



Введение
Показатели флуктуирующей асимметрии широко используются при характеристике состояния природных популяций, их динамики и структуры, как обоснование при установлении популяционных границ, для изучения микроэволюционных преобразований, в качестве индикаторов отклонения условий среды от оптимальных в целях биомониторинга и т. д. (Palmer, Strobeck, 2003; Захаров, 2001; Методические…, 2003; Гелашвили и др., 2004; Зорина, 2010). Несмотря на активное применение метрик асимметрии с целью изучения надорганизменных биосистем, ее величина отражает внутрииндивидуальную изменчивость фенотипа, проявляется на уровне отдельного организма. Она характеризует случайную изменчивость развития особи в пределах ее нормы реакции и используется для оценки «стабильности развития» и «онтогенетического шума» (Mitton, 1993; Leung, Forbes, 1997; Захаров, 2001; Рalmer, Strobeck, 2003; Зорина, 2010).
Количество публикаций, в которых обсуждаются вопросы естественной динамики показателей флуктуирующей асимметрии в процессе онтогенеза отдельной особи, явно недостаточно для полноценного изучения фоновой изменчивости ее величины в природных условиях (Developmental..., 2003; Зорина, Коросов, 2009; Зорина, 2010). Асимметрия отдельных признаков или их комплекса оценивается для особи, находящейся на определенной стадии развития, а для ее органов – в конкретный период морфогенеза. Однако в зависимости от возраста или стадии морфогенеза величина и изменчивость многих признаков может значительно меняться, так же как само качество особи со временем не остается постоянным (Cheverud et al., 1983; Kimball et al., 1997; Leung, Forbes, 1997; Kellner, Alford, 2003).
Без серьезного внимания к проблемам теоретического осмысления явления использование показателей асимметрии может быть биологически бесполезно (Рalmer, Strobeck, 2003). Изучение природных закономерностей на онтогенетическом уровне организации жизни необходимо для понимания механизма процессов, протекающих на других уровнях, в том числе на популяционном (Leung, Forbes, 1997; Developmental…, 2003). Сложность реализации наследственной информации при развитии как отдельных признаков, так и всего фенотипа особи определяет проблемы, возникающие при изучении данного вопроса. В настоящее время выделяют по меньшей мере семь моделей формирования флуктуирующей асимметрии (Kellner, Alford, 2003).
Для достижения цели нашего исследования – изучения изменчивости величины флуктуирующей асимметрии в процессе индивидуального развития березы повислой – решались следующие задачи:
- Оценить изменение показателей асимметрии близкородственных особей березы повислой в зависимости от их возрастного состояния.
- Изучить динамику величины асимметрии метрических признаков листа в течение его морфогенеза.
- Установить модель формирования уровня асимметрии в течение индивидуального развития березы повислой.
Материалы
Сбор листьев березы повислой проводился на о. Климецкий (Онежское озеро, Республика Карелия) во второй половине лета 2007 г. На зарастающем сенокосном лугу (более 2 км от береговой линии острова, равнина с ровницами, на лугу участки с разной степенью и возрастом зарастания) были выбраны по 10 деревьев только тех возрастных состояний, которые хорошо отличаются друг от друга по внешним признакам: имматурные (im), виргинильные (vir), генеративные молодые (g1), генеративные средневозрастные (g2), генеративные старые (g3), постгенеративные сенильные (s) (Лантратова, Сонина, 2005). При формировании выборок стремились учесть однородные условия произрастания деревьев, исключающие любые факторы внешней среды, существенно влияющие на асимметрию листьев, кроме их возрастного состояния (Зорина, Коросов, 2009; Зорина, 2010). Другими словами, были сформированы значительные объемы выборок в однородных условиях из берез со сходными морфофизиологическими характеристиками. Близкородственность особей определялась их произрастанием в непосредственной близости друг от друга в одном биотопе, по периферии окруженном ивняком, осинником или сосняком.
С брахибластов каждого отдельного дерева собиралось по 10 листьев равномерно с разных участков кроны. Таким образом, было сформировано шесть выборок разных возрастных состояний березы повислой, каждая объемом в 100 листьев: nim = nvir = ng1 = ng2 = ng3 = ns = 100.
Для решения второй задачи материал собирался с 10 особей березы повислой имматурного возрастного состояния, произрастающих в тех же условиях, которые были описаны выше. На фоне миллиметровой бумаги одни и те же листья (по 10 с каждого дерева), зафиксированные сверху тонким прозрачным пластиковым держателем, фотографировались на протяжении всего вегетационного периода пять раз: 25.05, 25.06, 25.07, 25.08, 25.09.2007 г. Во избежание путаницы экземпляров побеги с исследуемыми листьями метили яркой краской и съемка проводилась всегда в одинаковой последовательности снизу вверх. Было сформировано пять выборок разных стадий морфогенеза листа, каждая объемом в 100 листьев (10д.•10л.): n05 = n06 = n07 = n08 = n09 = 100.
Методы
Листья сканировались с тыльной стороны на фоне миллиметровой бумаги. Геометрическая коррекция (Коросов, Коросов, 2006) искаженных растровых изображений (фотографий) проводилась в среде QGIS.
В работе использовались данные по пяти пластическим (метрическим) свойствам (рис. 1). Промеры признаков электронных изображений левой (Lij) и правой (Rij) половинок листовой пластинки выполнили в реальном масштабе 1:1. Регистрация растров в среде MapInfo проводилась в проекции Широта/Долгота (для создания прямоугольных шкал) с единицами измерения – градусами в десятичном формате. В определенной последовательности ставились точки начала и конца промеров, вычислялись координаты этих точек (Коросов, Коросов, 2006), значения которых экспортировались в программу Excel. Расстояние между точками рассчитывалось как корень квадратный из суммы квадратов разницы координат x и y.

Рис. 1. Схема промеров признаков на левой (Lj) и правой (Rj) сторонах листовой пластинки березы повислой: j = 1 – ширина, 2, 3 – длина второй и третьей жилок второго порядка, 4 – расстояние между основаниями жилок второго порядка, 5 – расстояние между концами этих жилок
Fig. 1. Measurement scheme of signs on left (Lj) and right (Rj) sides of a silver birch leaf: j = 1 – width, 2, 3 – length of second and third ribs of the the second order, 4 – distance between rib bases of the second order, 5 – distance between rib ends
В результате в среде Excel каждая выборка образовала матрицу данных размером 2m•n (i = 1, 2, … n – номер листа, n = 100; j = 1, 2, … m – номер признака, m = 5). Для реализации первой задачи было сформировано 6 выборок, характеризующих разные возрастные состояния березы повислой; для второй – 5 выборок разных этапов морфогенеза листовой пластинки данного вида.
Из всего многообразия показателей (faij, faj) и индексов асимметрии (fai, FA) использовали простое отличие между сторонами и оценку на основе нормированного отклонения (табл. 1). Механизм расчета метрик асимметрии подробно описан в более ранних работах и в данной статье не приводится (Зорина, Коросов, 2009; Зорина, 2010, 2012, 2013).
Таблица 1. Формулы расчета показателей и индексов флуктуирующей асимметрии (Mj, Sj – среднее арифметическое и стандартное отклонение j-го билатерального признака)
Table 1.Formulas for the calculation of fluctuating asymmetry indicators and indexes (Mj, Sj – arithmetic mean and standard deviation of the j-th bilateral trait)
| № | Показатели асимметрии | |||
| Одной особи по одному признаку, faij | Выборки по одному признаку, faj | Одной особи по всем признакам, fai | Интегральный индекс, FA | |
| 1 | faij = (Lij ─ Rij) | faj = S^2(faij) | ─ | ─ |
| 2 | faij = (t(Lij) ─ t(Rij))
t(Lij) = (Lij ─ Mj) / Sj t(Rij) = (Rij ─ Mj) / Sj |
faj = S^2(faij) | fai = 1/m • ∑j=1→m (t(Lij) ─ t(Rij)) | FA = S^2(fai) |
Статистическая обработка была выполнена в программах Excel и StatGraphics с использованием параметрических или непараметрических критериев в зависимости от соответствия распределения показателя асимметрии нормальному закону.
Результаты
Для всех выборок был доказан флуктуирующий характер асимметрии пластических вегетативных признаков листа. Распределение разности между промерами сторон достоверно от нормального закона не отличается. Значимость асимметрии и эксцесса не выявлена, средние арифметические равны нулю (табл. 2, рис. 2). Для выбранных метрических признаков асимметричная изменчивость не несет генетической компоненты (которая проявляется в виде направленной или антисимметрии), что также подтверждается литературными данными (Зорина, Коросов, 2009; Зорина, 2010).
Таблица 2. Уровень значимости отличия средней (M), асимметрии (A), эксцесса (E) от нуля и отличие распределения показателя (Lij – Rij) признаков листа березы повислой от нормального закона для выборки nim (жирным шрифтом выделены достоверные отличия для α < 0,05)
Table 2. Significance Level of the difference between mean (M), asymmetry (A), kurtosis (E) from zero and the difference between the distribution of indicator (Lij – Rij) of silver birch leaf signs and normal distribution for nim sampling (in bold are significant differences for α < 0,05 are shown in bold)
| Нулевая гипотеза | Статистические критерии | Номер признака j | ||||
| 1 | 2 | 3 | 4 | 5 | ||
| Соответствие нормальному закону | χ2 Пирсона | 0,91 | 0,12 | 0,19 | 0,88 | 0,09 |
| W Шапиро – Уилкса | 0,11 | 0,12 | 0,84 | 0,88 | 0,03 | |
| λ Колмогорова - Смирнова | 0,80 | 0,48 | 0,95 | 0,97 | 0,51 | |
| A = 0 | t Стьюдента | 0,44 | 0,06 | 0,58 | 0,38 | 0,12 |
| E = 0 | 0,14 | 0,01 | 0,58 | 0,05 | 0,81 | |
| M = 0 | 0,70 | 0,60 | 0,41 | 0,72 | 0,90 | |
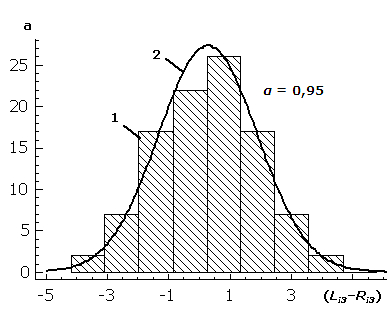
Рис. 2. Эмпирические частоты распределения (1) показателя асимметрии (Li3 – Ri3) третьего признака листа березы повислой выборки n05 и кривая нормального распределения с теми же параметрами (2) (указан уровень значимости соответствия нормальному закону по критерию χ2 Пирсона)
Fig. 2. The empirical frequency of the distribution (1) of asymmetry index (Li3 – Ri3) of the third sign of a silver birch leaf in the sample n05 and normal distribution curve with the same parameters (2) (the significance level of conformity to the normal distribution according to χ2 Pearson is noted)
Критерий W Шапиро – Уилкса показал статистически значимые отличия распределения разности сторон признаков от нормального закона в трех случаях (например, в табл. 2 для j = 5 уровень значимости 0,03<0,05), однако по остальным критериям достоверных отличий выявлено не было. Это подтверждает избыточную чувствительность данного метода (Palmer, Strobeck, 2003), и актуальность использования нескольких алгоритмов при анализе флуктуирующего характера асимметрии.)
Уровень асимметрии березы повислой разных возрастных состояний
Установлен тренд изменчивости показателей асимметрии по каждому признаку faj(1), faj(2) и по всем признакам FA(2) в зависимости от стадии возрастного состояния березы (рис. 3, нумерация признаков соответствует рис. 1).
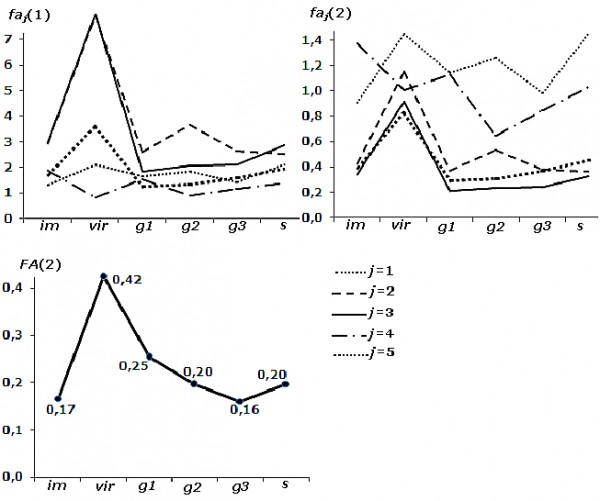
Рис. 3. Изменение показателей асимметрии по каждому признаку faj и интегрального индекса FA в зависимости от стадии онтогенеза березы повислой
Fig. 3. Changes in asymmetry indicators for each sign faj and for the integral index FA depending on the age-related state of the silver birch
Первый показатель использует именованные величины – разность исходных промеров признаков для оценки асимметрии. Никаких методов унификации показателей асимметрии признаков не проводится, следовательно, объединение faij(1) для расчета интегральных индексов – оценки асимметрии для одной особи по комплексу признаков (fai) и для выборки особей по всем признакам (FA) – невозможно (см. табл. 1). Оценка асимметрии, использующая нормированное отклонение t = (x – M) / S, адекватно оценивает дисперсию разностей исходных промеров между сторонами, подчиняется нормальному закону (анализ соответствия распределения всех выборок показателей faij(2) и fai(2) проводился с использованием критериев, представленных в табл. 2), снимает проблему «масштабного эффекта» (корреляцию между размером признака и величиной асимметрии) и позволяет объединять оценки асимметрии отдельных признаков (Зорина, Коросов, 2009). Базовый способ унификации статистических данных сохраняет различия в проявлении признака на левой и правой сторонах, а применение постоянных значений Mj и Sj при нормировании промеров позволяет сохранить исходную градацию величины признаков в сравниваемых группах.
В соответствии с первым показателем диапазон изменчивости величины асимметрии наибольший у признаков с максимальными размерами (j = 2 и 3, рис. 3). Нормирование, применяемое во второй оценке, позволило привести признаки к сопоставимому размаху варьирования (см. рис. 3). Тем не менее результаты, полученные при использовании обоих показателей faj(1) и faj(2), выявляют отличия в трендах изменчивости уровня асимметрии у четвертого и пятого признаков по сравнению с первыми тремя, что свидетельствует о разной реакции свойств листовой пластинки на изменение стадий онтогенеза деревьев.
В то же время период виргинильного возрастного состояния березы повислой характеризуется максимальным уровнем асимметрии для четырех признаков из пяти. Показатель асимметрии четвертого свойства (j = 4) максимален для имматурных деревьев. Минимальное значение отличается для каждого признака (но совпадает по показателям): для j = 1 и 3 минимум проявляется у генеративных молодых (g1) особей, для j = 2 – постгенеративных сенильных, для j = 4 – средневозрастных (g2), для j = 5 – имматурных (im). Тренд изменчивости интегрального индекса (FA(2)), рассчитанного по всем пяти показателям асимметрии признаков, отражает тенденцию, характерную для большинства свойств. Максимальное значение FA(2) проявляется у виргинильных (vir) деревьев, минимальное – у генеративных старых (g3) особей (см. рис. 3).
Проведена оценка достоверности влияния фактора «возраст» на величину асимметрии путем сравнения нескольких выборок по изменчивости признака с помощью критериев Кокрена, Бартлетта и Левена (табл. 3).
Таблица 3. Уровень значимости отличия выборок шести возрастных состояний березы повислой по изменчивости показателей faij(2) и индексов fai(2) асимметрии признаков листа
Table 3. Significance level of the difference between six age-related states of the silver birch in accordance with indicators variability faij(2) and indexes fai(2) of leaf sign asymmetry
| Статистические критерии | fai1 | fai2 | fai3 | fai4 | fai5 | fai (по всем признакам) |
| Кокрен С | 0,00 | 0,00 | 0,00 | 0,02 | 0,34 | 0,00 |
| Бартлетта | 0,00 | 0,00 | 0,00 | 0,01 | 0,09 | 0,00 |
| Левена | 0,00 | 0,00 | 0,00 | 0,06 | 0,16 | 0,00 |
Результаты сравнительного анализа свидетельствуют о том, что изменение возрастного состояния березы повислой оказывает статистически значимое влияние на уровень асимметрии большинства признаков листа (за исключением j = 5, табл. 3).
Общие статистические критерии не показывают структуру варьирования, и достоверность отличия всех шести выборок может быть обусловлена изменением величины асимметрии только в период одного возрастного состояния по сравнению с другими стадиями онтогенеза. Анализ трендов изменчивости показателей (см. рис. 3) позволяет предположить, что виргинильная стадия обеспечивает значимое влияние возраста деревьев на уровень асимметрии большинства признаков. Это подтверждается результатами сравнительного анализа (табл. 4), проведенного при исключении выборки nvir. Достоверное воздействие фактора «возраст» на величину асимметрии подтверждается по всем трем критериям только для четвертого признака (см. табл. 4).
Таблица 4. Уровень значимости отличия выборок пяти возрастных состояний (без виргинильного) березы повислой по изменчивости показателей faij(2) и индексов fai(2) асимметрии признаков листа
Table 4. Significance level of the difference between five age-related states (without virginilnymi) of the silver birch in accordance with indicators variability faij(2) and indexes fai(2) of leaf sign asymmetry
| Статистические критерии | fai1 | fai2 | fai3 | fai4 | fai5 | fai (по всем признакам) |
| Кокрен С | 0,16 | 0,10 | 0,16 | 0,02 | 0,12 | 0,06 |
| Бартлетта | 0,19 | 0,30 | 0,07 | 0,00 | 0,14 | 0,15 |
| Левена | 0,06 | 0,43 | 0,02 | 0,03 | 0,47 | 0,43 |
Более информативным оказывается сопоставление изменчивости двух выборок с помощью обычных критериев (табл. 5). Так, результаты попарного сравнения выборок разных этапов онтогенеза деревьев для оценок асимметрии признаков j = 1, 2 и 3 показали статистически значимые отличия между nvir и другими выборками. Дополнительно для показателя асимметрии первого признака достоверные отличия обнаружены между ng1 и ns; для j = 3 – nim и ng1, ns и ng1; для j = 4 – nim и ng2, nim и ng3, nvir и ng2, ng1 и ng2, ns и ng2; для j = 5 – между nim и nvir, nim и ns (табл. 5). Интегральный индекс FA(2) объединяет данные, характерные для большинства признаков: величина асимметрии листа деревьев виргинильного (vir) возрастного состояния достоверно выше, чем других стадий онтогенеза, а генеративных молодых (g1) особей – чем имматурных (im) и генеративных старых (g3).
Таблица 5. Уровень значимости отличия выборок показателей асимметрии faij(2) методом попарного сравнения дисперсий по критерию F Фишера четвертого (верхний правый угол) и пятого (нижний левый угол) признаков листа березы повислой
Table 5. significance level of the difference of samples of asymmetry indicators faij(2) by the method of paired comparison of dispersion according to F Fisher criterion of the fourth (the upper right corner) and the fifth (the bottom left corner) signs of a silver birch leaf
| Стадии онтогенеза | fai4 | ||||||
| im | vir | g1 | g2 | g3 | s | ||
| fai5 | im | 1 | 0,12 | 0,33 | 0,00 | 0,02 | 0,15 |
| vir | 0,02 | 1 | 0,56 | 0,03 | 0,41 | 0,89 | |
| g1 | 0,25 | 0,24 | 1 | 0,01 | 0,16 | 0,65 | |
| g2 | 0,10 | 0,49 | 0,63 | 1 | 0,17 | 0,02 | |
| g3 | 0,70 | 0,053 | 0,44 | 0,21 | 1 | 0,34 | |
| s | 0,02 | 0,99 | 0,24 | 0,49 | 0,054 | 1 | |
Анализ тренда изменчивости показателей асимметрии признаков листа и достоверности отличий между выборками разных стадий онтогенеза березы повислой позволяет нам разделить пять исследуемых свойств на три группы: в первую входят j = 1, 2 и 3, во вторую – j = 4, в третью – j = 5. В данном случае затрагивается проблема выбора модели формирования флуктуирующей асимметрии в процессе развития отдельной особи с учетом неоднородной реакции ее характеристик на изменение возрастного состояния.
Результаты классификации признаков методом кластерного анализа (рис. 4) показывают группу свойств, наиболее близких по степени их реакции на онтогенетическое изменение березы – j = 1, 3 и 2. Обособленное положение занимают четвертый и пятый признаки (см. рис. 4).
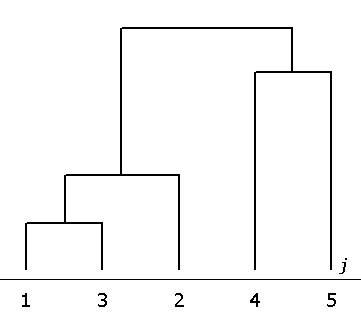
Рис. 4. Дендрограмма сходства признаков по степени влияния на показатель faj(2) асимметрии возраста деревьев (метод ближайшего соседа, Евклидова мера расстояния)
Fig. 4. The dendrogram of sign similarities according to the extend of the influence of tree age on faj(2) asymmetry indicators (method of the nearest neighbour, Euclidean distance measure)
Анализ изменчивости асимметрии отдельных признаков в зависимости от стадии онтогенеза деревьев и их кластеризация позволяют пересмотреть процедуру расчета интегрального индекса FA(2). Несмотря на то что все изучаемые признаки имеют общие характеристики – пластические (метрические), вегетативные, только одного органа (листовой пластинки) – объединять их в один индекс оказывается нецелесообразным. С одной стороны, разные тренды изменчивости могут гасить друг друга, что приведет к неопределенному результату, с другой – изменчивость FA может определяться влиянием нескольких доминантных признаков, одинаково реагирующих на изменение фактора. В обоих случаях интегральный показатель будет некорректно характеризовать изменение уровня асимметрии в зависимости от стадии возрастного состояния березы и не учтет разнообразие вариантов ее формирования в процессе развития особей.
Проведенный анализ изменчивости асимметрии признаков позволяет нам из пяти исходных свойств листовой пластинки сформировать три оценки с разной реакцией на индивидуальное развитие березы: fa4(2), fa5(2) и интегральный показатель FA1,2,3(2), рассчитанный на основе fai1(2), fai2(2), fai3(2) (рис. 5).
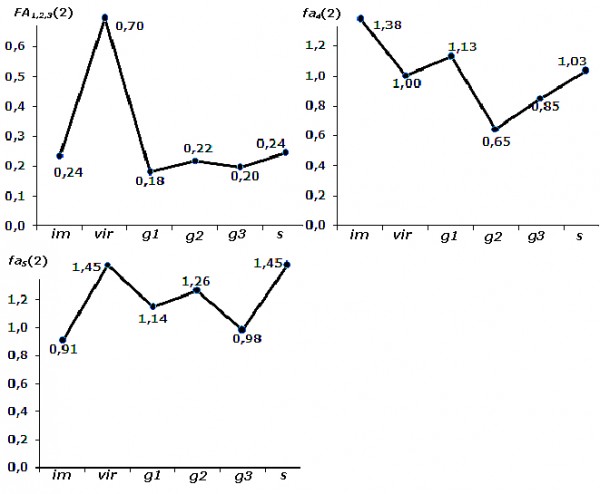
Рис. 5. Изменчивость трех значимых (пояснение в тексте) оценок асимметрии в зависимости от стадии онтогенеза березы повислой
Fig. 5. Variability of three significant (explanation is in the text) estimates of asymmetry depending on the ontogenesis stage of the silver birch
Для пересчитанного интегрального индекса FA1,2,3(2) результаты попарного сравнения выборок разных стадий онтогенеза деревьев показали статистически значимые отличия только между nvir и всеми другими выборками. Достоверные изменения показателей fa4(2) и fa5(2) в зависимости от возраста березы повислой представлены в табл. 5.
Изменение величины асимметрии листовой пластинки в течение морфогенеза
Результаты, полученные при изучении изменчивости оценок асимметрии березы повислой в течение одного вегетационного периода, показывают разные тренды ее динамики у исследуемых признаков и статистически значимые отличия между соответствующими выборками разных стадий морфогенеза листа (рис. 6, табл. 6).
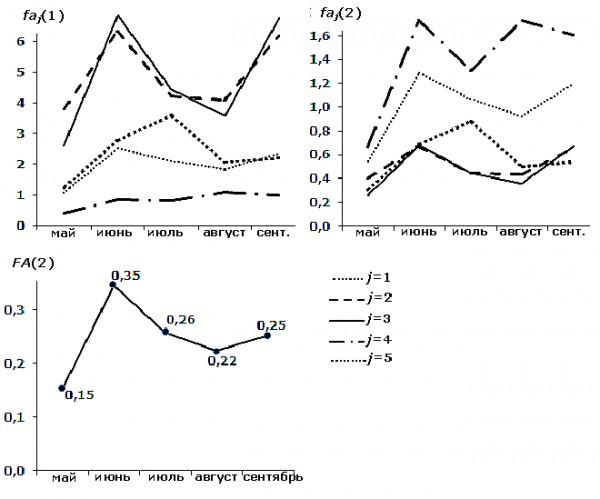
Рис. 6. Изменчивость показателей асимметрии по каждому признаку faj и интегрального индекса FA в зависимости от стадии морфогенеза листа (по месяцам вегетационного периода года)
Fig. 6. Variability of asymmetry indicators for each sign faj and the integral index FA depending on the morphogenesis stage of a leaf (according to the months of vegetation period)
Таблица 6. Уровень значимости отличия выборок по изменчивости показателей faij(2) и индексов fai(2) асимметрии признаков листа разных стадий морфогенеза у березы повислой (метод попарного сравнения дисперсий по критерию F Фишера)
Table 6. Significance level of the difference of samples in accordance with variability indicators faij(2) and fai(2) indexes of silver birch leaf signs asymmetry at different morphogenesis stages (method of paired comparison of dispersion according to F Fisher criterion)
| Сравниваемые выборки | fai1 | fai2 | fai3 | fai4 | fai5 | fai (по всем признакам) | fai по трем признакам: j = 1, 2 и 3 |
| n05 и n06 | 0,00 | 0,01 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 |
| n05 и n07 | 0,00 | 0,63 | 0,01 | 0,00 | 0,00 | 0,01 | 0,01 |
| n05 и n08 | 0,01 | 0,74 | 0,12 | 0,00 | 0,01 | 0,06 | 0,30 |
| n05 и n09 | 0,00 | 0,02 | 0,00 | 0,00 | 0,00 | 0,01 | 0,04 |
| n06 и n07 | 0,22 | 0,04 | 0,03 | 0,17 | 0,36 | 0,15 | 0,45 |
| n06 и n08 | 0,13 | 0,03 | 0,00 | 0,99 | 0,10 | 0,03 | 0,03 |
| n06 и n09 | 0,26 | 0,90 | 0,94 | 0,72 | 0,73 | 0,12 | 0,24 |
| n07 и n08 | 0,01 | 0,88 | 0,27 | 0,17 | 0,47 | 0,48 | 0,15 |
| n07 и n09 | 0,02 | 0,06 | 0,04 | 0,31 | 0,56 | 0,91 | 0,68 |
| n08 и n09 | 0,69 | 0,04 | 0,00 | 0,74 | 0,19 | 0,56 | 0,30 |
Интегральный индекс FA(2), рассчитанный на основе нормированных показателей асимметрии пяти признаков (см. табл. 1), учитывает тренды изменчивости уровня асимметрии большинства из них. Например, показатели асимметрии листовой пластинки в мае достоверно ниже, чем в другие месяцы вегетационного периода года отдельно по каждому признаку, так же как и по оценке асимметрии с учетом всех признаков (см. табл. 6). Однако статистически значимые отличия между выборками n05 и n08 при расчете FA(2) не проявляются, т. к. разнонаправленные тренды изменчивости оценок асимметрии для признаков j = 4 и j = 5 выборки n08 гасят друг друга.
Анализ направлений изменчивости показателей асимметрии (см. рис. 6) и результатов попарного сравнения выборок разных стадий морфогенеза (см. табл. 6) показывает, что индекс FA(2) в большей мере учитывает результаты первых трех признаков. Это также подтверждает расчет индексов fai(1,2,3)(2) и FA1,2,3(2) на основе показателей асимметрии этих свойств (j = 1, 2 и 3, табл. 6) – достоверные отличия между соответствующими выборками по fai(1,2,3) (2) сохраняются. Тем не менее при использовании общих статистических критериев влияние фактора «стадия морфогенеза листа березы повислой» на величину асимметрии статистически значимо для показателей всех признаков, как по отдельности, так и в совокупности (как минимум по двум критериям из трех, табл. 7).
Таблица 7. Уровень значимости отличия выборок пяти стадий морфогенеза листа по изменчивости показателей faij(2) и индексов FA(2) признаков листа березы повислой
Table 7. Significance level of the difference of five leaf morphogenesis stages samples in accordance with indicators variability faij(2) and FA(2) indexes of silver birch leaf signs
| Статистические критерии | fai1 | fai2 | fai3 | fai4 | fai5 | fai (все признаки) | fai по трем признакам: j = 1, 2 и 3 |
| Кокрен С | 0,00 | 0,08 | 0,01 | 0,20 | 0,08 | 0,01 | 0,04 |
| Бартлетта | 0,00 | 0,02 | 0,00 | 0,00 | 0,00 | 0,00 | 0,01 |
| Левена | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,03 | 0,01 |
Результаты классификации признаков методом кластерного анализа (рис. 7) показывают группу свойств, наиболее близких по степени реакции величины их асимметрии на изменение морфогенетического состояния листа березы повислой – j = 1, 3 и 2. Обособленное положение занимают четвертый и пятый признаки. В целом данные результаты подтверждают полученную ранее дендрограмму сходства признаков по степени влияния на уровень их асимметрии возраста деревьев (см. рис. 4).
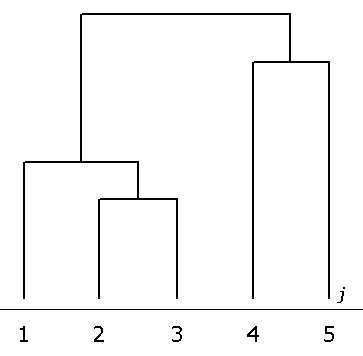
Рис. 7. Дендрограмма сходства признаков по степени влияния на показатель faj(2) асимметрии стадии морфогенеза листа деревьев (метод ближайшего соседа, Евклидова мера расстояния)
Fig. 7. The dendrogram of similarity of the signs according to the extend of the influence of leaf morphogenesis stage on faj(2) asymmetry indicator (method of the nearest neighbour, Euclidean distance measure)
В обоих случаях – при изучении влияния возраста деревьев и стадии морфогенеза листа на уровень асимметрии – подтверждена целесообразность формирования трех оценок асимметрии из пяти исходных показателей: fa4(2), fa5(2) и FA1,2,3(2) (рис. 8). Это важно для дальнейшего теоретического анализа закономерностей динамики асимметрии в процессе индивидуального развития березы повислой.
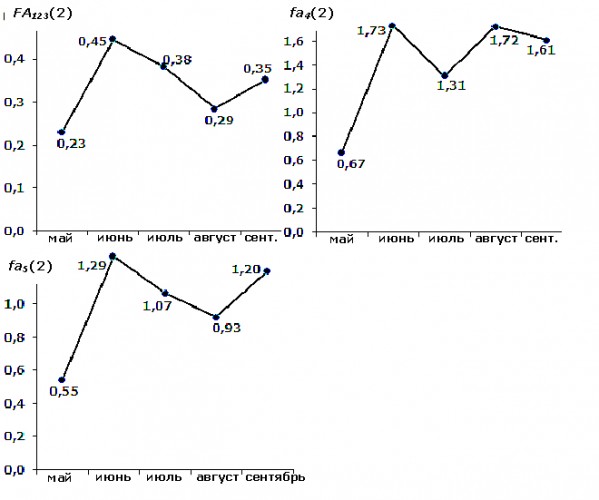
Рис. 8. Изменчивость трех значимых (пояснение в тексте) оценок асимметрии в зависимости от стадии морфогенеза листа березы повислой
Fig. 8. Variability of three significant (explanation is in the text) estimates of asymmetry depending on the leaf morphogenesis stage of the silver birch
Обсуждение
Онтогенетические состояния особей и стадии морфогенеза их органов достоверно влияют на уровень флуктуирующей асимметрии метрических, вегетативных признаков листовой пластинки березы повислой. Однако общей закономерности, единого тренда изменчивости оценок асимметрии для всех пяти исследованных характеристик деревьев установлено не было, несмотря на проявление некоторых общих тенденций (например, минимальный уровень асимметрии наблюдается на начальной стадии морфогенеза листа у всех рассмотренных признаков, см. рис. 8).
В современной литературе выделяют по меньшей мере семь моделей формирования асимметрии в процессе развития отдельных особей и их органов (Kellner, Alford, 2003; рис. 9).
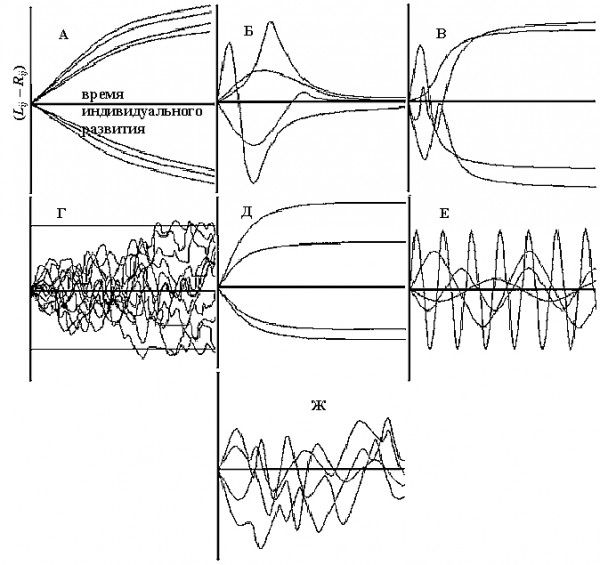
Рис. 9. Гипотетические модели формирования флуктуирующей асимметрии в процессе развития отдельных особей и их органов: А – модель направленных внешних сигналов; Б – бросания монеты; В – увеличения асимметрии; Г – накопления случайностей; Д – постоянной (стабильной) асимметрии; Е – компенсаторного роста или изменчивости; Ж – остаточной асимметрии
Fig. 9. Hypothetical models of the formation of fluctuating asymmetry during the development process of individuals and their parts. Models of: A – directed external signals; B – tossing coins; C – increasing asymmetry; D – accumulation of accidents; E – constant (stable) asymmetry; F – compensatory growth or variatbility; G – residual asymmetry
В рамках первой модели предполагается, что величина асимметрии определяется односторонним воздействием факторов как экзогенного (например, свет), так и эндогенного (отсутствие пальца) происхождения. В процессе развития особи прирост величины признака больше на той стороне тела, которая подвержена влиянию фактора, поэтому уровень асимметрии увеличивается параллельно с увеличением признака (см. рис. 9А).
Если морфогенез состоит из отдельных этапов (онтогенетических единиц), независимо проявляющихся в процессе индивидуального развития организма, то говорят о модели «бросания монеты». В пределах каждого этапа развития направление и величина асимметрии генетически детерминированы, но асимметрия одного этапа относительно другого задана случайно. Более того, на ранних стадиях развития особи изменчивость асимметрии больше, чем на поздних (см. рис. 9Б).
Когда онтогенетические единицы взаимозависимы, то малые ненаправленные отличия в раннем возрасте увеличиваются в течение морфогенеза. Прирост признака постоянно больше на одной стороне тела. Источником асимметрии могут быть небольшие отличия физико-химического состояния цитоплазмы клеток на двух сторонах тела и случайные отличия в скорости деления клеток. Так как асимметрия у каждой особи возникает случайно, то на популяционном уровне она направленности не имеет. Направление асимметрии в процессе развития особи остается постоянным, а величина возрастает (см. рис. 9В).
В модели «накопления случайностей» считается, что ни один из биологических объектов не развивается по идеальному пути, и онтогенетический шум накапливается с течением времени. Поэтому время развития признака положительно коррелирует с величиной флуктуирующей асимметрии и в процессе морфогенеза изменчивость асимметрии будет увеличиваться до определенного уровня, а затем может оставаться постоянной (рис. 9Г).
Теоретическая основа модели «постоянной (стабильной) асимметрии» в том, что направление и величина флуктуирующей асимметрии определяются генетическими или средовыми факторами на ранних этапах онтогенеза и в течение остальных стадий развития не изменяются (остаются постоянными). Но уровень асимметрии определяется для каждого признака отдельно и возможно на разных стадиях онтогенеза (рис. 9Д).
Отсутствие больших случайных отличий в проявлении признаков на разных сторонах тела организма наводит на мысль о существовании регуляторных механизмов обратной связи. В модели «компенсаторного роста» предполагается, что асимметрия возникает случайным образом и изменение ее величины находится под контролем системы обратных связей, стимулирующей компенсаторный рост на обеих сторонах. Выделяют два механизма регуляции величины асимметрии: 1 – система отрицательных обратных связей между клетками угнетает биосинтез на той стороне тела, которая начинает развиваться более интенсивно; 2 – система положительных обратных связей (реализующаяся через нервную и гуморальную системы) между сторонами тела обеспечивает интенсивный прирост отстающей стороны. Это может привести к временным колебаниям уровня асимметрии, и ее величину можно будет выразить функцией от индивидуальной скорости роста, времени реагирования систем положительных обратных связей, силы ростовых сигналов (рис. 9Е).
J. H. Graham (1993) предложил теоретическую модель нелинейной обратной связи, которая регулирует формирование асимметрии морфогенами, способными к распространению. По его мнению, обратная связь может реализовываться как минимум тремя способами: 1 – прямая взаимосвязь между лево- и правосторонними элементами билатерального признака; 2 – опосредованная связь, когда передача информации между сторонами происходит через центральную нервную или гуморальную системы; 3 – действуют как прямая, так и опосредованная обратные связи. Высока вероятность того, что относительная активность этих обратных связей в процессе развития особи будет меняться и разные стадии будут характеризоваться разной чувствительностью сторон к регуляторным компонентам. Более того, подобные отличия могут быть и между признаками одной и той же особи.
В настоящее время в онтогенетической биологии билатеральная симметрия как таковая и отклонения от нее обычно остаются без внимания, так как принимаются за побочный продукт моделей формирования. Это явление просто выходит за рамки обычных правил: нет специализированных онтогенетических механизмов, которые бы отвечали за формирование билатеральной симметрии самой по себе. Направление и этап развития клетки определяются ее локализацией в трехмерном пространстве. При этом левая и правая стороны тела назначаются после установления первых двух осей: передне-задней и спинно-брюшной. Однако третья ось – это не лево-правая ось сама по себе, а скорее проксимо-дистальная: клетки определенной стороны тела принимают онтогенетическое решение на основе их положения по отношению к средней линии. Так как развитие клетки происходит в трех этих осях, то формирование билатерально симметричных структур – это неизбежный результат. С флуктуирующей асимметрией связана одна из важных проблем современной биологии – упорядоченное выражение генотипа как комплексного трехмерно-пространственного фенотипа.
Последняя модель «остаточной асимметрии», предложенная J. R. Kellner и R. A. Alford (2003), представляет собой один из вариантов «компенсаторного роста», в котором вероятность регулярных колебаний величины асимметрии при онтогенезе особи сведена до минимума. Существует простой компенсаторный механизм, отвечающий за морфологию особи с относительно короткой временной задержкой. В результате формируются слабые и непродолжительные временные колебания величины асимметрии признака в процессе развития особи (см. рис. 9Ж). Уровень асимметрии каждой особи в конкретный временной промежуток представляет собой разницу между онтогенетическим шумом и его коррекцией.
Результаты проведенного нами исследования наиболее сопоставимы с моделью «компенсаторного роста или изменчивости» по сравнению с другими гипотезами формирования асимметрии в процессе развития отдельных особей и их органов (табл. 8).
Таблица 8. Сравнение моделей формирования асимметрии в процессе морфо- и онтогенеза с результатами исследования (буквенное обозначение соответствует рис. 9)
Table 8. Comparison of asymmetry formation models during morpho- and ontogenesis with the results of the study ( letter symbols correspond to fig. 9)
| Характеристика | Модель | Результаты исследования | ||||||
| А | Б | В | Г | Д | Е | Ж | ||
| Уровень асимметрии устанавливается в раннем онто(морфо)генезе и со временем не меняется | + | ─ | + | ─ | + | ─ | ─ | ─ |
| Время развития признака положительно коррелирует с величиной асимметрии | ─ | ─ | ─ | + | ─ | ─ | ─ | ─ |
| Время развития признака отрицательно коррелирует с величиной асимметрии | ─ | + | ─ | ─ | ─ | ─ | ─ | ─ |
| Уровень асимметрии меньше у крупных признаков по сравнению с мелкими | ─ | + | ─ | ─ | ─ | ─ | ─ | ─ |
| Величина асимметрии отражает текущее состояние особей и популяции в целом | ─ | ─ | ─ | ─ | ─ | + | + | + |
| Отличия показателей асимметрии признаков между разными стадиями развития особей статистически значимы | + | + | + | + | + | + | ─ | + |
| Тренды изменчивости асимметрии в процессе онто(морфо)генеза разных признаков могут отличаться | ─ | ─ | ─ | ─ | ─ | + | + | + |
Основное отличие между этими теоретическими построениями при экстраполяции на популяционный уровень заключается в ответе на вопрос о том, какую часть истории развития популяции отражает уровень флуктуирующей асимметрии. Первая, третья и пятая модели предполагают, что текущий уровень асимметрии отражает кратковременные случайные воздействия, которые оказывали влияние на особей в раннем онтогенезе. В случае их справедливости оценка популяционного уровня асимметрии будет характеризовать состояние популяции в прошлом, время которого определяется возрастом генерации, участвующей в исследовании. Четвертая модель предполагает, что одна и та же величина асимметрии может сформироваться под действием двух разных процессов: кратковременного интенсивного отрицательного воздействия на особь факторов среды или слабого влияния стресс-агентов на протяжении длительного периода времени в течение развития особи. В рамках последних двух предположений принимается то, что величина асимметрии в любое время отражает только текущее состояние особей и популяции в целом, тогда как вторая модель говорит об уменьшении асимметрии с увеличением размера тела при любой степени отрицательного воздействия факторов.
Проведенные исследования для выявления механизма формирования асимметрии редко тестируют все перечисленные модели, подтверждают иногда справедливость сразу нескольких из них, допускают, что модели изменения асимметрии у разных видов, также как и у разных признаков, могут отличаться (Moller, 1996; Swaddle, Witter, 1997; Collin, 1997; Kellner, Alford, 2003). Например, A. K. Chippendale и A. R. Palmer (1993) при изучении изменчивости уровня асимметрии сегментных конечностей краба Hemigrapsus nudus показали, что в процессе линьки особей (стадия интенсивного роста) ее показатели остаются постоянными. Это, по их мнению, подтвердило модель постоянной (стабильной) асимметрии, и они сделали заключение, что величина асимметрии генетически детерминирована. В то же самое время на дрозофиле они получили противоположные результаты, для объяснения которых ими была введена гипотеза «порогового уровня». Предполагается, что существует некоторая пороговая величина онтогенетического шума (отражающего средовое воздействие), ниже которого асимметрия остается неизменной в процессе развития. Более того, выбранные признаки не могли совпадать по функциональной значимости, что, по их мнению, также повлияло на отличие результатов по разным видам. Данные A. P. Moller (1996) по Hirundo rustica (хвостовые признаки) подтверждают сразу две модели – увеличения асимметрии и постоянной (стабильной) асимметрии; результаты исследования R. E. Young (1994) по Alpheus heterochelis свидетельствуют в пользу модели компенсаторной изменчивости и т. д.
Отсутствие согласованной изменчивости величины асимметрии разных признаков объясняется уникальностью комплекса процессов онтогенетического шума, стабильности развития и их взаимодействия для каждого конкретного признака. Единство генотипа не простирается на единство систем контроля развития. Разные признаки могут по-разному реагировать на одно и то же воздействие фактора, то есть обладать разной чувствительностью к воздействиям эндо- и экзогенных условий. У каждого свойства будет своя особая система репарации, своя способность к пластичности и стабильности. Так как случайная изменчивость развития находится в пределах нормы реакции генотипа и процессы, обладающие эффектом буфера, генетически детерминированы, то различия в асимметрии признаков будут связаны с особенностями реализации генотипа в процессе развития фенотипа (например, разные начала и скорости развития признаков). Более того, у каждой черты организма своя история развития (становление в процессе филогенеза) и набор сил направленной и стабилизирующей селекции, что тоже обуславливает различия трендов изменчивости показателей асимметрии разных признаков особей одного вида под влиянием изменяющихся факторов среды (Leung, Forbes, 1997).
Заключение
Уровень флуктуирующей асимметрии отражает результат взаимодействия онтогенетического шума (компоненты случайности, препятствующей реализации идеала в виде совершенной симметрии) и стабильности развития (системы репарации и корректировки отклонений от априорного состояния), достигнутый в течение онтогенеза особи (Захаров, 2001; Рalmer, Strobeck, 1992; Mitton, 1993; Рalmer, 1994; Зорина, 2010 и др.). Оба явления могут оказывать влияние на направление изменчивости показателей асимметрии и ее величину в конкретный момент времени. При этом считается, что чем больше уровень асимметрии, тем интенсивнее было воздействие, вызвавшее увеличение «шума», или тем слабее была буферная способность процессов развития по предотвращению вызванных изменений (Developmental…, 2003).
Изучение онто- и морфогенетической изменчивости показателей асимметрии листовой пластинки березы повислой позволило сформулировать следующие выводы:
- Уровень асимметрии листа зависит от стадии онтогенеза (возрастного состояния) березы повислой.
- Уровень асимметрии листа зависит от стадии его морфогенеза.
- Тренды изменчивости величины асимметрии в течение индивидуального развития особей достоверно отличаются у разных групп признаков. Так, для листа березы повислой пять исследуемых признаков классифицируются на три группы: первая включает j = 1, 2, 3, вторая – j = 4, третья – j = 5.
- Изменение показателей асимметрии в процессе онто(морфо)генеза березы повислой подтверждает модель «компенсаторного роста или изменчивости».
Установленные закономерности формирования уровня флуктуирующей асимметрии в течение индивидуального развития требуют подтверждения или опровержения при изучении описанного явления с использованием других биологических объектов. До сих пор многие прикладные и теоретические вопросы данного направления исследования остаются нерешенными.
Библиография
Гелашвили Д. Б., Якимов В. Н., Логинов В. В., Епланова Г. В. Статистический анализ флуктуирующей асимметрии билатеральных признаков разноцветной ящурки // Актуальные проблемы герпетологии и токсинологии: Сборник научных трудов. Тольятти, 2004. Вып. 7. С. 45—59.
Захаров В. М. Онтогенез и популяция (стабильность развития и популяционная изменчивость) // Экология. 2001. № 3. С. 164—168.
Зорина А. А. Методы статистического анализа флуктуирующей асимметрии // Принципы экологии. 2012. № 3. С. 24—47.
Зорина А. А. Оценка флуктуирующей асимметрии на основе нормированного отклонения // Принципы экологии. 2013. Т. 2. № 1. С. 69—72.
Зорина А. А. Флуктуирующая асимметрия животных и растений . Saarbrucken: Lap Lambert Academic Publishing GmbH & Co.KG, 2010. 180 с.
Зорина А. А., Коросов А. В. Изменчивость показателей и индексов асимметрии признаков листа в кроне Вetula pendula (Betulaceae) // Ботанический журнал. 2009. Т. 94. № 8. С. 1172—1192.
Коросов А. В., Коросов А. А. Техника ведения ГИС: Приложение в экологии . Петрозаводск: Изд-во ПетрГУ, 2006. 186 с.
Лантратова А. С., Сонина А. В. Экология растений . Ч. II. Петрозаводск: Изд-во ПетрГУ, 2005. 180 с.
Методические рекомендации по выполнению оценки качества среды по состоянию живых существ (оценка стабильности развития живых организмов по уровню асимметрии морфологических структур) . Распоряжение Росэкология от 16.10.2003. № 460–р. М., 2003. 28 с.
Cheverud J. M., Leamy L., Atchley W. R., Rutledge J. J. Quantitative genetics and the evolution of ontogeny. I. Ontogenetic changes in quantitative genetic variance components in randombred mice // Genetical Res. 1983. № 42. P. 65—75.
Collin R. Ontogeny of subtle skeletal asymmetries in individual larvae of the sand dollar Dendraster excentricus // Evolution. 1997. № 51. P. 999—1005.
Developmental instability: causes and consequences / ed. M. Polak. N. Y.: Oxford Univ. Press, 2003. 500 p.
Graham J. H., Freeman D. C., Emlen J. M. Antisymmetry, directional asymmetry, and dynamic morpogenesis // Genetica. 1993. № 89. P. 121—137.
Kellner J. R., Alford R. A. The ontogeny of fluctuating asymmetry // Amer. Natur. 2003. Vol. 161. N 6. P. 931—947.
Kimball R. T., Ligon J. D., Merola–Zwartjes M. Fluctuating asymmetry in red jungle fowl // Journal of Evolutionary Biology. 1997. № 10. P. 441—457.
Leung B., Forbes M. R. Modelling fluctuating asymmetry in relation to stress and fitness // Oikos. 1997. № 78. P. 397—405.
Mitton J. B. Enzyme heterozygosity, metabolism and developmental stability // Genetica. 1993. № 89. P. 47—65.
Moller A. P. Development of fluctuating asymmetry in tail feathers of the barn swallow Hirunda rustica // Journal of Evolutionary Biology. 1996. № 9. P. 677—694.
Palmer A. R., Strobeck C. Fluctuating asymmetry analyses revisited // Developmental instability: causes and consequences. N.Y.: Oxford Univ. Press, 2003. P. 279—319.
Swaddle J. P., Witter M. S. On the ontogeny of developmental stability in a stabilized trait // Proc. Roy. Soc. London. 1997. № 264. P. 329—334.
Young R. E., Pearce J., Govind C. K. Establishment and maintenance of claw bilateral asymmetry in snapping shrimps // J. exp. Biol. 1994. № 269. P. 319—326.
Благодарности
Автор благодарит Е. А. Петрову за помощь в сборе исходного материала.

 © 2011 - 2026
© 2011 - 2026