ХXХ Любищевские чтения. Современные проблемы эволюции и экологии. Ульяновск: УлГПУ, 2016. С. 104–110.
Свириденко Б. Ф., Евженко К. С., Ефремов А. Н. Фитоценотическое значение зигнемовых водорослей (Zygnematales) на западно-сибирской равнине // Вестн. ТГПУ. 2013. № 8. С. 35–42.
Свириденко Б. Ф. Флора и растительность водоемов Северного Казахстана . Омск: Изд-во Омского ГПУ, 2000. 196 с.
Сибгатуллина М. Ш., Валеева Г. Р. Металлы в травянистых растениях с разными типами адаптивных стратегий // Юг России: экология, развитие. 2013. № 1. С. 72–81.
Сорокина В. С. Кормовые преференции имаго мух-журчалок (Diptera, Syrphidae) в условиях Южного Зауралья // Евразиатский энтомол. журн. 2003. Т. 2. Вып. 3. С. 197–214.
Тихонович И. А., Проворов Н. А. Симбиозы растений и микроорганизмов: молекулярная генетика агросистем будущего . СПб.: Изд-во С.-Петерб. ун-та, 2009. 210 с.
Уиттекер Р. Сообщества и экосистемы . М.: Прогресс, 1980. 326 с.
Усманов И. Ю., Рахманкулова З. Ф., Кулагин А. Ю. Экологическая физиология растений . М.: Логос, 2001. 224 с.
Чайковский Ю. В. Активный связный мир. Опыт теории эволюции жизни . М.: Т-во науч. изданий КМК, 2008. 726 с.
Чепинога В. В., Росбах С. А. Активность различных видов водной флоры Иркутско-Черемховской равнины // География и природные ресурсы. 2008. № 1. С. 97–104.
Шубина Н. В., Юрьев Ю. Л. Влияние выбросов металлургического производства на микроэлементный состав хвои сосны // Химия растит. сырья. 2009. № 3. С. 173–176.
Щелчкова М. В., Стручкова Л. К., Федоров И. А. Комплексное влияние тяжелых металлов на ферментативную активность и эффективное плодородие мерзлотной луговочерноземной почвы // Вестн. СВФУ. 2010. Т. 7. № 4. С. 16–21.
Юрцев Б. А. Гипоарктический ботанико-географический пояс и происхождение его флоры . М.; Л.: Наука, 1966. 95 с.
Юрцев Б. А. Флора Сунтар-Хаята. Проблемы истории высокогорных ландшафтов Северо-Востока Сибири . Л.: Наука, 1968. 235 с.
Angert A. L., Huxman T. E., Barron-Gafford G. A., Gerst K. L., Venable D. L. Linking growth strategies to long-term population dynamics in a guild of desert annuals // J. Ecol. 2007. Vol. 95. № 2. P. 321–331.
Biosphere Origin and Evolution / Dobretsov N. et al. (Eds.). Berlin: Springer, 2008. 427 р.
Carlyle C. N., Fraser L. H. A test of three juvenile plant competitive response strategies // J. Veg. Sci. 2006. Volk. 17. № 1. Р. 11–18.
Cerabolini B., Brusa G., Ceriani R. M., De Andreis R., Luzzaro A., Pierce S. Can CSR classification be generally applied outside Britain? // Plant Ecol. 2010. Vol. 210. № 2. P. 253–261.
Cornelissen J. H. C., Aerts R., Cerabolini B. et al. Carbon cycling traits of plant species are linked with mycorrhizal strategy // Oecologia. 2001. Vol. 129. №. 4. P. 611–619.
Craine J. M. Reconciling plant strategy theories of Grime and Tilman // J. Ecol. 2005. Vol. 93. № 6. Р. 1041–1052.
Craine J. M. Plant strategy theories: replies to Grime and Tilman // J. Ecol. 2007. Vol. 95. № 2. P. 235–240.
Denisow B. Nectar secretion of Sisymbrium loeselii L. in some ruderal рhytocenoses in the city of Lublin area // J. Apic. Sci. 2005. Vol. 49. № 2. Р. 51–58.
Franzaring J., Fangmeier A., Hunt R. On the consistencies between CSR plant strategies and Ellenberg ecological indicator values // J. Appl. Bot. Food Qual. 2007. Vol. 81. № 1. P. 86–94.
Grime J. P. Vegetation classification by reference to strategies // Nature. 1974. Vol. 250. P. 26–31.
Grime J. P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory // Amer. Natur. 1977. Vol. 111. № 982. P. 1169–1194.
Grime J. P. Plant strategies and vegetation processes. Chichester: Wiley and Sons, 1979. 222 p.
Grime J. P., Hodgson J. G., Hunt R. Comparative plant ecology: a functional approach to common British species. London: Unwin Hyman, 1988. 742 p.
Grime J. P. Plant strategy theories: a comment on Craine (2005) // J. Ecol. 2007. Vol. 95. № 2. Р. 227–230.
Grime J. P., Pierce S. The evolutionary strategies that shape ecosystems. Chichester: Wiley-Blackwell, 2012. 240 р.
Gurevitch J., Scheiner S. M., Gordon A. F. The ecology of plants. Sunderland: Sinauer Associates, 2002. 464 p.
Kosakivska I. V. Biomarkers of plants with different types of ecological strategies // Gen. Appl. Plant Physiol. 2008. Vol. 34. № 1/2. P. 113–126.
Kozanecka T., Chojnicki J., Kwasowski W. Content of heavy metals in plant from pollution-free regions // Pol. J. Environ. Studies. 2002. Vol. 11. № 4. Р. 395–399.
Li Y., Shipley B. An experimental test of CSR theory using a globally calibrated ordination method // PLOS. 2017. April 7. P. 1–17. URL: https: //doi.org/10.1371/journal.pone.0175404.
Photosynthesis and the Environment / N. R. Baker (Ed.). Kluwer: Academic Publishers, 1996. 574 p.
Pierce S., Luzzaro A., Caccianiga M., Ceriani, R. M., Cerabolini B. Disturbance is the principal α‐scale filter determining niche differentiation, coexistence and biodiversity in an alpine community // J. Ecol. 2007. Vol. 95. № 4. P. 698–706.
Pierce S., Brusa G., Vagge I., Cerabolini B. E. L. Allocating CSR plant functional types: the use of leaf economics and size traits to classify woody and herbaceous vascular plants // Funct. Ecol. 2013. Vol. 27. № 4. P. 1002–1010.
Pierce S., Vagge I., Brusa G., Cerabolini B. E. L. The intimacy between sexual traits and Grime’s CSR strategies for orchids coexisting in semi-natural calcareous grassland at the Olive Lawn // Plant. Ecol. 2014. Vol. 215. P. 495–505.
Pierce S., Negreiros D., Cerabolini B. E. L., Kattge J., Díaz S., Kleyer M., et al. A global method for calculating plant CSR ecological strategies applied across biomes world-wide // Funct. Ecol. 2017. Vol. 31. № 2. P. 444–457.
Smith S. E., Read D. J. Mycorrhizal symbiosis. Oxford: Elsevier, 2008. 804 p.
Susiluoto S., Berninger F. Interactions between morphological and physiological drought responses in Eucalyptus microtheca // Silva Fennica. 2007. Vol. 41. № 2. P. 221–233.
Suzuki H., Kumagai H., Oohashi K., Sakamoto K., Inubushi K., Enomoto S. Transport of trace elements through the hyphae of an arbuscular mycorrhizal fungus into marigold determined by the multitracer technique // Soil Sci. Plant Nutr. 2001. Vol. 47. № 1. Р. 131–137.
Tilman D. Resource competition and plant traits: a response to Craine et al. 2005 // J. Ecol. 2007. Vol. 95. № 2. P. 231–234.
Благодарности
Авторы благодарны рецензентам, а также А. А. Протасову, Ч. М. Нигматуллину, А. А. Махрову за полезное обсуждение выдвинутых положений, что позволило дополнить статью важными литературными сведениями, расширить круг обсуждаемых вопросов; выражаем также признательность В. П. Воротникову за помощь в вопросах таксономии растений.

 © 2011 - 2026
© 2011 - 2026