



Введение
Под экологическим мониторингом понимается система регулярных повторяющихся наблюдений и оценок биотических компонентов природной среды. Как и всякое научное исследование, мониторинг имеет две основные цели: 1) возможность предсказания развития экологических процессов, 2) что еще более важно, возможность управления экологическими процессами. В наши дни значение регулярного мониторинга для науки и народного хозяйства возросло в связи с двумя обстоятельствами: 1) глобальный рост антропогенной нагрузки, 2) рост нестабильности климата и увеличения числа природных катастроф в силу не до конца изученных причин. Мониторинг водных объектов имеет среди всех форм мониторинга особое значение для нашей страны как одной из богатейших в мире гидроресурсами. Мониторинг позволяет создать базу данных, на основе которой можно строить прогностические модели. Последние можно разделить на динамические и статические. Динамические основаны на анализе динамики, временного ряда наблюдений с выявлением направления дальнейшего развития процесса. Статические основаны на одноразовой оценке состояния объекта и прогноза дальнейшего пути развития системы. Статические менее точны и требуют более наукоемких методов обработки. Но имеющийся в ряде случаев недостаток данных по динамике систем и достигнутый серьезный задел в области фундаментальной экологии приводят к необходимости и возможности активного использования статических моделей. Особо актуально это становится в наши дни в связи с недостаточным финансированием работ по мониторингу. В настоящей работе дается анализ контактных методов феногенетического мониторинга.
Методы. Феногенетическая индикация
Метод феногенетической индикации развивается с прошлого века и находит применение в изучении водных объектов (Sapunov, 1998; Sapunov, Dikinis, 2011; Сапунов, Углов, 2013). Практическое применение большинства математических моделей природных популяций требует знания многих переменных, которые можно измерить только путем долгих и дорогостоящих исследований в природе. Подобные наблюдения не всегда возможны и для большинства видов и экосистем не производились. Компенсация недостатка исходных данных может быть осуществлена на основе наукоемкого подхода с учетом достижений экологической науки. В табл. 1 представлена гипотетическая схема зависимости направления изменения популяции от экологических процессов. Можно ли на основе краткосрочного или даже единовременного анализа состояния популяций определить, к какой из возможных категорий она относится, какова ее дальнейшая судьба и какая минимальная информация требуется для предсказания судьбы популяции и экологической системы? Экологическая система включает в себя взаимодействующие популяции и окружающую их среду. Популяция характеризуется следующими признаками. Численность. На основании ее абсолютного значения выводы делать трудно. Более существенная характеристика – закон изменения численности (Nt) по времени (t). Неограниченный рост популяции описывается уравнением:
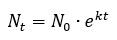
где N0 – исходная численность, k – коэффициент, отражающий репродуктивные потенции популяции.
Таблица 1. Возможные изменения популяции в зависимости от экологических условий
| Численность | Генетический состав | |
| стабильный | нестабильный | |
| Стабильная или циклически меняющаяся | стабильные, благоприятные условия |
медленные изменения среды, влекущие изменения генетического состава |
| Экспоненциально убывающая |
резко неблагоприятные условия, к которым нельзя адаптироваться |
относительно неблагоприятные условия, приводящие к сокращению численности |
| Экспоненциально возрастающая |
улучшение условий обитания и расширение возможного ареала популяции |
качественная перестройка экологической среды в лучшую сторону |
Возможны периодические колебания численности, описываемые зависимостью:
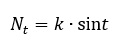
В этом случае популяцию можно рассматривать как относительно устойчивую. Изменчивость по количественным признакам обычно оценивается с помощью среднего квадратичного отклонения, коэффициента вариации. Вопрос о комплексной оценке изменчивости по ряду количественных признаков уже рассматривался в литературе (Черепанов, 1986). Предлагаем следующий несложным алгоритм. По каждому признаку все организмы разбиваются на три группы. В первой количественная мера признака (обозначим х) меньше 0.9 µ. Вторая группа включает организмы, у которых признак находится в пределах µ (1 ± 0.1), в третьей х больше 1.1 µ. Тогда количественная изменчивость по комплексу признаков описывается формулой:
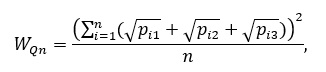 (1)
(1)
где
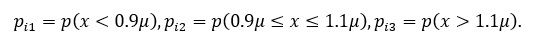
Ошибку этой величины формулы (1) следует оценивать:
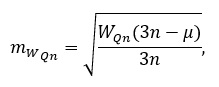
где n – число признаков.
Изменчивость по качественным признакам можно оценить двумя способами. Если преобладает один дикий тип и как исключение встречаются аномальные формы, можно оценить долю таких форм в популяции:
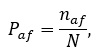 (2)
(2)
где 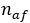 – число атипичных форм. Ошибка определяется по традиционной формуле для ошибки доли:
– число атипичных форм. Ошибка определяется по традиционной формуле для ошибки доли:
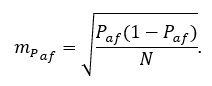
Если в популяции не столь четко выделяется дикий тип и имеется полиморфизм, то внутрипопуляционное разнообразие можно оценивать по формуле:
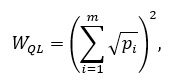 (3)
(3)
где pi – частота i-й морфы, m – число морф. Ошибка определяется:
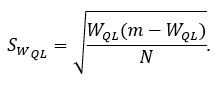
Следующая характеристика – асимметрия по билатеральным признакам. На ее основании традиционно оценивают состояние популяции (Sapunov, Dikinis, 2011). Асимметрию характеризуют показателем:
 (4)
(4)
где аr и al – значения признака на правой и левой стороне соответственно. Ошибка определяется:
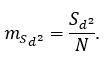
Вопрос об эволюционной роли полового диморфизма детально рассмотрен В. А. Геодакяном (Геодакян, 1983). Им установлено, что линия самцы – самки соответствует направлению эволюционного процесса. Степень полового диморфизма можно оценивать с помощью коэффициента:
 (5)
(5)
Ошибка:

Положительное значение коэффициента говорит о стремлении признака к возрастанию, отрицательное – к убыванию. Соотношение полов – легко измеряемый и информативный показатель. Оценивается как доля особей одного, чаще мужского пола:
 (6)
(6)
Ошибка:
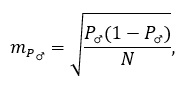
при этом 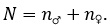 Половой диморфизм по степени количественной изменчивости, который имеет важное значение для адаптации на популяционном уровне (Геодакян, 1983), оценивается по формуле:
Половой диморфизм по степени количественной изменчивости, который имеет важное значение для адаптации на популяционном уровне (Геодакян, 1983), оценивается по формуле:
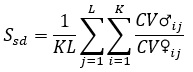 (7)
(7)
где CV – коэффициент вариации по i-му признаку у j-го самца или самки. С помощью всех этих параметров можно охарактеризовать популяцию и приближенно оценить ее судьбу в будущем. Наиболее благоприятен случай, при котором имеется хотя бы ограниченное число последовательных наблюдений. Однако возможна ситуация, когда имеется только единичная выборка животных. Как можно трактовать результаты анализа этой выборки? Данные по абсолютной численности популяции, если их можно получить, обычно малоинформативны, но и они позволяют сделать некоторые выводы. Если для раздельнополых видов численность сокращается по 150–200 особей, можно заключить, что популяция находится «при смерти». Существенная характеристика – изменчивость по количественным признакам. В норме по большинству морфологических признаков коэффициент вариации находится в пределах 0.1 (Яблоков, 1966; Sapunov, Dikinis, 2011). Достоверное превышение этой величины может свидетельствовать о генетической разбалансированности. Если речь идет о комплексе признаков (формула 1), то для порогового значения можно принять полученную на основе предыдущей цифру 2.67. Превышение этого значения позволяет предполагать, что популяция дестабилизирована. Доля аномальных форм находится в пределах 5 % в большинстве случаев. Не случайно биологическая статистика основана на эмпирически найденном уровне значимости в 0.05. Именно такая доля аномальных форм наблюдается во многих биологических популяциях (Сапунов, Углов, 2013; Федотов, 2020; Sapunov, Dikinis, 1998). На основании этого можно сказать, что появление в популяции атипичных форм с частотами больше 5 % свидетельствует, что состояние популяции неблагополучно. При анализе популяционного полиморфизма (формулы 2, 3) будем считать, что атипичные формы – редкие с регистрируемыми отклонениями от дикого типа. В качестве пороговой величины для показателя внутрипопуляционного разнообразия можно принять (формула 3) величину 1.4. Морфологическая асимметрия и степень полового диморфизма имеют значение не только по сравнению с видовой нормой. Половой диморфизм по изменчивости (формула 7) в стабильной популяции близок к 1, на начальной стадии адаптации достоверно повышается (Геодакян, 1983).
Что касается соотношения полов, то оно, несмотря на некоторые отклонения, обычно бывает близко к 1:1, т. е. доля самцов – 0.5. Сдвиг в сторону самок свидетельствует о предельной стабилизации и консервации генофонда, в сторону самцов – об адаптации популяции к неблагоприятным условиям. Чрезмерное преобладание самцов свидетельствует, что адаптивные потенции исчерпаны и популяция находится на грани вымирания. Те виды, где самки редки, можно рассматривать как эволюционно законсервированные. Таковы тли, палочники. У прогрессивных видов имеется некоторое преобладание самцов. Это большинство млекопитающих, включая человека. Преобладание самцов в несколько раз свидетельствует, что вид вымирает. Чтобы сделать более очевидными методы оценки состояния популяции, основанные на небольшом числе количественных и качественных оценок, они сведены в табл. 2. Ясно, что на основании одного параметра делать выводы о состоянии популяции трудно. Требуется комплексное изучение возможно большого числа из перечисленных параметров. Но практически иногда приходится ограничиваться лишь небольшим их числом.
Все упомянутые характеристики скоррелированы между собой. Однако степень корреляции может варьироваться в больших пределах в зависимости от того, какие конкретно признаки изучаются, от биологических особенностей объекта, экологических условий. Степень связи между всеми характеристиками – тоже признак, подверженный индивидуальной изменчивости. Поэтому алгоритм-формула должен описывать параметры суммарно, исходя из следующих соображений:
- формула должна учитывать все 6 характеристик, но работать и в том случае, когда имеется только часть характеристик;
- удельный вес функций, отражающих каждую из переменных, должен быть примерно одинаков.
Таблица 2. Возможные объяснения результатов оценки состояния популяции
| Популяционный параметр | Значение параметра | Возможное объяснение |
| 1. Изменчивость по количественным признакам | WQn < 2.67 | популяция в стабильных экологических условиях |
| WQn > 2.67 | популяция генетически разбалансирована | |
| 2. Характер распределения количественных признаков | симметричный | популяция стабильна или стрессирована |
| асимметричный | идет адаптация к новым условиям | |
| сильно асимметричный | адаптивные потенции в популяции исчерпаны | |
| бимодальный | в популяции в норме есть диморфизм или идет дизруптивный отбор | |
| 3. Изменчивость по качественным признакам | WQL < 1.4 | популяция стабильна |
| WQL < 1.4 | популяция дестабилизирована | |
| 4. Доля самцов | P = 0.05 | популяция стабильна |
| P < 0.05 | популяция высокостабильная | |
| P > 0.05 | идет адаптация к новым условиям | |
| P >> 0.05 | адаптивные возможности исчерпаны, возможно вымирание | |
| 5. Доля атипичных форм | Paf < 0.05 | популяция стабильна |
| Paf > 0.05 | популяция дестабилизирована |
Как отмечалось выше (Геодакян, 1983; Черепанов, 1986), в популяциях, находящихся в неблагоприятных условиях, повышается изменчивость, в первую очередь у особей мужского пола, растет доля самцов. Для большинства видов степень изменения этих характеристик оказывается близкой по порядку величины. Приводим пороговые значения, превышение которых говорит о неблагоприятном состоянии:

(аргументацию см. выше).
Поправочные коэффициенты позволяют выравнивать удельный вес всех характеристик и свести их в единую формулу, дающую величину С – коэффициент состояния популяции, обобщающая формулы (1–7):
 (8)
(8)
Ошибка величины определяется по формуле:
 (9)
(9)
где n – число параметров из перечисленных в формуле (8), N – объем выборки. Окончательное значение коэффициента С, отражающее то или иное состояние популяции, полученное на основе собственных и литературных данных, приведено в табл. 3.
Таблица 3. Оценка состояния популяции на основе коэффициента С
| Значение коэффициента | Состояние популяции |
| 22.02 ± mc
22.02 - mc 22.02 + mc |
неопределенное стабильное неблагоприятное |
Алгоритм работает даже при наличии данных только по нескольким из параметров. Например, если вид в норме имеет представителей только одного пола (гермафродитов или партеногенетических самок), то данные по соотношению полов и половому диморфизму не вводятся. На точность результатов это практически не влияет. Возможно, при этом потребуется увеличение размеров выборки, какое конкретно – покажет величина ошибки – формула (9).
Как пример применения феногенетической индикации можно рассмотреть ситуацию с интродукцией благородного рака Astacus astacus в озеро Еглино Бокситогорского района Ленинградской области. Этот эксперимент, начатый в 1997 г., продолжается по сей день (Федотов, 2020). За первые три года эксперимента было исследовано около 600 раков. Морфологический анализ этой выборки представлен в табл. 4. Приведены средние значения длины тела, среднее квадратичное отклонение (σ), ошибка (М) и коэффициент вариации (CV) как высокоинформативный для феногенетической индикации параметр.
Таблица 4. Морфологическое разнообразие раков Astacus astacus
| Параметры | 2000 г. | 2008 г. | 2016 г. |
| Самцы – средняя длина, мм | 112 | 102 | 105 |
| Σ | 13.7 | 22.0 | 9.9 |
| M | 1.02 | 2.9 | 1.2 |
| CV | 0.12 | 0.21 | 0.09 |
| Самки – средняя длина, мм | 103 | 101 | 98 |
| Σ | 9.46 | 9.80 | 7.0 |
| M | 1.35 | 1.2 | 1.0 |
| CV | 0.09 | 0.097 | 0.07 |
| % самцов | 48.0 | 58.5 | 59.3 |
Данные по соотношению полов и фенотипической изменчивости дополняют друг друга. Коэффициент вариации менее 0.1 для наземных животных и менее 0.15 для водных свидетельствует о популяционной нестабильности (Черепанов, 1986; Sapunov, Dikinis, 2011; Sapunov, 1998). Об этом же говорит преобладание самцов. Повышение доли самцов в отловах 2000 г. совпало с ростом фенотипической изменчивости. Самцы первыми откликаются на изменение экологической обстановки. По-видимому, в 2000 г. произошло изменение физико-химических характеристик экологической ниши раков в озере. Известно, что в 2000 г. уровень воды упал на 2–2.5 м в течение лета из-за сильной жары. После 2000 г. произошла стабилизация экологической обстановки и состояния популяции. Но превышение доли самцов по сравнению с самками говорит о том, что стабилизация не прошла до конца. Таким образом, коэффициент вариации и соотношение полов оказываются информативными показателями состояния популяции.
Постоянный мониторинг водных объектов – необходимая составляющая экологического прогноза и предотвращения экстремальных ситуаций на воде. Его проведение требует определенных расходов и регулярной работы специалистов. Вместе с тем уровень развития фундаментальных наук – экологии, математики – позволяет несколько снизить необходимые финансовые и людские ресурсы на основе развития наукоемких технологий обработки материала.
Библиография
Геодакян В. А. Эволюционная логика дифференциации полов и долголетие // Природа. 1983. № 1. С. 70–80. URL: https://textarchive.ru/c-1362724-pall.html (дата обращения: 17.08.2023).
Сапунов В. Б., Углов М. В. Из варяг в хазары по ТВС. Тихвинская водная система с древних времен до наших дней . СПб.: Химиздат, 2013. 140 с. URL: https://bigenc.ru/b/iz-variag-v-khazary-po-tvs-ti-e3c55e?ysclid=lle1dpvndf13015229 (дата обращения: 17.08.2023).
Федотов В. П. О раках – от промысла к отраслевому направлению – раководству . СПб.: Политех-Пресс, 2020. 256 с. URL: https://search.rsl.ru/ru/record/01010339654?ysclid=lle1fguhqw822100398 (дата обращения: 17.08.2023).
Черепанов В. В. Эволюционная изменчивость водных и наземных животных . Новосибирск: Наука, 1986. 380 с. URL: https://elib.natlibraryrm.ru/lib/document/SVOD/F0E01C7B-4FB2-4FC7-BA06-2C6AE5BF6F5B/ (дата обращения: 17.08.2023).
Яблоков А. В. Изменчивость млекопитающих . М.: Наука, 1966. 320 с. URL: https://www.phantastike.com/evolution/izmenchivost_mlekopitayushchikh/djvu/view/ (дата обращения: 17.08.2023).
Sapunov V. Clean ecological methods for sustainable development of urban area under pressure of urban pests // Geophysical Research Abstracts. Vol. 21, EGU2019-225, 2019 EGU General Assembly 2019. URL: https://meetingorganizer.copernicus.org/EGU2019/EGU2019-225.pdf (дата обращения: 17.08.2023).
Sapunov V. Quantitative approach to species variability of insects // VI Eur Congr Entomol: Ceske Budejovice, Ac. Sci Czech Rep. 1998. P. 309.
Sapunov V. B., Dikinis A. V. Approaches toward ecological quality essay for the Baltic region // XII Intern Environ forum “Baltic sea day”. St. Petersburg, 2011. P. 362.
Благодарности
Авторы благодарят астаколога Валерия Павловича Федотова за помощь в получении и обработке гидробиологических данных и посвящают статью его памяти.

 © 2011 - 2026
© 2011 - 2026