



Введение
Вопросы регуляции температуры тела рептилиями издавна привлекают внимание специалистов. Опубликовано множество работ, в которых так или иначе рассматриваются результаты регистрации температуры тела: ее уровень у разных видов, размах ее варьирования, критические пределы и т. д. Некоторые из них достойны называться основополагающими (Brattstrom, 1965; Avery, 1982; Huey, 1984; Seebacher, 2005; Seebacher, Franclin, 2005; Angilletta, 2009). Особое внимание уделяется способам регуляции температуры. Обычно основным называют поведенческий. Попытаемся и мы внести свою лепту в общую копилку термобиологии рептилий. При этом постараемся воздерживаться от обобщений и далеко идущих выводов.
Аналитический обзор
В соответствии с решением Commission for Thermal Physiology of the International Union of Physiological Sciences (Glossary of terms for Thermal Physiology, 2003) мы считаем основными температурными показателями рептилий следующие.
Температура тела (body temperature) – температура, измеренная в начальном отделе пищевода, кроме видов, у которых измерение в пищеводе невозможно (пискливый геккончик, круглоголовка-вертихвостка). У них температура измеряется в клоаке. При помещении регистратора iBDL в тело животного принимаем за температуру тела или подкожную, или полостную.
Допустимый диапазон температур (tolerated temperature range) – минимальные и максимальные значения, хотя бы раз зарегистрированные в течение сезона активности.
Термопреферендум (thermopreferendum) – минимальный температурный диапазон, на который приходится 50 % регистраций температур тела у данного вида (популяции).
Внешняя температура (ambient temperature). Измеряется температура субстрата, на котором было замечено животное, и температура приземного воздуха на высоте не более 1–3 см, т. е. на высоте тела животного. Регистрировать температуру тела выше или на метеостанции, на наш взгляд, бессмысленно.
Соотношение температуры тела и внешних температур. Эта зависимость в большинстве случаев не линейна, поэтому для определения силы связи температур нами применяется корреляционное отношение (η). Одним из показателей этого соотношения может считаться «абсолютный температурный оптимум» (АТО). Существует довольно нестабильный уровень температуры тела, достигаемый путем инсоляции, когда температура тела животного становится равной внешней температуре (обычно температуре субстрата). При достижении такого уровня у животного возникает потребность не допустить дальнейшего подъема температуры. Такую температуру мы называем «абсолютным температурным оптимумом». После этого внешняя температура продолжает расти, а температура тела, хотя и увеличивается, но с заметным отставанием (Литвинов, 2008).
Все полученные за время полевых работ температуры субстрата (возможно использование и температур воздуха) разбиваются на классы вариационного ряда по формуле: K = 1 + 3,32 lg n, где K – число классов, n – объем выборки. Величина классового интервала определялась по формуле: 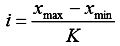 , где i – величина классового интервала, xmax – максимальная, а xmin – минимальная варианты. Каждому значению внешней температуры соответствует своя температура животного, замеченного на этом субстрате. Получаются среднестатистические значения внешних и внутренних температур для каждого из этих классов. Две кривые на графике, построенном на основе расчетов, перекрещиваются в определенной точке, которая и соответствует температуре абсолютного оптимума. Такой график достаточно точно иллюстрирует действительное соотношение этих температур.
, где i – величина классового интервала, xmax – максимальная, а xmin – минимальная варианты. Каждому значению внешней температуры соответствует своя температура животного, замеченного на этом субстрате. Получаются среднестатистические значения внешних и внутренних температур для каждого из этих классов. Две кривые на графике, построенном на основе расчетов, перекрещиваются в определенной точке, которая и соответствует температуре абсолютного оптимума. Такой график достаточно точно иллюстрирует действительное соотношение этих температур.
Рассмотрим процесс определения точки абсолютного температурного оптимума на примере обыкновенного ужа из Камского Предуралья. Данные по средним температурам поверхности субстрата и пищевода представлены в табл. 1, а их графическое отображение на рис. 1.
Таблица 1. Средние температуры поверхности субстрата и пищевода по классам у обыкновенного ужа (n = 40) из Камского Предуралья
| Классы по температуре | Средние значения температуры поверхности субстрата | Средние значения температуры пищевода |
| 1 | 11.0 | 28.1 |
| 2 | 14.8 | 19.9 |
| 3 | 18.7 | 26.0 |
| 4 | 22.5 | 27.7 |
| 5 | 26.3 | 30.2 |
| 6 | 30.2 | 32.3 |
| 7 | 34.0 | 31.1 |
| 8 | 37.9 | 33.5 |
График, отражающий температуру поверхности субстрата, представляет собой абсолютно прямую линию, что позволяет выразить его с помощью уравнения. График же температуры пищевода имеет несколько точек перегиба. Кроме того, возможно наличие двух и более точек пересечения графиков. На наш взгляд, это искажает реальную картину. Для более точного определения точки абсолютного температурного оптимума мы предлагаем использовать линейную регрессию при обработке данных по температуре пищевода. Этот метод приводит график температуры пищевода в линейную форму, что также позволит выразить его при помощи уравнения (рис. 1).
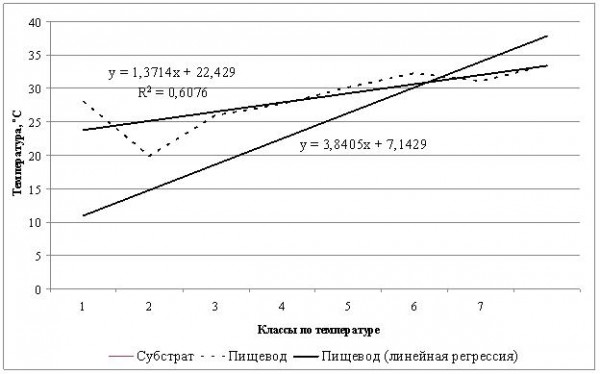
Рис. 1. Соотношение температур поверхности субстрата и пищевода у обыкновенного ужа (n = 40) из Камского Предуралья
Fig. 1. The ratio the between the surface temperature of the substrate and the esophagus in an grass snake (n = 40) of the Kamsky Preduralye
В таком случае точка абсолютного температурного оптимума находится не визуально, а вычисляется на основе решения системы двух уравнений.
В рассматриваемом нами случае температура поверхности субстрата выражается уравнением y = 3.8362x + 7.1638, а результат линейной регрессии температуры пищевода – y = 1.3716x + 22.413. При решении данной системы уравнений точка абсолютного температурного оптимума оказывается равной 30.9 ºС.
В виде обобщающей количественной характеристики для всей совокупности полученных нами данных использовалась средняя арифметическая. Поясним, с чем связан наш выбор.
Мы сразу же отказались от использования структурных средних (таких как мода и медиана), т. к. они в первую очередь направлены на работы с дискретными величинами, в то время как температура является величиной непрерывной. Чем менее точным является измерительный прибор, тем более дискретный вид принимают полученные температурные значения, однако при использовании высокоточных датчиков полученный ряд значений практически непрерывен, соответственно, структурные средние зачастую дают неверную картину.
Из степенных средних (арифметической, гармонической, геометрической, квадратической) нами используется средняя арифметическая в связи с тем, что практически все полученные данные относятся к состоянию полной активности рептилий (то есть в массив не входят данные по рептилиям из убежищ и в период нагревания), соответственно, простое арифметическое усреднение дает вполне корректную картину.
По поводу форм термоадаптивного поведения отметим, что их, на наш взгляд, не так много. По сути – две или три, если считать еще и ночную неактивность в укрытии у видов с дневной активностью.
Первая - это нагревание до уровня оптимальной температуры тела после выхода из убежища, причем это нагревание не обязательно утреннее. Первый выход из норы может быть как утром, так и позже в зависимости от состояния животного, удельной мощности видимого света, уровня внешней температуры. Иначе говоря, не всякая ящерица, находясь у своей норы, обязательно только что вышла и нагревается. Реализуя эту фазу поведения, животное достигает подъема температуры своего тела до нижней границы диапазона температурного оптимума. Нагревание может осуществляться, когда животное чуть показалось из норы, высунув только голову, или находится около убежища. Допускаем, что эта фаза поведения может начинаться еще в норе, особенно если нора расположена не глубоко и температура в ней поднимается с подъемом внешней температуры.
Вторая форма – это контроль за своей температурой во время суточной активности (термостабилизирующее поведение). В этот период животное может временно находиться в норе или в тени, снижая свою температуру, и затем снова нагреваться. В течение дня это может повторяться многократно. Это и первое нагревание, и последующее за ним остывание, и второе, и третье, и т. д. (рис. 2).
Рис. 2. Суточная динамика температуры тела обыкновенной гадюки
Fig. 2. Daily dynamics of body temperature of Common European adder
В особо жаркие часы дня летом животное может задерживаться в убежищах надолго (на несколько часов). Тогда речь может идти о двухпиковой дневной активности. По поводу используемого иногда понятия «термонейтральное поведение» можно сказать следующее. В этой фазе животное наиболее активно питается, спаривается, контролирует территорию и т. д. Употреблять этот термин следует с осторожностью, понимая, что на самом деле у эктотермов термонейтрального в полном смысле этого слова поведения быть не может. Допускаем, что оно может проявляться только в рамках оптимумов температуры тела и внешней температуры.
Вечернее остывание вряд ли можно считать настоящей формой поведения, т. к. это вынужденное явление в результате снижения внешней температуры, причем остывание продолжается и в укрытии, превращаясь в ночное, длящееся всю ночь, по крайней мере, у обыкновенной гадюки (Литвинов, Четанов, 2014). Оно обычно обусловлено вечерним снижением внешней температуры или снижением температуры при похолодании. Животное при этом некоторое время остается в активном состоянии, пока температура его тела не достигнет уровня допустимого минимума, соответствующего виду, подвиду, популяции, определенному времени года, возрасту, полу. После этого следует уход в убежище. Ночное пребывание в укрытии стабилизирует температуру тела животного на уровне, не опасном для жизни, хотя и в этом случае температура не постоянна, а ее уровень следует за уровнем внешней температуры.
Остановимся на размерах и окраске тела, которые, видимо, имеют термоадаптивное значение. В условиях недостатка тепла быстрый нагрев предпочтительнее, т. к. позволяет животному достичь оптимальной подвижности за относительно короткое время. Быстрое остывание в этом случае не выглядит адаптивным, хотя в конце дневной активности в укрытии, например в норе, любая скорость остывания, скорее всего, не имеет особого значения.
В условиях избытка тепла повышенная скорость утреннего нагрева, возможно, предпочтительнее, чем замедленная, все по той же причине. Скорость остывания, вероятно, также не играет существенной роли.
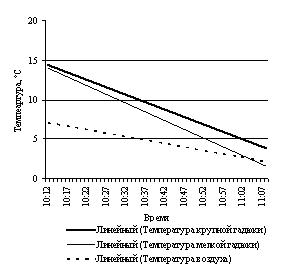
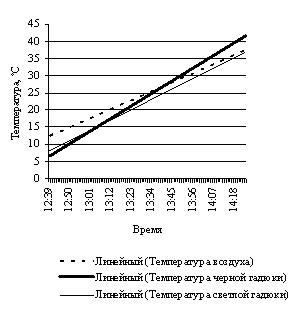
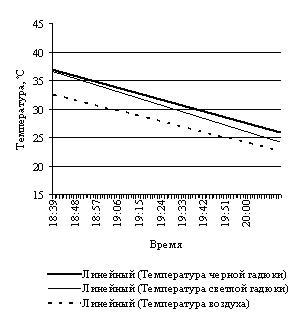
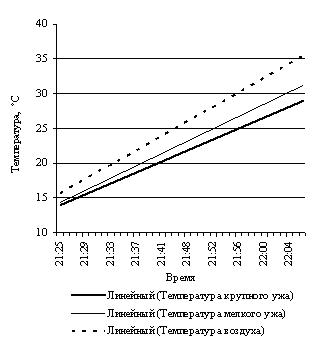
Эксперименты с нагреванием и последующим охлаждением крупной и небольшщой черной особей, черной и светлоокрашенной особей обыкновенной гадюки примерно одинакового размера, особей обыкновенного ужа разного размера и осоебей разноцветных ящурок более темного и более светлого оттенков окраски спины, на наш взгляд, могут в какой-то мере иллюстрировать вышесказанное (рис. 3).
A B
С D
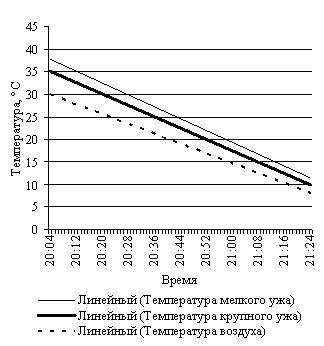
E F
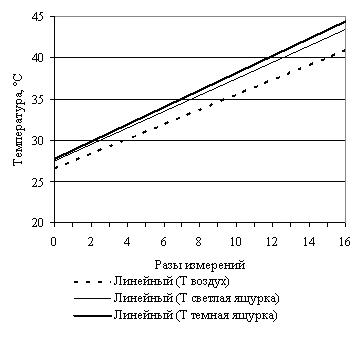
G
Рис. 3. Эксперименты с нагреванием и охлаждением рептилий: А – нагрев крупной и мелкой обыкновенных гадюк; B – охлаждение крупной и мелкой обыкновенных гадюк; С – нагрев черной и светлой обыкновенных гадюк; D – охлаждение черной и светлой обыкновенных гадюк; E – нагрев крупного и мелкого обыкновенных ужей; F – охлаждение крупного и мелкого обыкновенных ужей; G – нагрев темной и светлой разноцветных ящурок
Fig. 3. Experiments of heating and cooling reptiles: A – heating of large and small Common European adders; B – cooling of large and small Common European adders; C – heating of black and light Common European adders; D – cooling of black and light Common European adders; E – heating of large and small grass snakes; F – cooling of large and small grass snakes; G – heating of dark and light colored steppe-runners
Крупная гадюка (L.+L.cd. 695 мм) нагревается несколько медленнее, чем мелкая (L.+L.cd. 475 мм): 0.20 град./мин по сравнению с 0.22 град./мин. Остывает также медленнее: 0.37 град./мин по сравнению с 0.40 град./мин.
Температура тела гадюки средних размеров, находящейся в экспериментальной норе в июне в 8 ч., то есть перед ее выходом для обогрева, составляет по результатам 15 регистраций 15–18 ºС с небольшим округлением. Исходя из этого некрупная гадюка, покинувшая нору, нагревается до средней температуры, определенной для светлой морфы в 27.3 ± 0.37 ºС приблизительно за 51 мин., а крупная – за 57 мин. И у крупной, и у мелкой змей сила связи их температуры с температурой воздуха в климатической камере высока и практически не отличается: r = 0.94 ± 0.009 для первой и r = 0.96 ± 0.006 для второй (t = 1.89; Р > 0.05).
Подобную тенденцию демонстрирует и результат эксперимента с обыкновенными ужами разных размеров: некрупным (L.+L.cd. 565 мм) и очень крупным (L.+L.cd. 1110 мм).
При первом нагревании скорость роста температуры тела крупного ужа (0.13 град./мин) меньше скорости нагревания более мелкого (0.16 град./мин). То же соотношение скоростей сохраняется и при втором более интенсивном нагревании, соответственно 0.40 град./мин и 0.43 град./мин. Охлаждались оба ужа с одинаковой скоростью 0.30 град./мин.
В таком же эксперименте с утренним выходом из норы, как и с обыкновенными гадюками, крупный уж достигнет своей средней температуры активности в 25.3 ± 0.17 ºС примерно за 21 мин., а мелкий за 19 мин.
Некрупное животное с его большей относительной поверхностью тела по сравнению с более крупным обладает меньшей тепловой инерцией, то есть быстрее нагревается и быстрее остывает согласно физическим законам. Разница в скорости изменения температуры тела касается прежде всего нагревания и в меньшей степени охлаждения. Возможно, разница в достижении оптимальной температуры тела в 5–10 мин. покажется не столь существенной, но если речь идет о возможности быстрейшего обретения максимальной подвижности, необходимой для защиты от нападения, то ее как раз и может обеспечить приоритет в скорости нагревания.
Существуют ли температурные отличия между светлоокрашенными и черными обыкновенными гадюками (табл. 2)?
Таблица 2. Температурная характеристика черной и светлой морф обыкновенной гадюки в Камском Предуралье
| Параметр | Светлая морфа n=160 | Чёрная морфа n=42 | Достоверность отличий |
| L.+L.cd. (мм) | 573±7.38 | 577±20.3 | t=0.70; Р>0.05 |
| Температура воздуха в5 см(ºC) | 22.2±0.61 | 22.5±1.52 | t=1.36; Р>0.05 |
| Температура субстрата (ºC) | 24.8±0.52 | 21.4±0.99 | t=30.9; Р |
| Температура в пищеводе (ºC) | 28.2±0.36 | 26.5±0.79 | t=21.25; Р |
| Температура спины (ºC) | 24.3±0.41 | 23.3±0.78 | t=14.6; Р |
| Температура живота (ºC) | 23.9±0.38 | 23.0±0.77 | t=11.0; Р |
| Мощность ультрафиолета (Вт/м2) | 7.0±1.04 | 3.5±2.04 | t=9.2; Р |
| Мощность видимого света (Вт/м2) | 164.6±18.00 | 125.0±9.59 | t=34.7; Р |
| Падающее тепло (Вт/м2) | 37.3±4.72 | 29.5±4.97 | t=3.25; Р |
| Возвращённое тепло (Вт/м2) | 18.2±2.63 | 14.8±4.18 | t=2.43; Р |
| Сила связи температур воздух/пищевод (h) | 0.56±0.08 | 0.64±0.04 | t=0.36; Р>0.05 |
| Сила связи температур субстрат/пищевод (h) | 0.74±0.04 | 0.87±0.04 | t=1.12; Р>0.05 |
| Сила влияния темпер. воздуха на темпер. пищевода (%) (h2) | 23.49 | 33.78 | |
| Сила влияния темпер. субстрата на темпер. пищевода (%)(h2) | 49.77% | 74.61% |
Нами указывалось ранее на заметное биотопическое разобщение цветовых морф обыкновенной гадюки (Литвинов, 2004). Светлая морфа преимущественно обитает на склонах южной экспозиции, черная – на опушках и болотах. Весь север Прикамья занят исключительно черной морфой, а юго-восток преимущественно светлоокрашенной. Они демонстрируют заметно различную термофильность. Светлая морфа обитает на более теплом грунте, у нее выше температуры тела, сильнее воздействие на нее потока солнечной радиации. Тем не менее сила связи температуры тела с внешними температурами и сила влияния этих температур на температуру тела у этих двух морф отличается незначительно.
В одном из экспериментов нагревание гадюк проводилось от начальной температуры в клоаке у обеих змей 21.4 ºС до 37.5 ºС у черной и 34.6 ºС у светлой. За одно и то же время в 64 мин. температура черной поднялась на 16.1º со скоростью 0.25 град./мин, а температура светлой гадюки – на 13.2º со скоростью 0.21 град./мин. Соответственно, черная гадюка может достичь верхнего предела своего природного температурного оптимума (для температуры тела это 30.0 ºС) примерно за 34 мин. Светлоокрашенная гадюка достигает этого предела примерно за 46 мин. (31.0 ºС). Таким образом, черная гадюка получает заметное преимущество перед светлоокрашенной в скорости нагревания.
Охлаждение гадюк происходило в течение 120 мин. За это время черная гадюка снизила свою температуру с 25.0 ºС, то есть с оптимальной для нее температуры, до 6.9 ºС на 18.1º. Светлая гадюка за это же время снизила свою температуру тоже с 25.0 ºС до 6.1º на 18.8º.
Сходные результаты были получены и при проведении экспериментов с разноцветной ящуркой. Нагревали двух самок ящурок одинакового размера более светлого и темного оттенков окраски спины в течение 40 мин. За это время температура поверхности спины темной ящурки поднялась с 27.5 ºС до 43.0 ºС на 15.5º. Температура спины светлой ящурки выросла с исходной 27.5 ºС до 41.6 ºС на 14.1º. Скорость нагревания спины темной ящурки составила 0.39 град./мин, светлой ящурки – 0.35 град./мин. Оптимум температуры тела для этих ящурок из Нижнего Поволжья составляет 28.0–33.5 ºС. Таким образом, чтобы достичь верхнего предела оптимума, более темная ящурка должна нагреваться за 86 мин., а светлая – за 96 мин.
Заключение
Напрашивается вывод, что черная или даже просто темная окраска верхней части тела у рептилий дает некоторые преимущества в нагревании, сокращая время, затрачиваемое на этот процесс. Пресмыкающиеся, не успевшие нагреться до оптимальной температуры и поэтому не столь подвижные, могут быть уязвимы для врагов в период утреннего нагревания. Темные особи, таким образом, должны оказываться в более выгодном положении, т. к. быстрее нагреваются. Вечернее охлаждение не дает таких преимуществ ни темным, ни светлым особям, поскольку в абсолютном большинстве случаев оно происходит в укрытии, то есть в норе, где уязвимость не играет роли. В условиях сильно заросших биотопов черная окраска обыкновенной гадюки, скорее всего, термоадаптивна, мало демаскирует ее в силу закрытости биотопов. В условия остепненных склонов юго-востока Предуралья черный цвет не нужен и требованиям термоадаптивности соответствует серый, к тому же этот цвет хорошо маскирует гадюк на фоне относительно редкой растительности и известняковых и гипсовых обнажений.
Тем не менее нам бы не хотелось абсолютизировать вопрос о размерах тела и роли окраски, поскольку в природе существует баланс адаптаций, когда наиболее существенной, а значит, подхваченной отбором, является та, которая наиболее значима в существующих условиях среды.
Библиография
Литвинов Н. А. Термобиологические исследования // Змеи Волжско-Камского края. Самара: Изд-во Самарского научного центра РАН, 2004. С. 127–140.
Литвинов Н. А., Четанов Н. А. Непрерывная регистрация температуры тела обыкновенной гадюки летом // Известия Самарского НЦ РАН. 2014. Т. 16, № 5(1). С. 430–435.
Angilletta M. Thermal Adaptation. A Theoretical and Empirical Synthesis. Oxford University Press, 2009. 289 p.
Avery R. A. Field Studies of Body Temperatures and Thermoregulations // Biology of the Reptilia. 1982. Vol. 12. Р. 93–166.
Brattstrom В. Н. Body Temperatures of Reptiles // Amer. Midland Naturalist. 1965. Vol. 73, № 2. P. 376–422.
Glossary of terms for thermal physiology. Third Edition revised by The Commission for Thermal Physiology of the International Union of Physiological Sciences (IUPS Thermal Commission) // Journal of Thermal Biology. 2003. № 28. Р. 75–106.
Guillon М., Guiller G., DeNardo D., Lourdais O. Microclimate preferences correlate with contrasted evaporative water loss in parapatric vipers at their contact Zone // Can. J. Zool. 2014. Р. 81–86.
Elsworth P., Seebacher F., Franklin C. Sustained swimming performance in crocodiles (Crocodylus porosus): effects of body size and temperature // Journal of Herpetology. 2003. № 37(2). Р. 363–368.
Huey R. B. Temperature, Physiology, and the Ecology of Reptiles // Biology of the Reptilia. London: Academic Press Inc., 1984. P. 25–91.
Lourdais O., Guillon М., DeNardo D., Blouin-Demers G. Cold climate specialization: Adaptive covariation between metabolic rate and thermoregulation in pregnant vipers // Physiology & Behavior. 2013. № 119. Р. 149–155.
Seebacher F., Elsworth P., Franklin C. Ontogenetic changes of swimming kinematics in a semi-aquatic reptile (Crocodylus porosus) // Australian Journal of Zoology. 2003. № 51(1). Р. 15–24.
Seebacher F., Franklin C. The effect of heat transfer mode on heart rate responses and hysteresis during heating and cooling in the estuarine crocodile Crocodylus porosus // The Journal of Experimental Biology. 2003. № 206(7). Р. 1143–1151.
Seebacher F., Elsey R., Trosclair P. Body temperature null distributions in reptiles with nonzero heat capacity: Seasonal thermoregulation in the American alligator (Alligator mississippiensis) // Physiological and Biochemical Zoology: ecological and evolutionary approaches. 2003. № 76(3). Р. 348–359.
Seebacher F. A review of thermoregulation and physiological performance in reptiles: what is the role of phenotypic flexibility? // J. Comp. Physiol. B. 2005. Р. 175.
Seebacher F., Franclin C. E. Physiological mechanisms of thermoregulation in reptiles: a review // J. Comp. Physiol. B. 2005. Р. 175.
Благодарности
Выражаю глубокую благодарность своим коллегам Н. А. Четанову, С. В. Ганщук и С. А. Шуракову за помощь в настоящем исследовании.

 © 2011 - 2026
© 2011 - 2026