



Введение
Изучение видового состава и биотопического распределения малярийных комаров Верхневолжской низменности представляет большой теоретический и практический интерес, учитывая эпидемиологическое значение комаров рода Anopheles как переносчиков трансмиссивных заболеваний человека и животных. Исследования прошлых лет в обширном регионе Верхневолжья проводили в период борьбы с малярией в 30–50-е гг. XX в. (Егоров, 2011). Полученные в эти годы сведения по фауне и экологии малярийных комаров нуждаются в пересмотре в связи с описанием ряда новых видов-двойников комплекса Maculipennis. Современный список видов фауны комаров Верхневолжья включает только два вида Anopheles: Anopheles messeae Fal, 1926; Anopheles claviger Meig., 1804 (Смирнов и др., 2006; Егоров, 2011). Отсутствуют данные о других видах-двойниках комплекса Maculipennis, помимо An. messeae. Это обусловлено необходимостью применения современных цитогенетических или молекулярно-генетических методов диагностики видов. Наше исследование малярийных комаров Верхневолжской низменности призвано восполнить этот пробел.
Верхневолжская зандрово-аллювиальная низменность находится в подтаежной природной зоне. В этой зоне перекрываются ареалы ряда интерзональных видов малярийных комаров. Верхневолжская низменность простирается в центральной части Восточно-Европейской равнины и расположена на территории Московской, Тверской и Ярославской областей. Рельеф низменности сформирован московским оледенением, плоский, с отдельными холмами и заболоченными впадинами. Территорию Верхневолжской низменности окаймляет группа холмов и гряд высотой более 200 м: Клинско-Дмитровская гряда, Лихославльская гряда, Бежецкий верх, Тверская гряда (Вагнер, Манучарянц, 2003; Национальный атлас…, 2004). Низинно-болотный гидроландшафт обусловливает большое разнообразие мест выплода комаров. Особый интерес представляет биотопическое распределение близкородственных видов Anopheles, обитающих в условиях симпатрии и имеющих перекрывающиеся экологические ниши. В первую очередь это относится к видам-двойникам малярийных комаров комплекса Maculipennis. Виды этой группы способны развиваться в одних личиночных биотопах, а изменение относительной численности комаров в пространстве и во времени может служить показателем экологической специализации этих близкородственных видов. Целью данной работы было определение видового состава, относительной численности и биотопического распределения личинок малярийных комаров в местообитаниях Верхневолжской низменности и на сопредельных возвышенностях.
Материалы
Личинки 4-го возраста Anopheles были собраны в 2013–2021 гг. в 13 местообитаниях на территории Верхневолжской низменности и в 6 местообитаниях на сопредельных возвышенностях и грядах. На территории Московской области сбор материала проводили в Сергиево-Посадском городском округе: в п. Скоропусковский (56.371336, 38.142528); в Талдомском городском округе, в биотопах заказника «Журавлиная Родина» – д. Костенево (56.725472, 37.770389), д. Кунилово (56.730528, 37.757917), д. Айбутово (56.732556, 37.801722), д. Дмитровка (56.750167, 37.753944), д. Костолыгино (56.722417, 37.866972), а также в двух биотопах пгт. Вербилки (56.540160, 37.585839; 56.540042, 37.588119). На территории Тверской области выборки малярийных комаров взяты в Ржевском муниципальном округе, в д. Горки сельского поселения Итомля (56.455049, 33.891225); в г. Твери (56.797623, 36.043067); в Калининском муниципальном округе – в п. Чуприяновка (56.751500, 36.041028), в д. Старое Брянцево (56.898383, 35.796668); в Конаковском муниципальном округе – в пгт. Редкино (56.638194, 36.294667); в Лихославльском муниципальном округе – в д. Барановка (57.202067, 35.340563), в п. Приозерный (57.130333, 35.495694); в Спировском муниципальном округе – в д. Спирово (57.433425, 34.983388); в г. Кашине (57.358139, 37.595000); в Калязинском муниципальном округе – в д. Чигирёво (57.262528, 37.911056); в г. Бежецке (57.753194, 36.697861); в Сонковском муниципальном округе – в д. Новые Горицы (57.770806, 37.200167). Личинок собирали медицинской кюветой с поверхности воды в постоянных водоемах с обильной прибрежной водной растительностью.
Методы
Отловленных личинок фиксировали в спирт-уксусной смеси, приготовленной в соотношении 3:1. Видовую принадлежность малярийных комаров устанавливали по морфологическим признакам (Гуцевич и др., 1970; Званцов и др., 2003) и по рисунку дисков политенных хромосом. Парные слюнные железы извлекали под стереоскопическим микроскопом МБС-10. Из выделенных слюнных желез комаров получали временные препараты политенных хромосом. Хромосомы окрашивали 2%-ным лактоацеторсеином по стандартной методике (Перевозкин, 2007). Видовой состав малярийных комаров определяли с помощью сравнения политенных хромосом личинок с фотокартами кариотипов палеарктических видов Anopheles (Стегний, Кабанова, 1978; Artemov et al., 2018, 2021). Малярийные комары An. messeae s. l. включают два криптических вида: An. daciae Linton, Nicolescu & Harbach, 2004 и An. messeae Fall., 1926. В кариотипах личинок An. messeae s. l. регистрировали гомо- и гетерозиготные хромосомные перестройки, специфичные для этих видов (Naumenko et al., 2020; Brusentsov et al., 2023). Гомо-гетерозиготы с инверсией половой хромосомы XL0 встречаются исключительно у An. daciae, альтернативная инверсия XL1 имеется у обоих видов. Гомо- гетерозиготы с аутосомной инверсией 2R1 присутствуют в популяциях An. messeae s. s. Гомозиготы 2R00 представлены у обоих видов. Межвидовых гибридов An. daciae × An. messeae выявляли по одновременному наличию в кариотипах гетерозигот по обеим инверсиям XL0 и 2R1. Ранее было показано, что частоты гомо- и гетерозигот по инверсиям в популяциях An. daciae и An. messeae, за отдельными исключениями, значимо не отклоняются от ожидаемых согласно уравнению Харди – Вайнберга (Brusentsov et al., 2023). Для разделения видов число особей An. messeae с гомозиготами 2R00 оценивали на основании уравнения Харди – Вайнберга по формуле:
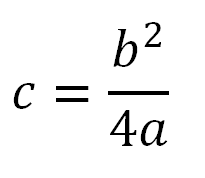
,
где c – число гомозигот 2R00 (соответствует частоте p2 · N); b – число гетерозигот 2R01 (соответствует частоте 2pq · N); a – число гомозигот 2R11 (соответствует частоте q2 · N); p и q – частоты инверсий 2R0 и 2R1; N – общее число особей в выборке.
Оценочную частоту комаров An. messeae s. s. определяли как отношение суммы особей с этими генотипами (a + b + c) к общему числу всех особей An. messeae s. l. в выборке. В отдельных выборках в случае избытка гетерозигот в локальных популяциях оценочная величина (с) может быть завышена. Учитывали, что расчетное количество всех особей An. messeae s. s. вместе с An. daciae и межвидовыми гибридами в таких популяциях не может превышать число всех особей в выборке. Кариотипы были определены у 3211 личинок.
Результаты
Результаты цитогенетического анализа выборок малярийных комаров показали, что на территории Верхневолжской низменности обитают все известные виды фауны Anopheles северной части Русской равнины: An. messeae Fall., 1926; An. maculipennis Meig., 1818; An. beklemishevi Stegn. et Kabanova, 1976; An. claviger Meig., 1804 (табл. 1). Во всех изученных местах выплода доминировали личинки An. messeae s. l., за исключением одного биотопа, где выявлен только An. claviger (выборка №22). Данный биотоп отличается от типичных мест выплода комаров комплекса Maculipennis. Личинки были отловлены в двух канавках (шириной 20–30 см, глубиной 5–12 см, длинной до 20 м) с проточной родниковой водой.
An. beklemishevi был встречен в южной части Верхневолжской низменности – в двух биотопах заказника «Журавлиная родина»; в биотопе, расположенном в пгт. Вербилки, а также в двух местообитаниях на сопредельных территориях Валдайской возвышенности и Лихославльской гряды (рис. 1). Доля An. beklemishevi среди других личинок в этих биотопах составляла 0.7–2.6 %.
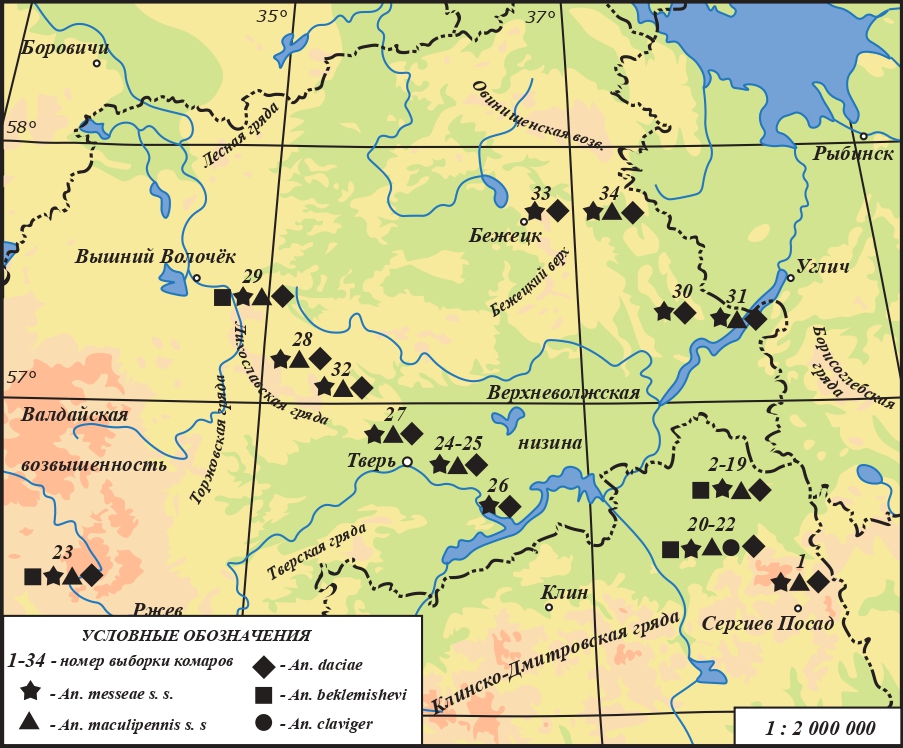
Рис. 1. Видовой состав малярийных комаров в местах сбора личинок на территории Верхневолжской низменности и на сопредельных возвышенностях
Fig. 1. Species composition of malaria mosquitoes in larval collection sites on the territory of the Upper Volga lowland and adjacent uplands
Таблица 1. Видовой состав малярийных комаров в изученных биотопах Верхневолжской низменности и сопредельных территорий
| Биотоп | Дата сбора | Видовой состав (число особей) | |||
| An. messeae s. l. | An. maculipe-nnis s. s. | An. bekle-mishevi | An.
claviger |
||
| Московская обл. | |||||
| п. Скоропусков-ский, пруд | 21.07.2016 | 51 | 30 | 0 | 0 |
| д. Костенево, пруд | 11.06.2019 -04.07.2020 | 275 | 0 | 0 | 0 |
| д. Кунилово, пруд | 12.06.2019 -30.07.2019 | 265 | 0 | 0 | 0 |
| д. Айбутово, пруд | 13.06.2019 -31.07.2020 | 452 | 0 | 0 | 0 |
| д. Дмитровка, пруд | 26.06.2019 | 128 | 0 | 1 | 0 |
| д. Костолыгино, пруд | 31.07.2019 -23.05.2021 | 487 | 2 | 5 | 0 |
| пгт. Вербилки, карьер | 05.08.2020 -03.08.2021 | 251 | 1 | 1 | 0 |
| пгт. Вербилки, родник | 03.08.2021 | 0 | 0 | 0 | 130 |
| Тверская обл. | |||||
| д. Горки, заводь р. Волги | 05.08.2013 | 119 | 2 | 1 | 0 |
| г. Тверь, пруд | 21.08.2016 | 120 | 1 | 0 | 0 |
| п. Чуприяновка, пруд | 21.06.2020 | 98 | 2 | 0 | 0 |
| пгт. Редкино, пруд | 22.06.2020 | 51 | 0 | 0 | 0 |
| д. Старое Брянцево, пруд | 22.06.2020 | 77 | 31 | 0 | 0 |
| д. Барановка, пруд | 02.07.2021 | 80 | 15 | 0 | 0 |
| д. Спирово, пруд | 03.07.2021 | 46 | 15 | 1 | 0 |
| г. Кашин, пруд | 31.07.2021 | 101 | 0 | 0 | 0 |
| д. Чигирёво, пруд | 30.07.2021 | 97 | 7 | 0 | 0 |
| п. Приозерный, озеро | 04.07.2021 | 101 | 4 | 0 | 0 |
| г. Бежецк, пруд | 13.08.2021 | 65 | 0 | 0 | 0 |
| д. Новые Горицы, пруд | 14.08.2021 | 94 | 4 | 0 | 0 |
Комары An. maculipennis были обнаружены в 12 местообитаниях как на территории Верхневолжской низменности, так и, в особенности, на сопредельных возвышенностях и грядах (рис. 1). Личинки данного вида развивались вместе с комарами An. messeae s. l. Доля особей An. maculipennis в постоянных личиночных биотопах варьировала от 1 до 37 %. Максимальные индексы доминирования этого вида отмечены в биотопах Клинско-Дмитровской гряды (37.0 ± 5.4 %; выборка № 1), а также в районе Лихославльской гряды (28.7 ± 5.3 % и 24.2 ± 5.4 %; выборки № 27 и 29 соответственно).
В изученных нами местах выплода малярийных комаров на территории Верхневолжской низменности и окружающих возвышенностей доминировали личинки An. messeae s. l. Доля An. messeae s. l. варьировала от 63 до 100 %.
Наличие преимагинальных стадий комаров An. messeae s. l. обусловлено определенными экологическими параметрами мест выплода. Для постоянных водоемов, в которых найдены комары An. messeae s. l., характерен следующий диапазон экологических характеристик. Водородный показатель колеблется в диапазоне от нейтрального до слабощелочного – 7.22–8.85. Показатели электропроводимости микросименс на сантиметр (μs) и общей доли минералов и солей в воде (ppm) варьировали в диапазоне 156–550 и 78–275 соответственно. Все водоемы, в которых были найдены личинки An. messeae s. l., характеризуются сходным составом водной растительности. В этих водоемах произрастали: роголистник погруженный (Ceratophyllum demersum L.), уруть колосистая (Myriophyllum spicatum L.), рогоз широколистный (Typha latifolia L.), рдест плавающий (Potamogeton natans L.), частуха подорожниковая (Alisma plantago-aquatica L.), элодея канадская (Elodea canadensis Michx.), хвощ топяной (Equisetum fluviatile L.), ситняг болотный (Eleocharis palustris L.), водокрас обыкновенный (Hydrocharis morsus-ranae L.), ряска (Lemna sp. L.), омежник водный (Oenanthe aquatica L.), камыш озерный (Schoenoplectus lacustris L.), подмаренник болотный (Galium palustre L.). Высокая плотность личинок выявлена в скоплениях нитчатых водорослей рода Спирогира (Spirogyra). В течение сезона размножения стабильность экологических характеристик водоемов может нарушаться под влиянием как эндогенных причин, так и антропогенных факторов. Наиболее весомую роль в изменении пригодности водоемов для проживания личинок оказывает трансформация водной и околоводной растительности. Снижение плотности личинок Anopheles неоднократно наблюдалось при активном повышении в водоемах численности свободноплавающих растений (Lemna ssp., Cladophora ssp. и др.) и резком уменьшении свободной от растений водной поверхности. В одном из контрольных водоемов заказника «Журавлиная родина» мы наблюдали исчезновение личинок An. messeae s. l. после того, как берега подверглись выкашиванию, в результате чего исчезли практически все представители свободноплавающих и укореняющихся растений.
На основании цитогенетического анализа нами было установлено, что в изученных местообитаниях комары An. messeae s. l. представлены двумя видами-двойниками: An. daciae и An. messeae s. s. Расчетная доля личинок An. messeae s. s. во всех биотопах варьирует в диапазоне 57.5–96.9 %, что свидетельствует о доминировании комаров этого вида.
Изучение хромосомного состава личинок позволило определить уровень межвидовой гибридизации между An. daciae и An. messeae s. s. Межвидовые гибриды An. daciae × An. messeae были найдены в 16 из 19 изученных личиночных биотопов. Доля межвидовых гибридов в биотопах варьировала от 0.4 до 7.8 % (табл. 2).
Таблица 2. Доля комаров An. daciae, An. messeae s. s. и их гибридов среди личинок An. messeae s. l. в местообитаниях Верхневолжской низменности и сопредельных возвышенностей
| Местообитание (номера выборок) |
Число особей | Расчетные частоты видов и их гибридов, f ± Sf (%) | ||
| An. daciae | An. messeae s. s. | Гибриды | ||
| Скоропуско-вский (№ 1) | 51 | 13.7 ± 4.8 | 82.3 ± 5.3 | 4.0 ± 2.7 |
| Костенево (№ 2–4) | 275 | 16.4 ± 2.2 | 83.6 ± 2.2 | 0 |
| Кунилово (№ 5–7) | 265 | 16.6 ± 2.3 | 82.6 ± 2.3 | 0.8 ± 0.5 |
| Айбутово (№ 8–13) | 452 | 19.2 ± 1.9 | 78.8 ± 1.9 | 2.0 ± 0.7 |
| Дмитровка (№ 14) | 128 | 18.7 ± 3.5 | 80.5 ± 3.5 | 0.8 ± 0.8 |
| Костолыгино (№ 15–19) | 487 | 15.4 ± 2.7 | 82.5 ± 1.7 | 2.1 ± 0.6 |
| Вербилки (№ 20–21) | 251 | 31.9 ± 2.9 | 66.9 ± 3.0 | 1.2 ± 0.7 |
| Горки (№ 23) | 119 | 17.6 ± 3.5 | 79.0 ± 3.7 | 3.4 ± 1.7 |
| Тверь (№ 24) | 120 | 25.0 ± 4.0 | 70.8 ± 4.2 | 4.2 ± 1.8 |
| Чуприяновка (№ 25) | 98 | 3.1 ± 1.8 | 96.9 ± 1.8 | 0 |
| Редкино (№ 26) | 51 | 21.5 ± 5.8 | 70.7 ± 6.4 | 7.8 ± 3.8 |
| Старое Брян-цево (№ 27) | 77 | 26.0 ± 5.0 | 72.7 ± 5.1 | 1.3 ± 1.3 |
| Барановка (№ 28) | 80 | 33.8 ± 5.3 | 59.9 ± 5.5 | 6.3 ± 2.7 |
| Спирово (№ 29) | 46 | 13.0 ± 5.0 | 87.0 ± 5.0 | 0 |
| Кашин (№ 30) | 101 | 14.8 ± 3.5 | 81.2 ± 3.9 | 4.0 ± 1.9 |
| Чигирево (№ 31) | 97 | 10.3 ± 3.1 | 85.6 ± 3.6 | 4.1 ± 2.0 |
| Приозерный (№ 32) | 101 | 28.7 ± 4.5 | 68.3 ± 4.6 | 3.0 ± 1.7 |
| Бежецк (№ 33) | 65 | 12.3 ± 4.1 | 86.2 ± 4.3 | 1.5 ± 1.5 |
| Новые Го-рицы (№ 34) | 94 | 18.0 ± 4.0 | 80.9 ± 4.1 | 1.1 ± 1.1 |
Обсуждение
Полученные нами данные позволяют уточнить видовой состав малярийных комаров, обитающих на территории Верхневолжской низменности. Виды An. beklemishevi, An. daciae и An. maculipennis не входили в имеющийся список видов Верхневолжья (Егоров, 2011). Впервые установлены места выплода комаров этих видов. Показано, что развитие личинок видов-двойников комплекса Maculipennis на изучаемой территории происходит в одних и тех же водоемах. Вместе с тем выявлены особенности биотопического распределения различных видов малярийных комаров.
An. claviger был найден в месте выхода грунтовых вод в окрестностях пгт. Вербилки (Талдомский городской округ Московской области). Поскольку данный вид предпочитает холодные пресные водоемы с проточной водой, совместное обитание An. claviger с видами комплекса Maculipennis наблюдается крайне редко.
An. beklemishevi является самым северным видом среди малярийных комаров. Этот вид обитает на заболоченных территориях в зоне хвойных и смешанных лесов (Novikov, 2016; Соболева и др., 2020). Самые южные местообитания данного вида были найдены на территории Мещерской низменности (Лопатин и др., 2020). An. beklemishevi приурочен к специфическим местообитаниям, его ареал не является сплошным. Биотопы, в которых был обнаружен An. beklemishevi, характеризуются повышенной затененностью и относительно низкой плотностью личинок малярийных комаров. Например, водоем в д. Дмитровка (выборка № 14) окружен сплошным кольцом из древесно-кустарниковой растительности. Берег водоема в д. Костолыгино (выборки № 17–18), в котором проводился сбор личинок, также затенен ивовыми кустарниками Salix spp. и рогозом широколистным Typha latifolia L., 1753, произрастающим в большом количестве вдоль береговой линии. Ранее экологические предпочтения совместно обитающих комаров An. beklemishevi и An. messeae s. l. исследовали в различных личиночных биотопах в пойме р. Чулым на юге Западной Сибири (Перевозкин и др., 2009). Личинки An. beklemishevi были обнаружены с повышенной частотой в мелководном затененном водоеме. В открытых прогреваемых водоемах доминировали комары An. messeae s. l. Имеющиеся данные о географическом распространении An. beklemishevi свидетельствуют о том, что южная часть ареала этого вида в подтаежной зоне Русской равнины имеет фрагментарный характер, краевые популяции малочисленны и частично изолированы друг от друга.
Малярийный комар An. maculipennis с высокой частотой встречается в местообитаниях в центре и на юге Русской равнины (Стегний, 1991). По-видимому, распространению этого вида на север таежной зоны препятствуют низкие температуры зимовки в сочетании с коротким репродуктивным сезоном. В настоящее время в условиях потепления климата происходит расширение ареала этого вида на север и на восток, на Средний и Южный Урал (Novikov, Vaulin, 2014). Согласно нашим данным, в Карелии вид продвинулся до 64 параллели. Самая северная популяция этого вида найдена в окрестностях г. Кемь (Перевозкин и др., 2012). Кроме совокупности абиотических факторов, продвижение An. maculipennis на север и восток Палеарктики может сдерживать конкуренция с комарами An. messeae s. l. Комары An. messeae s. l. относятся к полизональным видам и широко распространены в различных ландшафтно-климатических зонах Евразии. Не случайно у комаров An. messeae s. l. выявлен самый высокий уровень хромосомного полиморфизма по сравнению с другими представителями группы Maculipennis (Стегний и др., 2016).
Результаты выполненного нами цитогенетического анализа показали, что комары An. messeae s. l. представлены двумя видами-двойниками с неполной репродуктивной изоляцией – An. daciae и An. messeae s. s. Ареалы этих видов-двойников перекрываются в центре Русской равнины (Naumenko et al., 2020). Нами установлено, что оба вида встречаются во всех биотопах Верхневолжской низменности и сопредельных территорий, где происходит массовый выплод малярийных комаров (см. табл. 2). Точные данные о соотношении двух видов в совместно заселенных биотопах дает только молекулярно-генетический анализ. Главным таксономическим признаком служит нуклеотидный состав второго внутреннего транскрибируемого спейсера ITS2 кластера рибосомной ДНК: найдено 5 позиций, по которым имеются отличия у двух видов (Nicolescu et al., 2004; Naumenko et al., 2020). Молекулярно-генетическую диагностику усложняет полиморфизм по 3 из этих позиций, обнаруженный у An. daciae (Brusentsov et al., 2023). По нашему мнению, для определения географического распространения видов-двойников An. daciae и An. messeae s. s. можно использовать результаты анализа хромосомной изменчивости. Сопоставление данных молекулярно-генетического и цитогенетического анализа показало, что оба вида являются хромосомно полиморфными (Brusentsov et al., 2023). В популяциях обоих видов имеются общие и видоспецифические инверсии. Инверсия XL0 встречается с высокой частотой у An. daciae, но практически отсутствует у An. messeae s. s. С другой стороны, аутосомная инверсия 2R1 широко представлена в популяциях An. messeae s. s. на севере и в центре видового ареала, но крайне редко встречается в популяциях An. daciae (по-видимому, только в гетерозиготах у межвидовых гибридов). Наличие гомо- и гетерозигот по инверсиям XL0 и 2R1 доказывает, что оба вида присутствуют во всех водоемах, где есть личинки An. messeae s. l. Высокая частота гомо- и гетерозигот 2R11 и 2R01 (выше 50 % во всех биотопах, кроме Барановки и отдельных выборок в Вербилках и Костенево) свидетельствует о доминировании An. messeae s. s. над An. daciae. Среди гомозигот с инверсиями XL1 и 2R0 часть комаров относится к An. messeae s. s., а часть – к An. daciae. Определить видовой статус этих личинок возможно только молекулярно-генетическими методами. Но можно рассчитать долю гомозигот 2R00 An. messeae s. s., используя уравнение Харди – Вайнберга, которое является отправной точкой для анализа генетической структуры популяций (Животовский, 2021). Ранее было показано, что в большинстве популяций An. messeae s. s., обитающих в условиях симпатрии с An. daciae, отсутствуют значимые отклонения частот инверсионных генотипов от соотношения Харди – Вайнберга (Brusentsov et al., 2023). Зная число гетерозигот 2R01 и гомозигот 2R11, можно дать вероятностную оценку числа гомозигот 2R00 у An. messeae s. s. по приведенной выше формуле. Рассчитанные нами частоты гомозигот 2R00 позволили определить соотношение An. messeae s. s. и An. daciae в личиночных биотопах (см. табл. 2). Следует заметить, что в некоторых выборках оценочное число гомозигот 2R00 оказалось завышенным, поскольку сумма особей обоих видов превышает объем выборки. В этих случаях количество гомозигот 2R00 ограничили так, чтобы суммарная частота особей An. messeae s. s., An. daciae и их гибридов не превышала 100 %. Некоторое превышение оценочного числа гомозигот 2R00 у An. messeae s. s. может быть обусловлено ошибкой выборки либо эксцессом гетерозигот под воздействием факторов популяционной динамики (например, эффектом сверхдоминирования в локальных популяциях).
Полученные данные свидетельствуют о том, что в большинстве местообитаний Верхневолжской низменности происходит межвидовая гибридизация, что подтверждает вывод об их неполной репродуктивной изоляции An. daciae и An. messeae и наличии генетической интрогрессии у этих видов-двойников (Brusentsov et al., 2023). Результаты полногеномного секвенирования An. daciae и An. messeae s. s. показали, что до 20 % особей обоих видов в зонах симпатрии имеют гибридное происхождение. В то же время сравнение геномов свидетельствует о глубоких различиях нуклеотидного состава половых хромосом An. daciae и An. messeae s. s., что обеспечивает обособленность их генофондов (Naumenko et al., 2020).
Заключение
В результате проведенных исследований нами установлен видовой состав малярийных комаров в биотопах Верхневолжской низменности и окружающих ее гряд и возвышенностей. Фауна малярийных комаров представлена пятью видами: An. beklemishevi, An. claviger, An. daciae, An. maculipennis s. s., An. messeae s. s. Комар An. claviger является узкоспециализированным видом, обитает в водоемах родникового типа и экологически изолирован от других малярийных комаров. Четыре криптических вида комплекса Maculipennis развиваются совместно в личиночных биотопах. Популяции An. beklemishevi имеют очаговое пространственное распределение. Комары этого вида характеризуются низкой относительной численностью в личиночных биотопах и предпочитают затененные места выплода. An. maculipennis s. s. является субдоминантным видом и с наибольшей частотой встречается на возвышенностях. Виды-двойники An. messeae s. s. и An. daciae распространены повсеместно. Во всех совместно эксплуатируемых биотопах доминирует An. messeae s. s. Отсутствие биотопической подразделенности свидетельствует о значительном перекрывании экологических ниш у An. daciae и An. messeae s. s. Очевидно, что эти недавно разделившиеся виды находятся на раннем этапе экологической диверсификации. Об этом же свидетельствует неполная репродуктивная изоляция этих видов. Фактически вся территория Верхневолжской низменности является зоной межвидовой гибридизации между An. daciae и An. messeae s. s.
Библиография
Артемьев М. М., Баранова А. М., Ганушкина Л. А., Горностаева Р. М., Дарченкова Н. Н., Дремова В. Г., Ермишев Ю. В., Маркович Н. Я., Сергиев В. П. Малярийные комары и борьба с ними на территории Российской Федерации: Методические указания . М.: Федеральный центр госсанэпиднадзора Минздрава России, 2000. 56 с.
Беклемишев В. Н. Экология малярийного комара . М.: Медгиз, 1944. 299 с.
Гуцевич А. М., Мончадский А. С., Штакельберг А. А. Комары. Семейство Culicidae. Фауна СССР. Насекомые двукрылые . Л.: Наука, 1970. Т. 3. Вып. 4. 384 с.
Вагнер Б. Б., Манучарянц Б. О. Геология, рельеф и полезные ископаемые Московского региона . М.: МГПУ, 2003. 81 с.
Животовский Л. А. Генетика природных популяций . Йошкар-Ола: Вертикаль, 2021. 600 с.
Званцов А. Б., Ежов М. Н., Артемьев М. М. Переносчики малярии (Diptera, Culicidae, Anopheles) Содружества Независимых Государств (СНГ) . Копенгаген: ВОЗ, 2003. 312 с.
Егоров С. В. Видовой состав и структура фауны кровососущих комаров (Diptera, Culicidae) в Верхневолжье и факторы, определяющие ее динамику // Российский паразитологический журнал. 2011. № 1. С. 15–17.
Лопатин А. А., Панов В. И., Москаев А. В., Гордеев М. И. Изучение видового и хромосомного состава популяций малярийных комаров Рязанской Мещеры // Материалы Международной научной онлайн-конференции молодых ученых «Наука на благо человечества – 2020» (Москва, 20–24 апреля 2020 г.). М.: МГОУ, 2020. С. 251–256.
Национальный атлас России: В 4 т. Т. 1. Общая характеристика территории / Отв. ред.: Г. В. Поздняк, Н. Н. Полункина, Н. В. Смурова; Редкол. В. Ф. Хабаров и др. М.: Роскартография, 2004. 496 с.
Обзор погодных условий в Европейском регионе за 8–11 мая 2021 г. // Росгидромет. URL: https://www.meteorf.gov.ru/press/news/24534/?sphrase_id=791819 (дата обращения: 14.11.2023).
Перевозкин В. П. Адаптивный полиморфизм малярийных комаров комплекса Anopheles maculipennis // Научно-практическое руководство по малярии (эпидемиология, систематика, генетика). Томск: Изд-во Томского ун-та, 2007. С. 105–145.
Перевозкин В. П., Гордеев М. И., Бондарчук С. С. Хромосомный полиморфизм и закономерности формирования субпопуляционной организации малярийных комаров Anopheles (Diptera, Culicidae) в местообитаниях Томской области // Генетика. 2009. Т. 45, № 4. С. 478–487. DOI: 10.1134/S102279540904005X
Перевозкин В. П., Гордеев М. И., Москаев А. В., Ахметова Н. М., Бондарчук С. С. Распространение и инверсионный полиморфизм малярийных комаров Карелии // Генетика. 2012. Т. 48, № 7. С. 806–806.
Смирнов А. Л., Егоров C. В., Абарыкова О. Л., Петров Ю. Ф. Фауна комаров (Diptera, Culiddae) Восточного Верхневолжья Российской Федерации // Аграрный вестник Урала. Серия «Биология». 2006. № 2 (32). C. 54–56.
Соболева Е. С., Фёдорова В. С., Бурлак В. А., Шарахова М. В., Артемов Г. Н. Инверсионный полиморфизм природных популяций Anopheles beklemishevi Stegnii et Kabanova в Западной Сибири // Концептуальные и прикладные аспекты научных исследований и образования в области зоологии беспозвоночных: Сборник статей V Международной конференции, 26–28 октября 2020 г., г. Томск, Россия. Томск, 2020. С. 140–144.
Стегний В. Н. Популяционная генетика и эволюция малярийных комаров . Томск: Изд-во Томского ун-та, 1991. 136 с.
Стегний В. Н., Кабанова В. М. Хромосомный анализ малярийных комаров Anopheles atroparvus и A. maculipennis (Diptera, Culicidae) // Зоологический журнал. 1978. Т. 57, № 4. С. 613–619.
Стегний В. Н., Пищелко А. О., Сибатаев А. К., Абылкасымова Г. М. Пространственно-временные изменения частот хромосомных инверсий по ареалу малярийного комара Anopheles messeae Fall. (Culicidae) за 40-летний период мониторинга // Генетика. 2016. Т. 52, № 6. С. 664–664. DOI: 10.7868/S0016675816060138
Artemov G. N., Fedorova V. S., Karagodin D. A., Brusentsov I. I., Baricheva E. M., Sharakhov I. V., Gordeev M. I., Sharakhova M. V. New Cytogenetic Photomap and Molecular Diagnostics for the Cryptic Species of the Malaria Mosquitoes Anopheles messeae and Anopheles daciae from Eurasia // Insects. 2021. № 12(9):835. P. 1–16. DOI: 10.3390/insects12090835
Artemov G. N., Gordeev M. I., Kokhanenko A. A., Moskaev A. V., Velichevskaya A. I., Stegniy V. N., Sharakhov I. V., Sharakhova M. V. A standard photomap of ovarian nurse cell chromosomes and inversion polymorphism in Anopheles beklemishevi // Parasites and Vectors. 2018. № 11(1):211. P. 1–9. DOI: 10.1186/s13071-018-2657-3
Brusentsov I. I., Gordeev M. I., Yurchenko A. A., Karagodin D. A., Moskaev A. V., Hodge J. M., Burlak V. A., Artemov G. N., Sibataev A. K., Becker N., Sharakhov I. V., Baricheva E. M., Sharakhova M. V. Patterns of genetic differentiation imply distinct phylogeographic history of the mosquito species Anopheles messeae and Anopheles daciae in Eurasia // Mol Ecology. 2023. Sep 13. DOI: 10.1111/mec.17127
Naumenko A. N., Karagodin D. A., Yurchenko A. A., Moskaev A. V., Martin O. I., Baricheva E. M., Sharakhov I. V., Gordeev M. I., Sharakhova M. V. Chromosome and Genome Divergence between the Cryptic Eurasian Malaria Vector-Species Anopheles messeae and Anopheles daciae // Genes. 2020. № 11(2):165. P. 1–22. DOI: 10.3390/genes11020165
Nicolescu G., Linton Y. M., Vladimirescu A., Howard T. M., Harbach R. E. Mosquitoes of the Anopheles maculipennis group (Diptera: Culicidae) in Romania, with the discovery and formal recognition of a new species based on molecular and morphological evidence // Bull. Entomol. Res. 2004. № 94. P. 525–535. DOI: 10.1079/ber2004330
Novikov Y. M. On the ecology and range of Anopheles beklemishevi (Diptera: Culicidae) with reference to the taxonomy of An. lewisi // Journal of Vector Ecology. 2016. Т. 41, № 2. P. 204–214. DOI: 10.1111/jvec.12215
Novikov Y., Vaulin O. Expansion of Anopheles maculipennis s. s. (Diptera: Culicidae) to northeastern Europe and northwestern Asia: Causes and Consequences // Parasites & vectors. 2014. Vol. 7 (389). P. 1–10. DOI: 10.1186/1756-3305-7-389
Благодарности
Исследование выполнено за счет гранта РНФ № 22-24-00183 «Хромосомный полиморфизм в популяциях видов-двойников малярийных комаров таежной зоны Евразии», https://rscf.ru/project/22-24-00183/.

 © 2011 - 2026
© 2011 - 2026