



Введение
Литораль – это экотон трех сред: лито-, гидро- и атмосферы. Как экотоп она трехмерна: донная часть ее имеет длину вдоль линии прилива или отлива, ширину (между вдольбереговой границей (перпендикулярно ей) и линией уреза воды в отлив) и толщину, состоящую из обитаемого слоя грунта и слоя воды над дном (глубину во время прилива). Этот трехмерный экотоп предоставляет гидробионтам Белого моря различные условия, значительно более резко колеблющиеся (в суточном, сезонном и межгодовом режимах), чем в бентали, расположенной ниже нуля глубин (сублиторали и псевдобатиали). Тем не менее литоральный бентос высокопродуктивен и играет важную роль в трофической составляющей экосистемы Белого моря в целом. Систематическое изучение Белого моря ведется со второй половины XVIII столетия. Особенно в последнее время опубликовано множество работ, посвященных исследованиям бентоса Белого моря, а также конкретно Онежского и Кандалакшского его заливов, например тематический сборник «Экосистемы Онежского залива Белого моря» (1985); монографии «Гидробиокомплексы Белого моря» (1984), «Белое море. Биологические ресурсы и проблемы их рационального использования» (1995); масса отдельных статей и монографий, а также диссертаций и многое другое. Однако исследования, посвященные литоральному бентосу, весьма фрагментарны и далеко не достаточны.
Объектом наших исследований выбран макрозообентос литорали двух самых крупных заливов Белого моря исходя из современной его актуальности как важной мелководной трофической и в целом биотической составляющей экосистем Белого моря.
Цель работы заключалась в исследовании распределения различных систематических групп литорального зообентоса в двух самых крупных районах Белого моря – Кандалакшском и Онежском заливах с анализом биохорологической иерархической структуры фаунистических комплексов.
Материалы
Проводилось обследование литорали материковой части и островов Белого моря. По климатическим условиям исследованные нами районы не могут иметь существенных различий, так как в географическом широтном направлении они отстоят всего примерно на 2º друг от друга. Специфические черты биологического облика Кандалакшского и Онежского заливов определяются гидрологическими условиями, обусловленными геоморфологическим их строением, которые и создают довольно существенные отличия условий обитания литоральных беспозвоночных в этих двух крупнейших акваториях Белого моря.
Открытая часть Кандалакшского залива по геологическому строению идентична с центральной частью моря, называемой Бассейном. Здесь берет начало Кандалакшский глубоководный желоб, пролегающий вдоль всего залива до самой его вершины. В Кандалакшском заливе (у Турьего мыса) располагается самая большая глубина всего моря – 340 м, его средняя глубина составляет 109 м.
Бассейн – это гигантская ковшевая и глубоководная (средняя глубина 125 м) часть Белого моря, в которой содержится около половины всей водной массы. Течения, преобладающие здесь, направлены по концентрическим окружностям. Бассейн разделяет соседствующие Кандалакшский и Онежский заливы (рис. 1). Геологическое строение и гидрологический режим Бассейна обеспечивают своеобразие и относительную самостоятельность литоральной фауны исследованных акваторий. Онежский залив почти в 2 раза больше Кандалакшского по площади, но значительно более мелководный, средняя его глубина составляет 19 м, максимальная – 87 м. Солнечная радиация в Белом море проникает до глубины не более чем 25–30 м, поэтому в Онежском заливе в летний период водные массы прогреваются до 15 ºС (Бабков, 1985). Вследствие мелководности, а также приливно-отливных и ветроволновых процессов водные массы перемешиваются, образуя гомогенные термогалинные условия для всей населяющей этот залив биоты, в том числе и литоральной. В Кандалакшском заливе водные массы имеют контрастную стратифицированность – у дна температура всегда отрицательная (от -0,1 до -1,3 ºС) и соленость до 30 ‰ (Бабков, Голиков, 1984); у поверхности летом – всегда положительная (около +9–12 ºС, зимой до -1,7 ºС). Соленость поверхностных слоев воды в разных местах акватории от 5 до 27–29 ‰ (Бабков, Голиков, 1984). В результате непрерывно происходящих приливно-отливных, а в летнее время и ветроволновых процессов отдельные потоки и струи холодных глубинных вод подходят к поверхности, особенно в отдельных местах (апвеллинг).
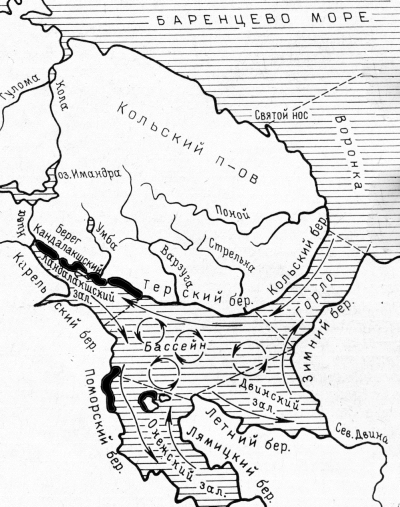
Рис. 1. Картосхема Белого моря (Наумов, Оленев, 1981). Стрелками показана циркуляция вод, черными полосами –места сбора материалов
Fig. 1. Schematic map of the White Sea (Naumov, Olenev, 1981). The arrows indicate the water circulation, black stripes – places of material collection
В 2000–2010 гг. была обследована литораль материковой части Кандалакшского залива (Лувеньга, Порья Губа, Турий мыс) и ряда крупных островов: Олений, Большой Березовый, Лодейный, Вороний, Ряшков, Девичья луда. В 2010–2011 гг. литоральный бентос изучался в Онежском заливе: на материковой части в районе поселка Колежма и на островной – на Большом Соловецком острове (мыс Толстик и губа Долгая).
Методы
Качественные учеты и сборы беспозвоночных проводились маршрутным способом вдоль литоральной полосы. Через каждые 100–200 м от 0 глубин до супралиторали поперек литоральной полосы трансектным способом проводились описания и сборы беспозвоночных. Для исследования эпифауны беспозвоночные собирались с поверхности всех типов грунтов и водорослей. Для изучения обитателей литоральной инфауны грунт выбирался на глубине 30–40 см, собранные пробы промывались через сито с ячеей 1 мм2; верхние слои грунта промывались через более мелкое сито с размером ячеи 0,7 мм2 с целью обнаружения ювенильных стадий организмов некоторых видов, не имеющих равномерного распространения, таких, например, как Mya arenaria, Acanthogammarus loricatus и некоторых других. В случае отсутствия взрослых форм таких беспозвоночных эти виды вносились в списки обнаруженных по их легко идентифицируемым молодым особям.
Всего было выполнено в Кандалакшском заливе 200, в Онежском – 189 разрезов.
Идентификационная, систематическая и биогеографическая аналитическая работа проводилась по различным источникам: Определитель…, 1948; Колтун, 1959; Клюге, 1962; Цветкова, 1975; Цетлин, 1980; Моллюски…, 1987; Малахов, Адрианов, 1999; Иллюстрированный…, 2006; Каталог…, 2008; Флора…, 2010).
Результаты
В результате составлен список всех беспозвоночных, обнаруженных на литорали Кандалакшского и Онежского заливов (табл. 1).
Таблица 1. Аннотированный список беспозвоночных, обитающих на литорали Кандалакшского и Онежского заливов Белого моря
| Таксономический статус беспозвоночных | Экологические группы | Широтные категории | Долготные категории | Кандалакшский залив | Онежский залив | |||||
| Тип | Класс | Отряд | Семейство | Род | Вид | |||||
| Porifera Губки | Desmospongiae | Halichondrida | Halichhondriidae | Halichondria | Halichondria panicea (Pallas,1766) | Эп; 2 | А-b | Kos в северном полушарии | + | + |
| Haliclonidae | Halichlona | Halichlona gracilis (Miclucho-Maclay, 1870) | Эп; 2 | А-b | Cp, pac | + | + |
| Cnidaria (Coelenterata) | Anthozoa | Actiniaria | Actiniidae | Aulactinia | Aulactinia stella (Verrill, 1864) | Эп, он; 2 | A | Cp | + | + |
| Scyphozoa | Stauromedusae | Haliclystidae | Haliclystus | Haliclystus auricula Rathke, 1806) | Эп; 1 | B | Atl, pac | + | ||
| Hydrozoa | Anthoatecatae | Clavidae | Clava | Clava multicornis (Forskai, 1775) | Эп; 2 | B | Atl | + | + | |
| Thecatae | Campanulariidae | Obelia | Obelia geniculata (L., 1758) | Эп; 2 | B | Atl-pac | + | + | ||
| Obelia loveni (Allman, 1859) | Эп; 2 | B | Atl | + | + | |||||
| Obelia longissima (Pallas, 1766) | Эп; 2 | A-b, bp | Atl-pac, | + | + | |||||
| Sertullariidae | Dynamena | Dynamena pumila (L., 1758) | Эп; 2 | B | Atl | + | + |
| Nemertea | Anopla | Heteronemertea | Lineidae | Poseidon | Poseidon ruber (Müller, 1774) | Ин; 2 | ? | Неизв? | + | + |
| Enopla | Hoplonemertea | Amphiporidae | Amphiporus | Amphiporus lactifloreus (Johnston, 1828) | Ин; 2 | ? | Неизв? | + | + |
| Polychaeta | Phyllodocida | Phyllodocidae | Phyllodoce | Phyllodoce maculata (Linnaeus, 1767) | Он; 2 | A-b | Atl, pac | + | ||
| Phyllodoce groenlandica (Oersted, 1842) | A-b | Cp, ws | + | + | ||||||
| Phyllodocida | Phyllodocidae | Eulalia | Eulalia bilineata Johnston,1840 | Он; 2 | B | Cp | + | |||
| Eteone | Eteone longa (Fabricius, 1780) | A-b | Cp | + | + | |||||
| Polynoidae | Harmothoe | Harmothoe imbricata (Linnaeus,1767) | Он; 2 | A-b | Pac | + | + | |||
| Nereidae | Nereis | Nereis pelagica Linnaeus, 1767 | Ин; 2 | B | Kos | + | + | |||
| Alitta | Alitta virens M. Sars, 1835 | Ин; 2 | B | Atl, pac | + | + | ||||
| Nephthyidae | Micronephthys | Micronephthys minuta Theel 1879 | Ин; 2 | A | A | + | ||||
| Orbiniida | Orbiniidae | Scoloplos | Scoloplos armiger (Muller,1776) | Ин; 2 | Cp | Wos | + | + | ||
| Spionidae | Polydora | Polydora quadrilobata Jacobi, 1883. | Ин; 2 | B-a | Ws cp | + | + | |||
| Pygospio | Pygospio elegans Claparede, 1869. | Ин; 3 | Amb | Аmb | + | + | ||||
| Capitellida | Arenicolidae | Arenicola | Arenicola marina (Linnaeus, 1758). | Ин; 2 | B | Atl | + | + | ||
| Terebellida | Pectinariidae | Pectinaria | Pectinaria hyperborea (Malmgren, 1865) | Ин; 2 | B-a | Ws cp | + | + | ||
| Pectinaria koreni (Malmgren, 1865). | Ин; 2 | St-b | Atl eur | + | ||||||
| Sabellida | Sabellidae | Fabricia | Fabricia sabella (Ehrenberg,1837) | Ин; 2 | A-b | Atl | + | + | ||
| Spirorbidae | Spirorbis | Spirorbis spirorbis (Linnaeus,1758). | Эп; 2 | A-b | Pac | + | ||||
| Oligochaeta | Naidomorpha | Tubificidae | Clitellio | Clitellio arenarius (O.F. Muller, 1776) | Ин; 2 | ? | ? | + | ||
| Tubifex | Tubifex costatus (Claparede, 1863). | Ин; 2 | ? | ? | + | |||||
| Oligochaeta | Naidomorpha | Tubificidae | Tubificoides | Tubificoides benedeni (Udekem,1855). | Ин; 2 | ? | ? | + | ||
| Naididae | Paranais | Paranais Litoralis (O.F. Muller, 1784). | Ин; 2 | ? | ? | + | ||||
| Enchytreidae | Enchytraeus | Enchytraeus albidus Henle.1837. | Ин; 2 | ? | ? | + | ||||
| Lumbricillus | Lumbricillus rivalis Levinsen, 1883. | Ин; 2 | ? | ? | + | |||||
| Enchytreidae | Lumbricillus | Lumbricillus viridis Stephenson, 1911. | Ин; 2 | ? | ? | + |
| Mollusca | Gastropoda
|
Patelliformes | Lottiidae | Testudinalia | Testudinalia tesselata (Muller, 1776). | Он; 2 | B | Ws atl eur | + | + |
| Trochiformes | Trchidae | Margarites | Margarites helicinus Phipps, 1774). | Он; 3 | B | Amb atl pac | + | + | ||
| Littoriniformes | Littorinidae | Epheria | Epheria vincta (Montagu, 1803). | Он; 3 | B | Amb ws atl pac | + | + | ||
| Lacuna | Lacuna neritoidea Gould, 1840. | Он; 3 | H-b | Atl | + | + | ||||
| Littorina | Littorina saxatilis (Olivi,1792). | Он; 2 | B | Atl | + | + | ||||
| Littorina obtusata (Linnaeus,1758). | Он; 2 | B | Atl ws | + | + | |||||
| Littorina littorea (Linnaeus,1758). | Он; 2 | B | Atl ws | + | + | |||||
| Rissoidae | Onoba | Onoba aculeus (Gould, 1841). | Он; 2 | H-b | Atl | + | + | |||
| Hydrobiidae | Hydrobia | Hydrobia ulvae (Pennant,1777). | Он; 2 | B | Atl ws | + | + | |||
| Skeneopsidae | Skeneopsis | Skeneopsis planorbis (Fabricius, 1780). | Он; 2 | B | Atl ws | + | + | |||
| Natciformes | Naticidae | Cryptonatica | Cryptonatica clausa (Gmelin, 1791). | Он; 2 | B-a | Ws atl pac | + | + | ||
| Lunatia | Lunatia pallida (Broderip et Sowerby,1829). | Он; 2 | B-a | Cp atl pac | + | + | ||||
| Bucciniformes | Buccinidae | Buccinum | Buccinum undatum Linnaeus, 1758. | Он; 3 | B | Ws atl | + | + | ||
| Bivalvia | Mytiliformes | Mytilidae | Musculus | Musculus laevigatus (Grey, 1824). | Он; 2 | B-a | Ws atl pac | + | + | |
| Mytilus | Mytilus edulis Linnaeus, 1758. | Эп,он; 2 | B | Amb ws atl pac | + | + | ||||
| Cardiiformes
|
Tellinidae | Macoma | Macoma balthica (Linnaeus, 1758). | Ин; 2 | B | Amb atl pac | + | + | ||
| Myidae | Mya | Mya arenaria (Linnaeus, 1767). | Ин; 2 | B | Atl | + | + |
| Arthropoda
Подтип Mandibulata Инфратип Crustacea
|
Maxillipoda | Надотряд Thoracica
Отряд Cirripedia |
Balanidae | Semibalanus | Semibalanus balanoides (Linnaeus, 1758) | Эп; 2 | A-b | Ws | + | + |
| Malacostraca
Eumalacostraca |
Надотряд Peracarida
Отряд Amphipoda
|
Pontoporeiidae | Pontoporeia | Pontoporea affinis Lindstrom,1885 | Он; 2 | A-b | Ws atl pac | + | ||
| Pontoporea femorata (Kroyer,1842) | Он; 2 | A-b | Ws atl pac | + | + | |||||
| Atylidae | Atylus | Atylus carinatus (Fabricius, 1793) | Он; 2 | A | Cp ws | + | ||||
| Gammaridae | Gammarus | Gammarus oceanicus (Segerstrale, 1947) | Он; 2 | B | Ws atl | + | + | |||
| Gammarus duebeni (Lilljeborg, 1851) | Он; 2 | B | Ws atl | + | + | |||||
| Marinogammarus | Marinogammarus obtusatus (Dahl,1938) | Он; 2 | B | Ws atl | + | |||||
| Lagunogammarus | Lagunogammarus setosus Dementieva, 1931 | Он; 2 | B-a | Ws cp atl pac | + | |||||
| Lagunogammarus zaddachi Sexton, 1912 | Он; 2 | B | Ws atl | + | ||||||
| Acanthogammarus | Acanthogammarus loricatus (Sabine, 1821 et 1824) | Он; 2 | A-b | Atl | + | |||||
| Lysianassidae | Pseudolibrotus | Pseudolibrotus litoralis (Kroyer, 1845) | Ин; 1 | A-b | Ws | + | + | |||
| Amphithoidae | Amphithoe | Amphithoe rubricata (Montagu, 1808) | Он; 2 | B | Amb ws | + | + | |||
| Corophiidae | Crassicorophium | Crassicorophium bonellii (Milne Edwards, 1830) | Ин; 2 | A-b | Kos atl pac | + | + | |||
| Isopoda | Janiridae | Jaera | Jaera albifrons Leach, 1814 | Он; 2 | B | Ws atl | + | + | ||
| Decapoda | Crangonidae | Crangon | Crangon crangon (Linnaeus, 1758) | Ин; 1 | B | Atl | + | + |
| Nematoda | Enoplea | Enoplida | Enoplidae | Enoplus | Enoplus communis (Bastian, 1865). | Ин; 1 | A-b | Atl | + | |
| Oncholaimidae | Pontonema | Pontonema vulgare (Bastian, 1865). | Ин; 1 | A-b | Atl | + | ||||
| Cephalorhyncha | Priapulida | Priapulomorpha | Priapulidae | Priapulus | Priapulus caudatus Lamark,1816 | Ин; 2 | A-b | Cp | + | + |
| Halicriptomorpha | Halicriptydae | Halicryptus | Halicryptus spinulosus Siebold, 1849 | Ин; 2 | A-b | Cp | + | |||
| Bryozoa | Euristomata | Alcyonidida | Flustrellidae | Flustrellidra | Flustrellidra hispida (Fabricius, 1780). | Эп; 1 | A-b | Atl pac | + | + |
| Membraniporida | Membraniporidae | Electra | Electra pilosa (Linnaeus, 1767). | Эп; 2 | A-b | Atl | + | + | ||
| Electra crustulenta Borg, 1931. | Эп; 2 | A-b | Atl | + | + | |||||
| Echinodermata | Asteroidea | Forcipulatida | Asteriidae | Asterias | Asterias rubens (Linnaeus, 1758) | Он; 3 | B | Ws atl | + | + |
Примечание. Названия биогеографических категорий приведены под рис. 4 и 6.
Фауна беспозвоночных литорали Кандалакшского и Онежского заливов насчитывает всего 83 вида, относящихся к 71 роду, 56 семействам, 32 отрядам, 17 классам и 10 типам (табл. 2; рис. 2).
Таблица 2. Фауна беспозвоночных литорали Кандалакшского и Онежского заливов
| Исследованные регионы Белого моря | Количество таксономических рангов | |||||
| Типов | Классов | Отрядов | Семейств | Родов | Видов | |
| Кандалакшский залив | 10 | 17 | 32 | 56 | 71 | 83 |
| Онежский залив | 10 | 14 | 26 | 43 | 50 | 56 |
| Вся фауна исследованных заливов | 10 | 17 | 32 | 56 | 71 | 83 |
| Коэффициент Жаккара, % | 100 | 82 | 81 | 77 | 70 | 67 |
| Коэффициент общности Съëренсена, % | 100 | 90 | 90 | 87 | 83 | 81 |
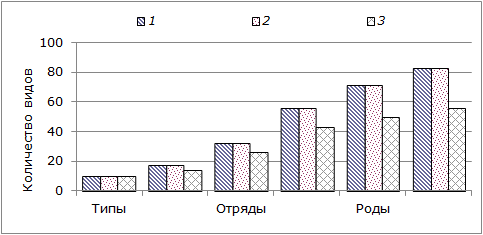
Рис. 2. Таксономическая представленность морских беспозвоночных на литорали всей исследованной акватории (1), Кандалакшского (2) и Онежского (3) заливов Белого моря
Fig. 2. Taksonomic representation of marine invertebrates in the intertidal zone within the investigated area (1), Kandalaksha (2) and Onega (3) Bay of the White Sea. The ordinate axis – the number of species
На рис. 2 хорошо видно, что наименьшим относительным разнообразием таксонов по мере снижения ранга обладают беспозвоночные литорали Онежского залива.
Обсуждение
Существует известная гипотеза о том, что в Белом море, оформившемся как самостоятельный морской водоем только в послеледниковое время и не испытавшем больших перестроек физико-химического режима, процессы дифференциации населения беспозвоночных на видовом и надвидовом уровнях не происходили (Дерюгин, 1928; Зенкевич, 1956; Бабков, Голиков, 1984). Соотношение в нем числа таксонов различного ранга в пределах каждой из систематических групп является лишь следствием дивергенции, прошедшей в прошлые геологические времена, в бассейне Северного Ледовитого и в северных частях Атлантического и Тихого океанов.
При сравнении таксономического состава бентосных беспозвоночных Белого моря с таковым в сопредельном Баренцевом необходимо отметить, что в Белое море затруднено проникновение различных видовых генотипов и морфофункциональных уровней организации в виде высших таксонов различных систематических групп. Одной из причин этого явления служит имеющийся гидрографический барьер в его Горле (проливе, соединяющем эти два водоема). Такая относительная изоляция с течением времени выражается в увеличении различий в таксономическом составе беспозвоночных между Баренцевым и Белым морями по мере снижения рангов таксонов. Другой причиной является значительно более низкая соленость беломорских вод в результате поступления большого объема пресных вод (около 200 км3 в год). В результате в целом фауна Белого моря существенно обеднена по сравнению с баренцевоморской и составляет в среднем 50–70 % (Бабков, Голиков, 1984). Обеднение коснулось в различной степени видов всех систематических и биогеографических групп. В наибольшей степени уменьшение числа видов наблюдается у стеногалинных систематических групп. В результате происходит обеднение Белого моря на родовом и особенно на видовом уровнях. В то же время подавляющее большинство семейств и отрядов в обоих морях оказываются общими.
Отношение числа видов к числу родов (среднее число видов в роде) в Белом море несколько меньше и составляет у большинства систематических групп около 2, а в более древнем и открытом Баренцевом море – 3 (Бабков, Голиков, 1984).
Если сравнивать таксономический состав литорального населения беспозвоночных Кандалакшского и Онежского заливов, то, по нашим данным, получается, что более глубоководная акватория Кандалакшского залива имеет большее видовое богатство литорального зообентоса, чем таковое в более мелководном Онежском заливе. Эта тенденция, вероятно, аналогична той, в результате которой наблюдается обеднение видового состава более мелководного Белого моря в целом по сравнению с более глубоководным Баренцевым.
Подтверждения нашим выводам содержатся в работе А. А. Голикова (1985), посвященной распределению и экологии амфипод, где на основании полученных материалов Онежский залив выделяется в отдельный биогеографический район Белого моря, а также в сводке по моллюскам Белого моря (Моллюски…, 1987), в которой отмечается, что наибольшим разнообразием малакологической таксономической группы беспозвоночных отличается Кандалакшский залив.
Распространено мнение (Белое море…, 1995), что на участках литорали, представленных твердыми грунтами (скальными и каменистыми различных модификаций), литоральное население Кандалакшского и Онежского заливов близко по составу. Нашими исследованиями в этих заливах зарегистрированы бóльшие различия для инфауны (на мягких грунтах) и сессильной онфауны (рис. 3).
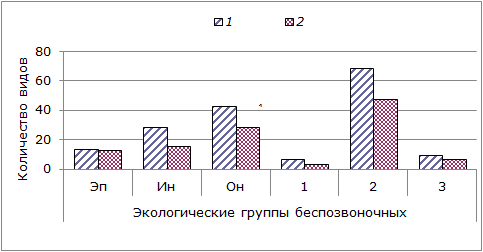
Рис. 3. Представленность различных экологических групп литорального макрозообентоса в Кандалакшском (1) и Онежском (2) заливах Белого моря. Экологические группы литоральных организмов: Эп – эпифауна – это обитатели твердых субстратов (они заселяют в первую очередь нижний и средний горизонты литорали, а на прибойных участках поднимаются и в верхний горизонт); Ин – инфауна – обитатели толщи грунта, глубина их распространения до 40 см в зависимости от характера грунта (виды инфауны заселяют почти всю литораль, избегая самого верхнего горизонта); Он – онфауна – подвижная, способная в большей или меньшей степени избегать неблагоприятных условий с использованием стратегии преферентного поведения (представители обитают по всем горизонтам литорали); 1 – виды, относящиеся к собственно литоральной фауне, весь жизненный цикл которой развертывается на периодически осушаемой литоральной полосе; 2 – фауна, общая для литорали и сублиторали или литорали и супралиторали; 3 – виды, периодически посещающие литораль, например Buccinum undatum, Asterias rubens, некоторые голожаберные моллюски и др.
Fig. 3. Representation of different ecological groups of littoral macrozoobenthos in Kandalaskha (1) and Onega (2) Bay of the White Sea. The ordinate axis – the number of species. Ecological groups of intertidal organisms: Ep – epifauna – inhabitants of solid substrates; (they inhabit primarily the lower and middle littoral horizons, and in surf line rise even to the upper horizon); In – infauna - inhabitants of soil strata , the depth of their distribution is up to 40 cm depending on the nature of the soil (infauna species inhabit almost all the littoral avoiding the uppermost horizon); On – onfauna – mobile, capable of avoiding adverse conditions to a greater or lesser extent using the strategy of preferential behavior (onfauna representatives dwells on all horizons of littoral); 1 – species pertaining to the intertidal fauna proper, its entire life cycle passes inside the intertidal zone periodically drained; 2 – fauna, common to intertidal and subtidal zones or littoral and supralittoral; 3 – species, periodically visiting the littoral, such as Buccinum undatum, Asterias rubens, some nudibranchs, etc.
Фауна беспозвоночных литорали преобладающей части прибрежной зоны морской части гидросферы, в том числе и беломорской, является «амфибиотичной». Белое море относится к бассейну Северного Ледовитого океана и с ним сообщается через открытое Баренцево море, с которым непосредственно соединено проливом, называемым Горлом, со средней глубиной 37 м. В Белом море наблюдаются правильные и мелководные полусуточные приливы. Приливная волна входит в Белое море и выходит из него через Горло, в результате уровень воды в Кандалакшском и Онежском заливах поднимается в среднем на 1.5–2 м дважды в сутки. Литораль во время отлива обсыхает, и некоторые представители инфауны выдерживают отсутствие воды максимально до 16 часов в сутки, например полихета Arenicola marina, и даже до 20 часов в сутки, например двустворчатый моллюск Macoma balthica (Свешников, 1963). Фауна литорали не так богата видами, как сублиторали (ниже уровня максимального отлива), где вода всегда присутствует, но количественно – более обильна. Обитатели литорали широко адаптированы к переживанию времени отсутствия воды и обуславливаемого им комплекса резко колеблющихся факторов окружающей среды.
На рис. 2 хорошо видно, что ин- и онфауна развиты лучше эпифауны в обоих заливах. Это закономерно, так как во время отливов физиологически в более выгодных условиях оказываются беспозвоночные, обладающие способностью глубже уйти во влажный грунт (представители инфауны) или переместиться во влажные укрытия – под камни или среди водорослей (представители подвижной онфауны).
На исследованной осушной полосе преобладает фауна, общая для литорали и сублиторали. Мало собственно литоральных форм и несколько больше видов, периодически посещающих эту прибрежную зону моря.
Интересен широтный анализ распределения различных экологических групп литорального зообентоса (рис. 4).
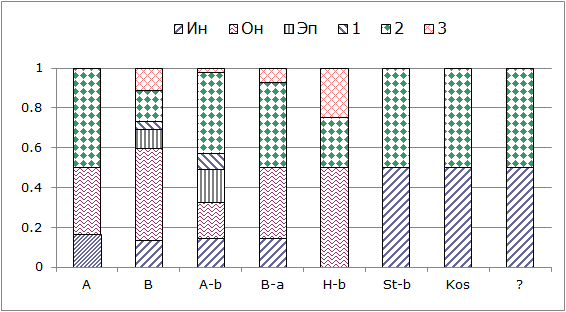
Рис. 4. Соотношение биогеографических и экологических групп литоральных беспозвоночных. Биогеографические категории: A – арктический вид, B – бореальный, A-b – арктическо-бореальный, B-a – бореально-арктический, H-b – высокобореальный, St-b – субтропическо-бореальный, Kos – космополит, ? – виды с пока неизученной биогеографической природой
Fig. 4. Ratio of biogeographical and ecological groups of intertidal invertebrates. Biogeographical categories: A – Arctic species, B – boreal species, A-b – arctic-boreal species, B-a – boreal-Arctic species, H-b – highly boreal species, St-b – subtropical-boreal species, Kos – a cosmopolitan, ? – species of unexplored biogeographical nature
Анализ показал, что арктическую фауну на литорали представляют инфаунные, онфаунные и виды, общие для литорали и сублиторали. Наиболее богато представлены все экологические группы беспозвоночных бореальной, арктическо-бореальной и бореально-арктической природы.
Наиболее распространены среди всех экологических групп литоральных беспозвоночных виды бореальной и арктическо-бореальной биогеографической природы.
Виды, входящие в состав эпифауны (эп) и относящиеся к собственно литоральной фауне (группа 1), являются преимущественно прикрепленными организмами, поэтому они имеют только бореальную и арктическо-бореальную природу. Представители ин- и онфаун, а также виды, общие для литорали и сублиторали, и виды, периодически посещающие литораль в небольшом количестве, имеют чаще всего другую биографическую природу.
Широтное распределение групп беспозвоночных представлено на рис. 5.
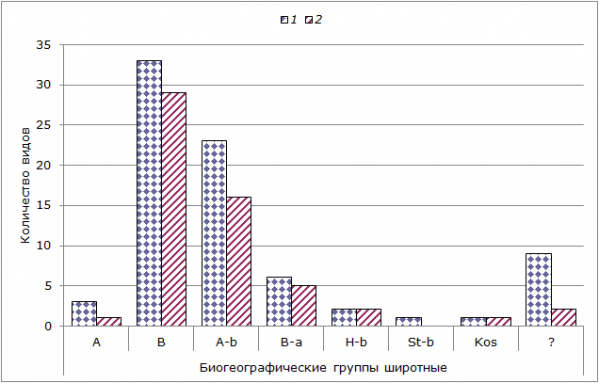
Рис. 5. Распределение биохорологических (по общности биоценологических адаптивных связей) групп беспозвоночных на литорали Кандалакшского (1) и Онежского (2) заливов. По оси ординат – количество видов
Fig. 5. Distribution of biochorologic groups (according to common biocenological adaptive relations) in the intertidal zone of Kandalaksha (1) and Onega (2) bay. The ordinate axis – the number of species.
Анализ распределения биогеографических широтных групп выявил наиболее распространенные – бореальные и арктическо-бореальные. Также довольно существенной группой являются бореально-арктические виды. Аналогичная закономерность распределения проявляется и для всего Белого моря в целом.
Арктических видов на литорали Кандалакшского залива зарегистрировано всего 3 (Cnidaria - Aulactinia stella; Annelida – Micronephthys minuta; Arthropoda – Atylus carinatus), а в Онежском заливе – 1 (Cnidaria – Aulactinia stella). Арктические условия поддерживаются в Белом море благодаря зимней конвекции и поступлению охлажденных в зимний период прибрежных баренцевоморских вод через Горло.
Большую долю составляют беспозвоночные с неопределенной пока биогеографической природой. Это недавно определенные до вида трудно идентифицируемые виды олигохет и немертин. В целом анализ биогеографического состава фаун исследованных акваторий в широтном спектре показал относительную их однородность в обоих заливах.
Распределение биогеографических долготных групп на литорали Кандалакшского и Онежского заливов приведено на рис. 6.
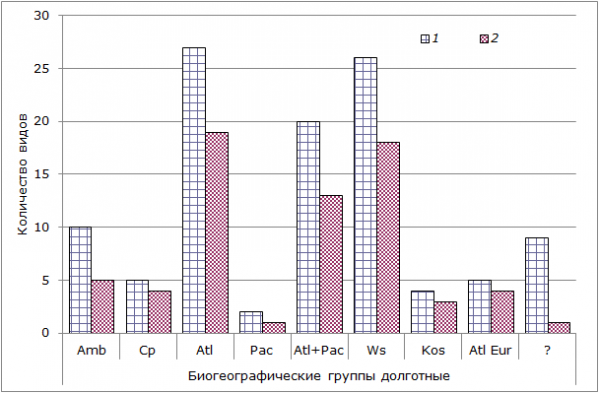
Рис. 6. Распределение биохронологических (по общности филогенетических связей) групп беспозвоночных на литорали Кандалакшского (1) и Онежского (2) заливов. Биогеографические категории: Amb – амфибореальный, Cp – циркумполярный, Atl – атлантический, Pac – тихоокеанский, Atl + Pac – атлантический и тихоокеанский, Ws – широкораспространенный, Kos – космополит, Atl Eur – атлантический европейский, ? – виды с пока неизученной биогеографической природой
Fig. 6. Distribution of biochronologic groups (according to common phylogenetic relations) in the intertidal zone of Kandalaksha (1) and Onega (2) bay. Biogeographical categories: Amb – amphiboreal, Cp – circumpolar, Atl– Atlantic, Pac – Pacific, Atl + Pac –Atlantic and Pacific, Ws – the widespread, Kos – kosmopolit, Atl Eur – Atlantic Europian, ? – Species of unexplored biogeographical nature
Анализ биогеографических групп, обитающих на литорали Кандалакшского и Онежского заливов, показал, что основу исследованной фауны составляют группы атлантической фауны, широко распространенные (широко эврибионтные) и смешанного атлантическо-тихоокеанского происхождения.
Заключение
На основании анализа литоральных беспозвоночных, собранных в двух заливах, было обнаружено сходство комплексов организмов на уровне таксонов от семейства и выше; различие – на уровне родовых и видовых.
Видовой и биогеографический состав типов Porifera, Nemertea, Bryozoa и Echinodermata в Кандалакшском и Онежском заливах аналогичны; типов Cnidaria, Nematoda и Cephalorhyncha – имеют незначительное различие. Видовой и биогеографический состав фауны литоральных червей, моллюсков и членистоногих в исследованных регионах имеет значительные различия.
Вслед за рядом исследователей (Бобринский, Зенкевич, Бирштейн, 1946; Зенкевич, 1947; Левушкин, 1974 и 1975; Тимофеев-Рессовский, 1961) мы считаем, что любой комплекс организмов, выделенный в пространстве по его положению, можно назвать биохорологическим. В результате биохорологического анализа границы комплексов литоральных организмов мы выделяли двумя взаимосвязанными между собой способами:
1 – по распространению различных систематических групп с их биохронологической иерархией (географическое распространение);
2 – по параметрам экологических условий среды в пределах распространения таксономической структуры комплексов организмов соответственно с их спектром жизненных форм (экологическое распределение).
Периодически обсыхаемое и затопляемое местообитание литоральных беспозвоночных лентовидной полосой различной ширины (от нескольких десятков до сотен метров) опоясывает изрезанное многочисленными губами и бухтами побережье Кандалакшского и Онежского заливов. На протяжении около 100 км (по прямой) оно прерывается литоральной полосой, окаймляющей Бассейн по Карельскому берегу Белого моря. Является ли этот локальный участок географической изоляцией или преградой для взаимного распространения популяций видов литоральных беспозвоночных из Кандалакшского в Онежский залив и наоборот? Находятся ли биотопы литорального населения каждого из двух исследованных нами заливов за пределами расселения, учитывая и разделение литоральной их полосы Бассейновой литоралью, и специфику водных масс, а также существенное отличие бассейнового гидрологического режима от такового в Кандалакшском и Онежском заливах?
Ответ и на первый, и на второй вопросы, по всей вероятности, в основном отрицателен. У большинства морских беспозвоночных, в том числе и литоральных беломорских, в процессе размножения образуется личиночная планктонная стадия развития, которая выполняет расселительную функцию. Радиус разноса личинок, например, отдельного массового поселения двустворчатого моллюска Mytilus edulis в Белом море составляет около 280 км (Бабков, Голиков, 1984). Кроме мидий планктонную личинку имеют Mollusca: Macoma balthica, Mya arenaria, Tonicella marmorea, Testudinalia tesselata; Arthropoda: Semibalanus balanoides,Crangon crangon; Cephalorhyncha: Priapulus caudatus; все найденные нами на литорали виды типа Bryozoa и Echinodermata, а также некоторые Annelida.
В соответствии с одной из гипотез вселения полихеты Alitta virens в Белое море из Атлантического океана, проникновение произошло постепенно путем многократного поэтапного переноса ее пелагических стадий (нерестящихся особей и их личинок) нордкапской струей теплого течения Гольфстрим (Шкляревич, 1993) от мелководья у мыса Берген, являющегося восточной границей ареала вида этой полихеты. Вместе с тем среди литоральных беспозвоночных многих типов есть и живородящие формы, например Halichondria panacea, Aulactinia stella, Littorina saxatilis, Gammarus duebeni, и откладывающие кладки, например Phyllodoce maculata, все виды отряда Nudibranchia, Littorina obtusata, Hydrobia ulvae, Buccinum undatum и др. Кроме расселения с помощью пелагических стадий развития, переносимых течениями, литоральные беспозвоночные могут перемещаться, примерзая к нижней поверхности льда, ложащегося во время отливов на субстрат литорали в осенне-зимний период образования ледового покрова. Во время разрушения ледового покрова весной отдельные фрагменты сообществ Mytilus edulis, например, уносит за пределы массовых поселений этих моллюсков (Шкляревич, 2009). Таким образом, с помощью льда могут перемещаться особи живородящих и откладывающих кладки видов с литорали Кандалакшского залива в Онежский по направлению прибрежных течений (см. рис. 1).
Выводы
В результате проведенного нами таксономического и биогеографического анализа литоральной фауны Кандалакшского и Онежского заливов можно констатировать значительное их сходство, и в то же время есть различие. Наибольшее сходство отмечается в таксономической структуре высших рангов, наименьшее – низших: видов и родов. Вместе с тем беспозвоночные литоральной фауны Кандалакшского и Онежского заливов обнаруживают определенные различия жизненных форм, которые ярко выражаются в морфологических и фенотипических отличиях особей популяций многих видов, обитающих в них. Например, двустворчатые моллюски Mya arenaria отличаются размерами, структурой поверхностного слоя раковины: кандалакшские – более крупные, внешняя поверхность раковины более ровная и гладкая, онежские – более мелкие, внешняя поверхность раковины более складчатая и морщинистая (Смолькова, 2012).
В результате наших исследований выявлено, что таксономическое и биохронологическое, а также биохорологическое распространение комплексов литоральных беспозвоночных Кандалакшского и Онежского заливов обуславливается экологической ситуацией в них. Эти комплексы обособлены как самостоятельные типы пространственного распределения организмов. Таким образом, мы считаем, что в исследованных нами регионах на литорали со времен конца последнего оледенения идет процесс становления двух относительно самостоятельных элементарных биохорологических фаунистических комплексов, внутри которых нельзя провести какой-либо зоогеографической границы.
Библиография
Бабков А. И., Голиков А. Н. Гидробиокомплексы Белого моря. Л.: Изд-во Зоол. ин-та АН СССР, 1984. 104 с.
Бабков А. И. Гидрологическая характеристика Онежского залива Белого моря // Экосистемы Онежского залива Белого моря. Л.: Изд-во Зоол. ин-та АН СССР, 1985. С. 3–10.
Белое море. Биологические ресурсы и проблемы их рационального использования / под ред. О. А. Скарлато; изд. Зоол. ин-та АН СССР. Ч. I. СПб., 1995. 249 с.
Бобринский Н. А., Зенкевич Л. А., Бирштейн Я. А. География животных. М.: Сов. наука, 1946. 456 с.
Голиков А. А. К распределению и экологии амфипод Онежского залива Белого моря // Экосистемы Онежского залива Белого моря. Л.: Изд-во Зоол. ин-та АН СССР, 1985. С. 150–170.
Дерюгин К. М. Фауна Белого моря и условия ее существования // Исследования морей СССР. 1928. Вып. 7–8.
Зенкевич Л. А. О задачах, объекте и методе морской биогеографии // Зоол. журн. 1947. Т. 26. Вып. 3. С. 201–220.
Зенкевич Л. А. Моря СССР, их фауна и флора. М.: Гос. учебно-педагогическое изд-во Министерства просвещения РСФСР, 1956. 424 с.
Иллюстрированный атлас беспозвоночных Белого моря / под общ. ред. Н. Н. Марфенина. М.: Товарищество научных изданий КМК, 2006. 312 с.
Клюге Г. А. Мшанки северных морей СССР. М.; Л.: Изд-во АН СССР, 1962. 584 с.
Колтун В. М. Кремнероговые губки северных и дальневосточных морей СССР. М.; Л.: Изд-во АН СССР, 1959. 236 с.
Левушкин С. И. К постановке вопроса об экологическом фаунистическом комплексе // Журн. общ. биол. 1974. Т. 35. № 5. С. 692–709.
Левушкин С. И. Об экологических фаунистических комплексах (на примере подземных фаун) // Журн. общ. биол. 1975. Т. 37. № 6. С. 814–828.
Малахов В. В., Адрианов А. В. Головохоботные черви (Cephalorhyncha) Мирового океана: (Определитель морской фауны). М.: Товарищество науч. изданий КМК, 1999. 328 с.
Моллюски Белого моря. Л.: Наука, 1987.
Наумов А. Д., Оленев А. В. Зоологические экскурсии на Белом море: пособие для летней учебной практики по зоологии беспозвоночных / под ред. А. А. Стрелкова Л.: Изд-во Ленингр. ун-та, 1981. 176 с.
Определители по фауне СССР, издаваемые Зоол. ин-том АН СССР. № 151. 328 с.
Свешников В. А. Биоценотические связи и условия существования некоторых кормовых беспозвоночных инфауны литорали Кандалакшского залива Белого моря // Тр. Кандалакшского гос. заповедника. Вып 4; Тр. ББС МГУ. Т. 2. Воронежское кн. изд-во, 1963. С. 114–134.
Смолькова О. В. Биология двустворчатого моллюска Mya arenaria в экосистемах литорали Белого моря: дисс. ... канд. биол. наук. Петрозаводск, 2012. 125 с.
Тимофеев-Рессовский Н. В. О некоторых принципах классификации биохорологических единиц // Тр. УРФАН СССР. 1961. Т. 27. С. 23–28.
Фауна и флора северных морей СССР. Определитель / ред. Н. С. Гаевская. М.: Сов. наука, 1948. 740 с.
Флора и фауна Белого моря: иллюстрированный атлас / под ред. А. Б. Цетлина, А. Э. Жадан, Н. Н. Марфенина. М.: Товаришество научных изданий КМК, 2010. 471 с.
Цетлин А. Б. Практический определитель многощетинковых червей Белого моря. М.: Изд-во МГУ, 1980. 114 с.
Цветкова Н. Л. Прибрежные гаммариды северных и дальневосточных морей СССР и сопредельных вод. Л.: Наука, 1975. 257 с.
Чесунов А. В., Калякина Н. М., Бубнова Е. Н. (ред.). Каталог биоты Беломорской биологической станции МГУ. М.: Товарищество научных изданий КМК, 2008. 384 с.
Шкляревич Г. А. Особенности биологии и жизненного цикла Nereis virens Sars в Белом море: Автореф. дисс. ... канд. биол. наук. Петрозаводск, 1993. 24 с.
Шкляревич Г. А. Сообщества мидий на литорали Кандалакшского залива Белого моря // Уч. зап. Петрозаводского гос. ун-та. Сер. Естеств. и техн. науки. 2009. № 11. С. 26–30.

 © 2011 - 2026
© 2011 - 2026