



Введение
Река представляет собой важнейший узел связей природных процессов, одно из самых главных звеньев в круговороте веществ на Земле, своеобразное зеркало окружающей географической среды, включая изменения, произведенные в ней человеческой деятельностью. Населенные пункты, промышленность и наиболее интенсивное сельское хозяйство размещаются вблизи рек. Это связано с тем, что речная долина обладает самым большим из природных ландшафтов разнообразием природных условий, а также с тем, что в большинстве случаев она образует более благоприятную естественную базу для производства, чем прилегающие водоразделы (Саушкин, 1980). На территории Забайкальского края речная сеть представлена более чем 40000 водотоков, около 98 % которых имеют длину менее 25 км. Большинство рек в той или иной степени используется в хозяйственной деятельности человека: добыча полезных ископаемых (Бальджа, Кыра, Могоча, Желтуга, Унда и др.), нужды гидроэнергетики (Онон и Ингода), естественный водный (Шилка) и санный (в зимнее время) путь, промышленное и бытовое водоснабжение, строительство гидротехнических сооружений (Онон, Аргунь), рыбное хозяйство (Чита) и пр.
Использование территории р. Амазар – одной из крупных водных артерии Забайкальского края – в хозяйственной деятельности человека началось в середине XIX в. с добычи россыпных месторождений золота. В последнее десятилетие интерес к данному водотоку увеличился, что обусловлено строительством гидроузла водоснабжения целлюлозного завода (ООО Целлюлозно-промышленный комбинат (ЦПК) «Полярная») в Могочинском районе Забайкальского края. В соответствии с бизнес-планом ООО ЦПК «Полярная» строительство лесопромышленного комплекса в пгт Амазар планируется на двух площадках. Первая площадка – строительство целлюлозного завода по производству небеленой сульфатной целлюлозы; вторая – строительство лесопильного завода по производству пиломатериалов, отходы от которого будут применяться для производства целлюлозы. Для бесперебойной работы в зимний период Амазарского целлюлозного завода было создано водохранилище путем перегораживания плотиной р. Амазар, в 8 км от пгт Амазар. Строительство плотины завершено в 2017 г.
Биота р. Амазар практически не изучена. Имеются лишь краткие сведения о фауне рыб (Замана, Михеев, 2005) и беспозвоночных (Матафонов, 2005; Казыкина, 2011; Зыкова, 2011; Афонина, Зыкова, 2019). Целью гидробиологических исследований, проведенных в 2018, 2019 и 2021 гг., являлось изучение разнообразия и структуры планктонных организмов – водорослей и беспозвоночных в бассейне р. Амазар (естественные участки – р. Амазар, притоки Крестовая и Большая Чичатка и искусственно созданные гидротехнические сооружения – водохранилище и рыбоход).
Материалы
Изучение биоты в бассейне р. Амазар проводилось в мае, августе, октябре 2018 г., мае, октябре 2019 г. и июле 2021 г. Обследовались р. Амазар (средний и нижний участки), притоки Крестовая и Большая Чичатка, а также искусственно созданные гидротехнические сооружения (водохранилище, рыбоход) (рис. 1). Основные характеристики абиотических параметров водных объектов бассейна р. Амазар, полученные при проведении исследований в 2018, 2019 и 2021 гг., приведены в табл. 1.

Рис. 1. Карта-схема мест отбора гидробиологических проб в бассейне р. Амазар в 2018, 2019 гг. Номера станций: 1 – р. Амазар, пгт Амазар (N 53.848630° E 120.864033°); 2 – р. Амазар, выше плотины (N 53.467120° E 120.554360°); 3 – р. Амазар, ниже плотины (N 53.774483° E 120.922900°); 4 – р. Амазар, ниже впадения р. Крестовая (N 53.772673° E 120.921453°); 5 – р. Амазар, старица; 6 – р. Амазар, устье (N 53.463382° E 122.031382°); 7 – р. Крестовая, устье (N 53.774950° E 120.921683°); 8 – р. Крестовая, стройка (N 53.785884° E 120.898976°); 9 – р. Большая Чичатка (выше плотины водохранилища) (N 53.785884° E 120.898976°); 10 – водохранилище, верхний участок (N 53.865680° E 120.885639°); 11 – водохранилище, средний участок (N 53.802519° E 120.939566°); 12 – водохранилище, нижний участок (N 53.778883° E 120.92583°); 13 – рыбоход (N 53.777622° E 120.922089°)
Fig. 1. Schematic map of hydrobiological sampling sites in the Amazar River basin in 2018, 2019. Site numbers: 1 – Amazar River, Amazar Settlement (N 53.848630° E 120.864033°); 2 – Amazar River, above the dam (N 53.467120° E 120.554360°); 3 – Amazar river, below the dam (N 53.774483° E 120.922900°); 4 – Amazar River, below the confluence of the Krestovaya River (N 53.772673° E 120.921453°); 5 – Amazar River, eldress; 6 – Amazar River, mouth (N 53.463382° E 122.031382°); 7 – Krestovaya River, mouth (N 53.774950° E 120.921683°); 8 – Krestovaya River – construction (N 53.785884° E 120.898976°); 9 – Bolshaya Chichatka River (above the reservoir dam) (N 53.785884° E 120.898976°); 10 – reservoir, upper section (N 53.865680° E 120.885639°); 11 – reservoir, middle section (N 53.802519° E 120.939566°); 12 – reservoir, lower section (N 53.778883° E 120.92583°); 13 – fish passage (N 53.777622° E 120.922089°)
Таблица 1. Некоторые физико-химические параметры воды водных объектов бассейна р. Амазар
| Водный объект | Показатель | Глубина отбора | Температура* | Прозрачность | Мутность | Минерализация | pH | Окислительно-восстановительный потенциал | Электропроводность |
| Река Амазар | min-max | 0.3–0.5 | 1.4–21.8 | 0.3–0.5 | 15–51.4 | 33–50.9 | 6.77–7.5 | 67–202 | 52–73 |
| mean ± std.d. | 0.44 ± 0.09 | 9.10 ± 8.88 | 0.44 ± 0.09 | 26.15 ± 24.57 | 44.7 ± 6.91 | 7.23 ± 0.33 | 108.88 ± 60.5 | 60.33 ± 7.89 | |
| Водохранилище ЦПК «Полярная» | min-max | 2.1–9.8 | 0.9–21.3 | 1–2.9 | 15–52.87 | 26.3–44.7 | 6.55–7.44 | 46–195.7 | 41.44–45.3 |
| mean ± std.d. | 7.14 ± 3.51 | 8.54 ± 8.33 | 1.8 ± 0.74 | 28.77 ± 25.49 | 32.06 ± 7.73 | 7.14 ± 0.39 | 102.4 ± 70.69 | 42.26 ± 1.78 | |
| Рыбопропускное сооружение | min-max | 0.2–0.5 | 1.8–21.4 | 0.2–0.5 | 35.2–50.5 | 29–44 | 6.28–7.71 | 66–221.1 | 46–55 |
| mean ± std.d. | 0.32 ± 0.11 | 8.98 ± 8.36 | 0.32 ± 0.11 | 46.33 ± 6.43 | 37 ± 5.61 | 7.02 ± 0.56 | 135.14 ± 67.84 | 49.33 ± 3.49 | |
| Река Крестовая | min-max | 0.30–3.50 | 1.3–20.8 | 0.30–3.50 | 17–69.6 | 13–50.5 | 5.75–7.65 | 47.1–182.1 | 20–39 |
| mean ± std.d. | 1 ± 1.4 | 6.96 ± 7.91 | 1 ± 1.4 | 37.23 ± 21.97 | 30.38 ± 13.63 | 7.02 ± 0.77 | 101.28 ± 67.55 | 28 ± 6.96 | |
| Река Большая Чичатка | min-max | 0.3–0.5 | 2.0–16.3 | 0.30–0.50 | 0.5–45.1 | 26–35 | 6–8.26 | 48–218.4 | 41–50 |
| mean ± std.d. | 0.46 ± 0.09 | 7.06 ± 6.52 | 0.46 ± 0.09 | 30.3 ± 17.56 | 30 ± 3.67 | 7.08 ± 0.85 | 133.28 ± 80.44 | 45.5 ± 3.18 |
Примечание. * – температура поверхностных вод; глубина и прозрачность измерялась в м; температура – в °С; мутность – в NTU; минерализация – в мг/л; окислительно-восстановительный потенциал – в мВ; электропроводность – в мкСм/см; min-max – минимальное – максимальное значение; mean ± std.d. – среднее значение ± стандартное отклонение.
Река Амазар – левый приток р. Амур – образуется слиянием Большого и Малого Амазара, берущих начало на юго-восточных отрогах Олёкминского Становика на высоте 634 м над уровнем моря. Общая протяженность составляет 290 км, площадь водосбора – 11100 км2. На верхнем и среднем участках реки находятся дражные полигоны для добычи россыпных полезных ископаемых, вследствие чего русло и пойма утеряли свой естественный вид. По характеру течения р. Амазар относится к полугорному типу, на отдельных участках с большими уклонами носит характер горного русла. Скорость течения изменяется от 0.8 до 1.2 м/с, в паводки – от 1.3 до 2.7 м/с. Мутность воды 50–150 г/м3, вблизи горных выработок и русловых карьеров она резко возрастает. Минерализация воды небольшая (40–50 мг/л). По химическому составу вода относится к гидрокарбонатному классу и кальциевой группе (Ресурсы..., 1966; Матафонов, 2005; География..., 2009).
Река Большая Чичатка (длина 46 км, площадь водосбора 2840 км2 (Ресурсы..., 1966)) обследовалась в районе нижнего бьефа ранее построенного водохранилища, предназначенного для водоснабжения ст. Амазар. В настоящее время этого водохранилища не существует, мощный паводок весной 2018 г. разрушил и снес плотину.
В р. Крестовая, общей протяженностью 20 км, исследовался нижний участок. Здесь устье заболочено, течение замедленное, преобладали протяженные мелководные плесы, глубина не превышала 0.6 м. На водосборной площади реки ведется строительство основных объектов инфраструктуры ЦПК «Полярная». Ниже устья реки подготовлена площадка сброса сточных вод комбината.
Амазарское водохранилище расположено на правом берегу одноименной реки, в 8 км от пгт Амазар. Его протяженность составляет 7.5 км. Характерный уровень при НПУ – 429.60 м, площадь зеркала – 1.32 км2, полный статический объем – 4.73 м3. Наполнение водохранилища происходит в период весеннего половодья (апрель – май). В летне-осенний период обеспечение водопотребления производится за счет транзитного стока. Прозрачность воды изменялась от 1.0 (в мае 2019 г.) до 2.8–2.9 м (в августе 2018 г.). В соответствии с классификацией (Авакян и др., 1987) созданное водохранилище по площади и объему относится к малым, по глубине – мелководным, по морфологии ложа – речным руслового типа, по форме – простым линейно-протяженным, по характеру регулирования стока – сезонного регулирования.
На правом берегу р. Амазар находится рыбопропускное сооружение в виде железобетонного лестничного рыбохода, предназначенное для пропуска на нерест ценных пород рыб из нижнего участка гидроузла в верхний. Общая протяженность тракта составляет 215 м и включает 35 камер.
Методы
В водотоках отбор фитопланктонных проб проводился путем зачерпывания воды из поверхностного слоя, в водохранилище – послойно (поверхностный слой, глубина прозрачности и придонный горизонт) с помощью батометра Паталаса. Пробы зоопланктона в водохранилище отбирались сетью Джеди (средняя модель, диаметр ячеи 0.064 мм) тотально, в реках – процеживанием 70–120 л воды через гидробиологический сачок (диаметр ячеи 0.094). Обработку фиксированных 4 % раствором формалина планктонных проб выполняли в лабораторных условиях согласно стандартным гидробиологическим методам (Киселев, 1969; Садчиков, 2003). Биомасса фитопланктона определялась по объему отдельных клеток или колоний водорослей, при этом удельный вес принимался равным единице. Объемы водорослей приравнивали к объемам соответствующих геометрических фигур (Садчиков, 2003). Биомасса зоопланктеров вычислялась по уравнениям связи длины тела и сырой массы (Ruttner-Kolisko, 1977; Балушкина, Винберг, 1979). Общий список водорослей приведен в соответствии с системой, установленной на крупнейшем мировом альгологическом сайте AlgaeBase (Guiry, Guiry, 2018), название видов и таксонов зоопланктона дано в соответствии с современной номенклатурой (WorMS, 2021). Идентификацию видов фитопланктона проводили по определителям и сводкам, указанным в работе (Ташлыкова, 2009), зоопланктона – по определителям (Кутикова, 1970; Смирнов, 1971; Определитель..., 1995). Индекс сходства рассчитывали на основе индекса Чекановского – Съеренсена (Вайнштейн, 1976; Садчиков, 2003).
Синхронно с отбором планктонных проб с помощью многопараметрического портативного анализатора качества вод GPS-AQVAMETER (Aquaread, Великобритания) проводили измерения абиотических параметров среды (минерализация (TDS), активный водородный показатель (рН), температура воды (T), мутность (Tur), окислительно-восстановительный потенциал (ORP), электропроводность (EC)). В водных объектах реки Амазар глубину (H) измеряли с помощью лота, прозрачность воды (Tr) определяли стандартным диском Секки.
Полученные данные были подвергнуты статистической и математической обработке с использованием пакета программ Microsoft Excel 2010 и надстройки для программы Microsoft Excel XLSTAT (Addinsoft, США). Для изучения взаимосвязей структурных характеристик планктона и абиотических факторов среды применяли метод главных компонент (Principal Component Analysis, PCA). В качестве переменных моделей взяты по 32 параметра: 8 абиотических (глубина, прозрачность, температура воды, pH, минерализация, мутность, окислительно-восстановительный потенциал, электропроводность) и 24 биотичеcких (число видов фито- (nph) и зоопланктона (nz), общая численность и общая биомасса фито- (Nph, Bph) и зоопланктона (Nz, Bz), численность и биомасса основных таксономических групп водорослей (Cyanobacteria (Ncya, Bcya), Chrysophyta (Nchr, Bchr), Bacillariophyta (Nbac, Bbac), Chlorophyta (Nchl, Bchl), Charophyta (Ncha, Bcha), Euglenophyta (Neug, Beug)) и беспозвоночных (Rotifera (Nrot, Brot), Cladocera (Nclad, Bclad), Copepoda (Ncop, Bcop)). Нормализацию исходных данных проводили путем приведения к стандартной шкале с нулевыми средними значениями и единичными среднеквадратичными отклонениями (Шипунов и др., 2014). Абсолютное значение нагрузки выше 0.80 принимали за существенную связь.
Результаты
Таксономический состав. В фитопланктоне обследованных водотоков и водоемов обнаружено 85 таксонов водорослей рангом ниже рода, относящихся к 6 отделам (Cyanobacteria – 6 таксонов, Bacillariophyta – 45, Chrysophyta – 8, Dinophyta – 1, Charophyta – 8, Chlorophyta – 15 и Euglenophyta – 2). Число обнаруженных таксонов водорослей изменялось от 29 (р. Большая Чичатка) и до 64 (р. Амазар) (табл. 2).
Таблица 2. Таксономический состав водорослей и беспозвоночных бассейна р. Амазар
| Таксон | Река Амазар | Водохранилище | Река Крестовая | Рыбоход | Река Большая Чичатка |
| Водоросли | |||||
| Cyanobacteria | |||||
| Oscillatoria planctonica Woloszynska 1912 | + | + | – | – | + |
| O. geminata Schwabe ex Gomont 1892 | + | + | + | – | – |
| O. ornata Kützing ex Gomont 1892 | – | + | – | – | – |
| O. sp. | + | + | + | + | + |
| Arthrospira fusiformis (Voronikhin) Komárek & J. W. G. Lund 1990 | – | + | – | – | + |
| A. verrucosa J. B. Petersen 1923 | – | – | – | + | – |
| Bacillariophyta | |||||
| Cyclotella sp. | + | + | + | + | – |
| Aulacoseira islandica (Otto Müller) Simonsen 1979 | – | – | + | – | – |
| A. sp. | + | – | – | – | – |
| Melosira varians C. Agardh 1827 | + | + | – | – | – |
| Fragilaria crotonensis Kitton 1869 | + | – | + | + | – |
| F. radians (Kützing) D. M. Williams & Round 1987 | + | + | + | + | + |
| F. virescens (Ralfs) D. M. Williams & Round 1988 | + | + | + | + | + |
| Diatoma vulgaris Bory 1824 | + | + | – | – | + |
| D. vulgaris var. producta Grunow 1862 | + | + | + | + | – |
| D. vulgaris f. capitata Cleve-Euler, 1953 | + | + | – | – | – |
| D. vulgaris var. constrictum Grunov | – | + | – | – | + |
| D. vulgaris var. ehrenbergii (Kutzing) Grunow | + | + | – | + | – |
| Synedra sp. | + | + | + | – | – |
| Ulnaria ulna (Nitzsch) Compère in Jahn et al. 2001 | + | + | + | + | + |
| Hannaea arcus (Ehrenberg) R. M. Patrick in R. M. Patrick & C. W. Reimer 1966 |
+ | + | + | + | + |
| Cocconeis placentula Ehrenberg 1838 | + | + | + | + | – |
| Achnanthes lanceolata (Brébisson ex Kützing) Grunow in Van Heurck 1880 | + | + | + | + | + |
| Nitzschia graciliformis Lange-Bertalot & Simonsen 1978 | + | + | + | + | – |
| N. sigma (Kützing) W. Smith 1853 | + | – | – | – | – |
| N. sigmoidea (Nitzsch) W. Smith 1853 | + | + | – | + | – |
| N. sp. | + | + | + | + | + |
| Tabularia fasciculata (C. Agardh) D. M. Williams & Round 1986 | + | + | + | + | + |
| Meridion circulare (Greville) C. Agardh 1831 | + | + | + | + | + |
| Tabellaria flocculosa (Roth) Kützing 1844 | + | + | + | + | – |
| T. fenestrata (Lyngbye) Kützing 1844 | + | + | + | + | + |
| Cymbella. sp. | + | + | + | + | + |
| C. sp.2 | + | + | – | + | + |
| Didymosphenia geminata (Lyngbye) Mart. Schmidt in A.Schmidt 1899 | + | + | – | – | – |
| Gomphonema olivaceum (Hornemann) Brébisson 1838 | + | + | + | + | + |
| G. coronatum Ehrenberg 1841 | + | + | + | + | + |
| Encyonema leibleinii (C. Agardh) W. J. Silva, R. Jahn, T. A. Veiga Ludwig & M. Menezes 2013 |
+ | – | – | + | – |
| Rhoicosphenia abbreviata (C. Agardh) Lange-Bertalot 1980 | + | + | – | – | – |
| Hippodonta capitata (Ehrenberg) Lange-Bertalot, Metzeltin & Witkowski 1996 | + | + | – | – | + |
| Navicula sp | + | + | + | + | + |
| N. sp.1 | – | + | + | + | – |
| Stauroneis anceps Ehrenberg 1843 | – | – | + | – | – |
| S. sp. | – | + | – | – | + |
| Gyrosigma acuminatum var. gallicum (Grunow) Cleve 1894 | – | – | + | – | + |
| Pinnularia sp. | – | – | + | – | – |
| Amphora ovalis (Kützing) Kützing 1844 | + | + | + | + | – |
| Rhopalodia gibba (Ehrenberg) Otto Müller 1895 | + | – | – | – | – |
| Epithemia sp. | – | + | – | – | – |
| Eunotia gracilis W. Smith 1853 | + | + | + | + | + |
| E. bilunaris (Ehrenberg) Schaarschmidt in Kanitz 1880 | – | – | + | – | – |
| Cymatopleura solea (Brébisson) W. Smith 1851 | – | + | – | – | – |
| Dinophyta | |||||
| Peridinium sp. | + | – | – | – | – |
| Chrysophyta | |||||
| Chrysococcus rufescens Klebs 1892 | + | + | + | – | + |
| C. biporus Skuja 1939 | + | + | – | – | – |
| Dinobryon bavaricum Imhof 1890 | + | – | – | – | – |
| D. divergens O.E. Imhof 1887 | + | – | – | + | – |
| D. sertularia Ehrenberg 1834 | + | + | – | – | – |
| Kephyrion spirale (Lackey) Conrad 1939 | + | – | – | – | – |
| Syncrypta volvox Ehrenberg, 1834 | – | + | – | – | – |
| Uroglena sp. | + | – | – | – | – |
| Charophyta | |||||
| Elakatothrix genevensis (Reverdin) Hindák 1962 | – | + | – | + | – |
| Closterium leibleinii Kützing ex Ralfs, 1848 | + | + | + | + | + |
| C. tumidulum F. Gay, 1884 | + | + | + | + | + |
| C. gracile Brébisson ex Ralfs 1848 | – | – | – | + | – |
| C. sp. | – | + | + | – | – |
| Cosmarium sp. | + | + | + | + | + |
| C. sp.1 | + | + | – | + | – |
| C. sp.2 | + | – | – | – | – |
| Staurastrum sp. | + | – | – | – | – |
| Chlorophyta | |||||
| Desmodesmus communis (E.Hegewald) E. Hegewald 2000 | + | – | – | – | – |
| Scenedesmus acuminatus (Lagerheim) Chodat 1902 | + | – | – | – | – |
| Monoraphidium arcuatum (Korshikov) Hindák 1970 | – | + | – | – | + |
| M. contortum (Thuret) Komárková-Legnerová in Fott 1969 | + | + | + | – | – |
| M. griffithii (Berkeley) Komárková-Legnerová 1969 | + | + | – | + | – |
| M. minutum (Nägeli) Komárková-Legnerová 1969 | + | – | + | – | – |
| M. komarkovae Nygaard 1979 | + | + | – | – | – |
| Ankistrodesmus fusiformis Corda 1838 | – | + | – | – | – |
| Ankyra ancora (G.M. Smith) Fott 1957 | + | + | + | – | – |
| Chlorotetraedron incus (Teiling) Komárek & Kovácik 1985 | – | + | – | – | – |
| Chlamydomonas sp.1 | + | + | + | + | – |
| C. sp.2 | + | + | – | – | – |
| Chlorogonium sp. | – | + | – | – | – |
| Actinastrum hantzschii Lagerheim 1882 | + | + | + | – | – |
| A. hantzschii var. subtile J. Woloszynska 1911 | + | + | – | – | – |
| Euglenophyta | |||||
| Euglena sp. | + | – | + | – | + |
| Phacus sp. | – | – | + | – | – |
| Всего таксонов | 64 | 61 | 42 | 37 | 29 |
| Беспозвоночные | |||||
| Rotifera | |||||
| Bdelloidea sp. n. | + | + | + | + | + |
| Cephalodella gibba (Ehrenberg, 1830) | + | – | – | – | – |
| C. sp. | + | – | – | – | – |
| Notommata collaris (Еhrenberg, 1832) | – | – | - | + | – |
| N. sp. | + | – | + | + | + |
| Trichocerca longiseta (Schrank, 1802) | + | + | - | + | – |
| T. elongata (Gosse, 1886) | + | – | – | – | – |
| Proales theodora (Gosse, 1887) | + | + | – | + | – |
| P. dolaris (Rousselet, 1895) | – | – | – | – | + |
| P. decipiens (Ehrenberg, 1832) | + | – | – | – | – |
| P. sp. | + | + | + | + | – |
| Lecane luna (Müller, 1776) | + | – | - | + | – |
| L. lunaris (Ehrenberg, 1832) | – | + | – | – | – |
| Ploesoma truncatum (Levander, 1894) | – | + | – | – | – |
| Polyarthra vulgaris Carlin, 1943 | + | – | – | – | – |
| Synchaeta pectinata Ehrenberg, 1832 | – | + | – | – | – |
| S. sp. | + | – | – | – | – |
| Lepadella patella (Müller, 1773) | – | – | + | - | – |
| Encentrum sp. | – | + | – | – | – |
| Euchlanis dilatata Ehrenberg, 1832 | + | + | – | – | + |
| E. meneta Myers, 1930 | + | – | - | + | – |
| E. deflexa (Gosse, 1851) | + | + | + | – | – |
| E. alata Voronkov, 1912 | – | – | + | - | – |
| E. incisa Carlin, 1939 | – | + | – | – | – |
| E. triquetra Ehrenberg, 1838 | + | – | – | – | – |
| E. lyra (Hudson, 1886) | + | + | – | – | – |
| Brachionus quadridentatus quadridentatus Hermann, 1783 | + | – | – | – | – |
| Keratella cochlearis (Gosse, 1851) | – | + | – | – | – |
| K. quadrata (Müller, 1786) | – | + | – | – | – |
| K. serrulata curvicornis (Rylov, 1926) | – | – | - | + | – |
| Notholca acuminata (Ehrenberg, 1832) | – | + | – | – | – |
| Platyias quadricornis (Ehrenberg, 1832) | – | + | – | – | – |
| Trichotria similis (Stenroos, 1898) | + | + | + | – | – |
| T. tetractis (Ehrenberg, 1830) | + | + | – | – | – |
| T. truncata Whitelegge, 1889 | – | + | – | – | – |
| Filinia longiseta (Ehrenberg, 1834) | + | – | – | – | – |
| Testudinella patina (Hermann, 1783) | – | + | – | – | – |
| Conochilus unicornis Rousselet, 1892 | + | + | – | – | – |
| C. hippicrepis (Schrank, 1803) | – | + | – | – | – |
| Cladocera | |||||
| Diaphanasoma sp. | + | + | – | – | – |
| Simocephalus vetulus (O. F. Müller, 1776) | – | + | – | – | – |
| Daphnia sp. | + | + | – | – | – |
| Scapholeberis mucronata (O. F. Müller, 1776) | + | + | – | – | – |
| Bosmina (Bosmina) longirostris (O. F. Müller, 1785) | + | + | – | – | – |
| Eurycercus lamellatus (O. F. Müller, 1776) | – | + | – | – | – |
| Chydorus sphaericus (O. F. Müller, 1785) | + | + | + | + | + |
| Alona quadrangularis (O. F. Müller, 1776) | – | + | – | – | – |
| A. costata Sars G. O., 1862 | – | + | + | – | – |
| A. guttata Sars G. O., 1862 | - | + | - | - | - |
| Coronatella rectangula (Sars G. O., 1862) | + | + | + | – | – |
| Alonella excisa (Fischer, 1854) | + | – | – | – | – |
| Disparalona rostrata (Koch, 1841) | – | – | - | + | – |
| Graptoleberis testudinaria (Fischer, 1848) | + | + | + | + | – |
| Biapertura intermedia (Sars, 1862) | – | – | – | + | – |
| Acroperus harpae (Baird, 1834) | + | – | - | + | + |
| Camptocercus fennicus Stenroos, 1898 | – | – | - | - | – |
| Macrothrix laticornis (Jurine, 1820) | – | + | – | + | – |
| M. sp. | + | – | – | – | – |
| Polyphemus pediculus (Linnaeus, 1761) | - | + | - | - | - |
| Copepoda | |||||
| Arctodiaptomus sp. | – | + | – | – | – |
| Nauplii, copepodita Diaptomidae | + | + | – | – | – |
| Heterocope sp. | + | – | – | – | – |
| Eucyclops serrulatus (Fischer, 1851) | – | + | + | + | – |
| E. denticulatus (Graeter, 1903) | – | + | – | – | – |
| Paracyclops fimbriatus (Fischer, 1853) | – | + | + | – | – |
| Acantocyclops vernalis (Fischer, 1853) | – | + | – | – | – |
| Diacyclops bicuspidatus (Claus, 1857) | – | + | – | – | – |
| Microcyclops rubellus (Lilljeborg, 1901) | – | – | – | + | – |
| Cryptocyclops bicolor Sars, 1863 | + | + | – | – | – |
| Mesocyclops leuckarti (Claus, 1857) | + | – | – | – | – |
| Nauplii, copepodita Cyclopoida | + | + | + | + | + |
| Nauplii, copepodita Harpacticoida | + | + | + | – | + |
| Всего таксонов | 34 | 41 | 15 | 17 | 5 |
Примечание. «+» – вид присутствует; «–» – вид отсутствует.
Таксономический состав водорослей планктона во все даты исследований носил диатомовый характер: в мае на долю диатомей приходилось около 50 % от общего числа таксонов, в августе и октябре – 65 и 63 % соответственно. Весенний планктон формировался при участии зеленых (28 %) и золотистых (11 %) водорослей, летний – при увеличении цианобактерий (6.7 %) и снижении Chrysophyta (8.9 %) и Chlorophyta (8.9 %), осенний – при активизации цианобактерий (9 %) и зеленых водорослей (16 %).
К наиболее часто встречаемым видам (интервал 50–100 %) относились: из диатомовых – Cyclotella sp., F. radians, D. vulgaris, U. ulna, H. arcus, C. placentula, A. lanceolata, N. graciliformis, T. fasciculata, M. circulare, T. fenestrata, Cymbella sp., из золотистых – C. rufescens, из зеленых – M. contortum, M. griffithii, из цианопрокариот – Oscillatoria sp., из харовых – C. leibleinii. Общими видами для обследованных водных объектов являлись: F. radians, U. ulna, H. arcus, A. lanceolata, T. fasciculata, M. circulare, T. fenestrata, G. olivaceum, G. coronatum, E. gracilis, C. leibleinii.
В составе зоопланктона идентифицировано 68 видов и подвидов, из которых 38 – Rotifera, 20 – Cladocera и 10 – Copepoda. Встречались также коловратки отряда Bdelloida, ювенильные стадии ракообразных из Harpacticoida, Cyclopoida и Calanoida, определение которых до вида не представлялось возможным. Общее количество обнаруженных таксонов варьировало от 5 (р. Большая Чичатка) до 41 (водохранилище) (см. табл. 2).
Зоопланктеры в пробах встречались редко и часто однажды, поэтому частота встречаемости большинства отмеченных видов составляла менее 10 %. Вид-убиквист Chydorus sphaericus, бделлоидные коловратки и ювенильные стадии циклопов встречались во всех обследованных водных объектах. Чаще других отмечались такие широко распространенные виды, как Proales theodora, Trichocerca longiseta, Euchlanis dilatata, E. deflexa, C. rectangula, Graptoleberis testudinaria, A. harpae, Eucyclops serrulatus. В сезонном аспекте наблюдалось увеличение качественного состава от весны к осени.
Индекс сходства продемонстрировал близость альгосообществ и разность мейофауны. Коэффициент сходства для водорослей варьировал от 38 до 77 %, для беспозвоночных – от 10 до 40 %. Наибольшее сходство водорослей отмечено для пары р. Амазар – водохранилище (55–77 %).
Количественные показатели альгофлоры и беспозвоночных в разных водных объектах были низкими (рис. 2, 3).
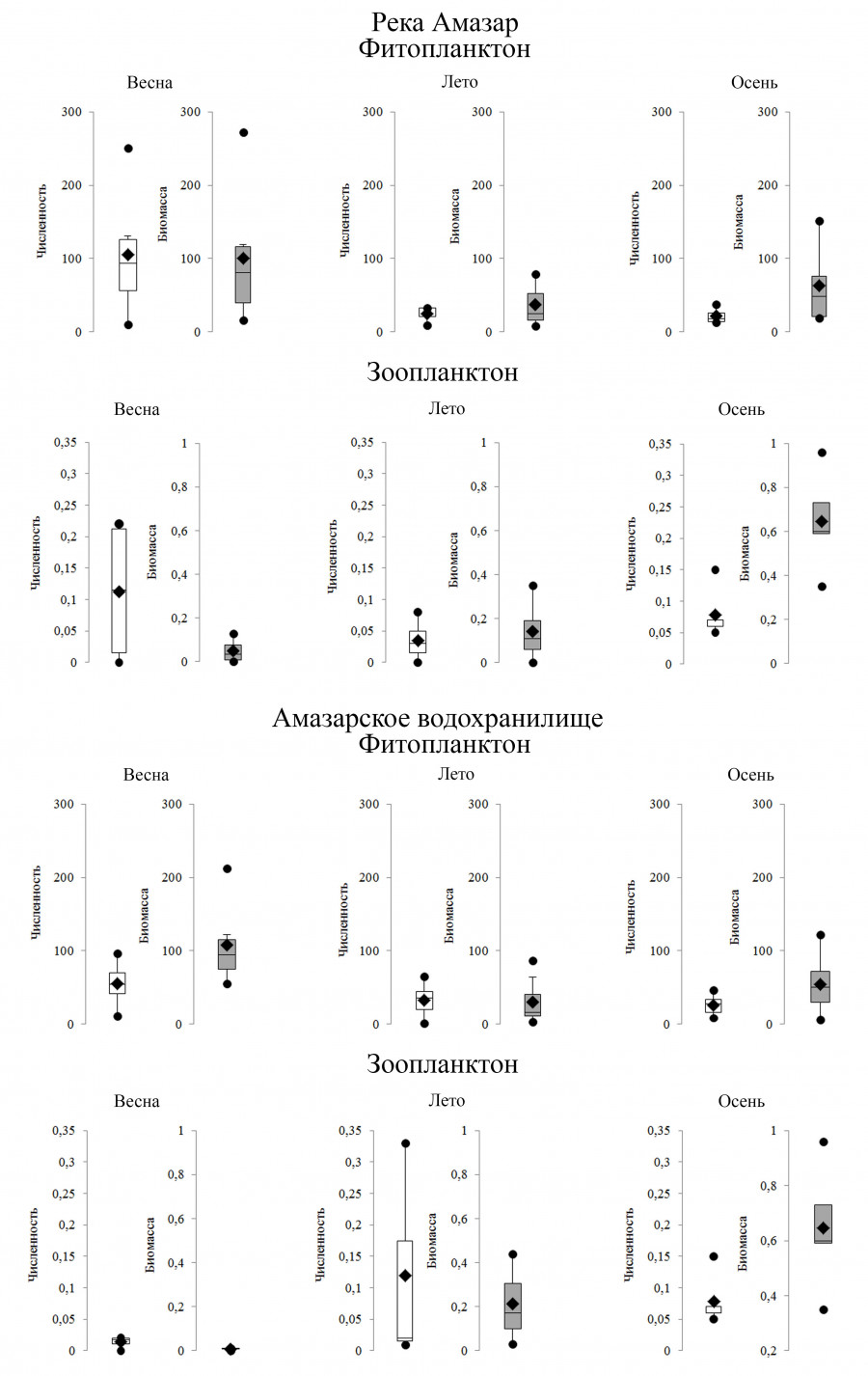
Рис. 2. Динамика численность и биомассы водорослей и беспозвоночных реки Амазар и Амазарского водохранилища в 2018, 2019 и 2021 гг. Здесь и на рисунке 3: численность фитопланктона приведена в тыс. кл./л, биомасса – в мг/м3; численность зоопланктона – в тыс. экз./м3; биомасса – в мг/м3; круг – значение min/max; ромб – среднее значение; верхняя и нижняя строны блока – 1-й и 3-й квартили
Fig. 2. Dynamics of the abundance and biomass of algae and invertebrates in the Amazar River and Amazar reservoir in 2018, 2019 and 2021. Here and in Figure 3: phytoplankton abundance is given in *103 cell/l, biomass – in mg/m3; abundance of zooplankton – in *103 ind./m3; biomass – in mg/m3; circle – min/max value; rhombus – average value; the top and bottom sides of the block – 1st and 3rd quartiles

Рис. 3. Динамика численности и биомассы водорослей и беспозвоночных, отмеченных в планктоне рек Крестовая, Большая Чичатка и рыбопропускном сооружении реки Амазар в 2018, 2019 и 2021 гг.
Fig. 3. Dynamics of the abundance and biomass of algae and invertebrates observed in the plankton of the Krestovaya and Bolshaya Chichatka rivers and the fish-passing facility of the Amazar river in 2018, 2019 and 2021
Уровень вегетации водорослей в р. Амазар изменялся от 9.6 до 250.9 тыс. кл./л. (численность) и от 8.32 до 272.44 мг/м3 (биомасса). Минимальные значения численности отмечались в августе 2018 г., максимальные – в мае 2019 г. По количественному развитию в планктоне реки доминировали диатомеи. Их доля в общей численности фитопланктона составляла 36–99 %, в биомассе – 44–99 %. В обследованной пойменной старице реки Амазар численно (86.11 % от общей численности и 78.70 % от общей биомассы) преобладали золотистые водоросли, в частности C. rufescens.
В водохранилище показатели численности и биомассы определялись в пределах 12.24–55.49 тыс. кл./л и 6.4–212.2 мг/м3. Максимальные значения отмечены в мае 2018 и 2019 гг., минимальные – в октябре 2019 г. Уровень вегетации водорослей планктона на верхнем от водохранилища участке был выше, чем на среднем и нижнем. Структурообразующий комплекс видов фитопланктона образован представителями Bacillariophyta и Chlorophyta.
В рыбоходе уровень вегетации фитопланктона также был невысокий. Численность варьировала в пределах 8.74–63.21 тыс. кл./л, биомасса – 9.42–67.96 мг/м3. Значительная роль в составе фитопланктона также принадлежала представителям диатомовых водорослей.
Количественные показатели развития водорослей планктона р. Крестовая в период исследований колебались в пределах 1.96–46.58 тыс. кл./л и 1.42–177.21 мг/м3. Максимальные значения зарегистрированы в августе 2018 г., минимальные значения – в сентябре 2018 г. Основу численности и биомассы составляли диатомовые водоросли (до 96 % общей численности и до 99 % общей биомассы фитопланктона).
В р. Большая Чичатка численность водорослей варьировала в пределах 14.11–72.52 тыс. кл./л, биомасса – 26.05–64.52 мг/м3. По развитию численности и биомассы основу фитопланктона составляли Bacillariophyta (38–98 % общей численности и до 53–99 % общей биомассы фитопланктона).
В р. Амазар общая численность зоопланктона за период исследований и на разных станциях изменялась в пределах 0.02–0.22 тыс. экз./м3. Наибольшая плотность организмов в оба года регистрировалась в мае за счет развития коловраток (Proales, Euchlanis). Общая биомасса зоопланктеров составляла 0.02–0.93 тыс. экз./м3 с максимумом в октябре за счет веслоногих ракообразных (копеподиты Cyclopoida и Harpacticoida) (см. рис. 2).
В водохранилище общая численность зоопланктона составляла 0.01–0.94 тыс. экз./м3, общая биомасса 0.02–22.38 мг/м3. Наибольшая плотность организмов отмечалась в октябре 2018 г. Численно превалировали копеподы (Arctodiaptomus sp., Acantocyclops vernalis, Diacyclops bicuspidatus, Harpacticoida).
В рыбоходе количественные показатели варьировали в пределах 0.01–0.22 тыс. экз./м3 и 0.02–1.02 мг/м3 с наибольшей плотностью организмов в октябре 2018 г. В мейофауне превалировали науплии и копеподиты Cyclopoida.
Количественные показатели мейофауны в р. Крестовая варьировали от 0.01 до 0.18 тыс. экз./м3 и от 0.02 до 0.99 мг/м3, в р. Большая Чичатка – 0.02–0.14 тыс. экз./м3 и 0.02–1.15 мг/м3. Наибольшие значения показателей отмечались в октябре 2018 г. за счет развития Copepoda (Paracyclops fimbriatus, E. serrulatus, Harpacticoida) и Rotifera (Trichocerca, Proales, Notommata, Keratella).
Факторы, определяющие развитие планктона водных объектов бассейна р. Амазар. Для выявления влияния ведущих факторов, определяющих изменение структурных показателей фито- и зоопланктона, был проведен факторный анализ методом главных компонент.
При обработке данных, полученных в результате анализа, выявлено, что первые два фактора определяют 62.95–75.12 % дисперсионной нагрузки для обследованных объектов. Эти факторы были выбраны для дальнейшего анализа, т. к. остальные вносят незначительный вклад в общую дисперсиию. Для реки Амазар первые две компоненты объясняли в сумме 73.8 % дисперсии; для водохранилища – 69.91 %; для рыбохода – 62.95 %. В реках Крестовая и Большая Чичатка первые две компоненты определяли 75.12 и 72.46 % дисперсии фактических данных соответственно (рис. 4).
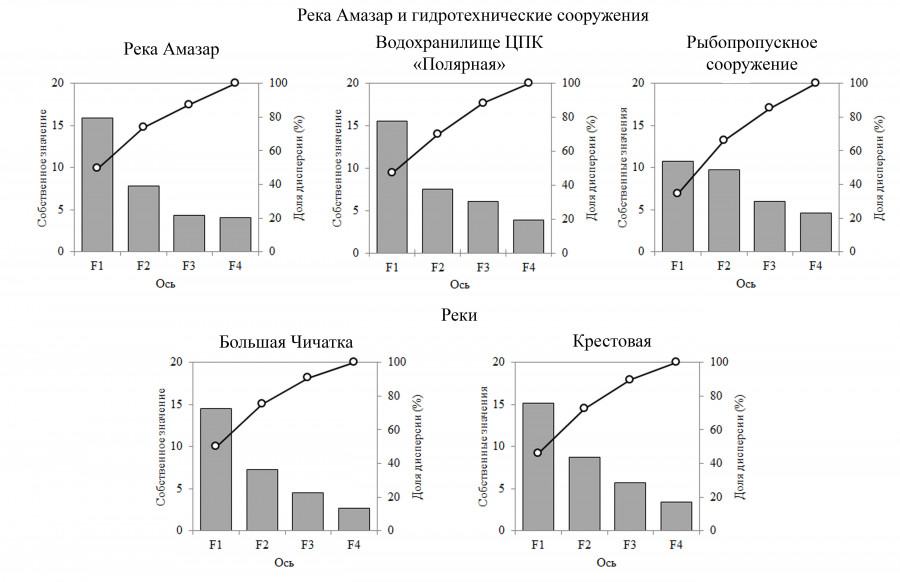
Рис. 4. Величина собственных значений (столбцы) и график накопления доли объясненной дисперсии по результатам анализа главных компонент для обследованных водных объектов
Fig. 4. The magnitude of the eigenvalues (columns) and the graph of the accumulation of the proportion of the explained variance based on the results of the analysis of the principal components for the studied water bodies
Первый фактор, отражающий общий тренд изменений (общую коррелированность) всех переменных, имеет самое высокое значение общей дисперсии в факторной матрице – 45.97–50.00 %.
Основную долю первого фактора в реке Амазар (F1: 49.51 %) и в искусственно созданных водных объектах (водохранилище (F1: 47.01 %) и рыбоход (F1: 50.03 %)) составлял гидрологический параметр, определяющий глубину исследованных водных объектов. Факторные нагрузки демонстрировали положительную реакцию структурных характеристик (общее число видов, численность и биомасса общая и таксономических групп) практически всех групп планктона на изменение данного параметра. Факторные нагрузки в первой компоненте рек Крестовая и Большая Чичатка определялись в 45.97 и 50.00 % соответственно и демонстрировали реакцию всех групп планктона по числу видов, численности и биомассе на физико-химические параметры, такие как минерализация, электропроводность, pH, окислительно-восстановительный потенциал (рис. 5).

Рис. 5. Расположение гидрохимических, гидрофизических показателей и основных характеристик планктона в пространстве двух компонент для обследованных водных объектов
Fig. 5. The location of hydrochemical, hydrophysical indicators and the main characteristics of plankton in
the space of two factors for the studied water bodies
Второй фактор в реке Амазар (F2: 24.28 %) и водных объектах (водохранилище (F2: 22.89 %) и рыбоход (F2: 25.11 %)) интегрирует информацию о физико-химических параметрах (температура, прозрачность, мутность, электропроводность, pH, минерализация) и количественных характеристиках харовых, диатомовых и динофитовых водорослей, а также ветвистоусых ракообразных. Для рек Крестовая и Большая Чичатка второй фактор, объясняя 26.49 и 25.12 % вариабельности исходных показателей соответственно, тесно связан с температурой воды (см. рис. 5), которая влияет на численность и биомассу зеленых водорослей, веслоногих рачков и коловраток.
Обсуждение
Фитопланктон Амазарского водохранилища характеризовался на момент обследования низким видовым разнообразием. Аналогичная картина отмечена для Павловского (Полева и др., 2008), Юмагузинского (Мутхудинов и др., 2012), Бурейского (Медведева и др., 2015) и других русловых водохранилищ. Для водорослей планктона Амазарского водохранилища не отмечено обильного развития цианобактерий в первые годы становления (Пирожников, 1963; Agawin et al., 2000; Bell, Kalff, 2001; Callieri, Stockner, 2002). Напротив, в обследованном водохранилище в течение всего вегетационного периода доминируют диатомовые водоросли, что характерно и для других водохранилищ подобного типа (Полева и др., 2008; Мутхудинов и др., 2012; Медведева и др., 2015 и др.).
Основу альгофлоры водохранилища создавали водоросли р. Амазар. Состав доминирующего комплекса водохранилища и реки схож, он характеризуется высокими значениями коэффициента Серенсена. Отмечены низкие значения численности и биомассы как в водотоках, так и в водохранилище.
Фауна коловраток и ракообразных Амазарского водохранилища, по данным 2018–2021 гг., характеризовалась бедным видовым разнообразием, что также отмечалось для русловых водохранилищ Енисея (Ермолаева, 2008; Попов, 2009), Иртыша (Киселева, 1967), Завхана (Крылов и др., 2019). Низкая плотность планктонной фауны Амазарского водохранилища в первые годы его существования связана с большой бедностью речного планктона, отсутствием близко расположенных пойменных водоемов, литоральной зоны, большим количеством минеральных взвесей, привносимых с амазарской водой, пагубно влияющих на жизнедеятельность зоопланктона.
Основным биофондом, за счет которого формируется зоопланктон Амазарского водохранилища, является мейофауна р. Амазар. Как для реки, так и для водохранилища отмечаются низкие количественные показатели гидробионтов. Максимальные значения численности и биомассы выявлены в лимнической зоне водохранилища, преимущественно за счет веслоногих ракообразных.
Применение метода главных компонент позволило выявить основные абиотические факторы среды, определяющие уровень развития планктона в реках и гидросооружениях бассейна реки Амазар. Установлена зависимость структурных характеристик планктона от гидрологических, физических и физико-химических параметров.
Известно, что при зарегулировании рек и образовании водохранилищ изменяется целый ряд факторов среды, водоем приобретает иной облик, происходит становление нового растительного и животного населения водоема (Ермолаева, 2008). Как отмечает ряд исследователей (Huisman et al., 1999; O'Brien et al., 2003; Huisman et al., 2002, 2006; Jäger et al., 2008 и др.), физические условия, в частности глубина водной толщи и интенсивность перемешивания, влияют на численность и динамику планктона. Эти факторы обуславливают рассеивание водорослей планктона в водной толще и противодействуют его седиментации (Visser et al., 1996; Condie and Bormans, 1997; Diehl, 2002; Huisman et al., 2002).
Такие физико-химические факторы, как температура, прозрачность, мутность, окислительно-восстановительный потенциал, рН, минерализация, определяли функционирование планктона и других речных гидробиоценозов (Descy, 1987; Potapova et al., 1999; Piirsoo et al., 2008).
Заключение
Наши исследования показали, что в р. Амазар и ее притоках Крестовая и Большая Чичатка, характеризующихся горным и полугорным характером течения, и в созданном глубоководном водохранилище, расположенном в горно-таежной зоне, сообщества альгофлоры и мейофауны мало развиты. Всего обнаружено 85 таксонов водорослей планктона рангом ниже рода и 68 видов и подвидов беспозвоночных. Наибольшее видовое разнообразие гидробионтов отмечалось в р. Амазар и водохранилище (61–64 таксона водорослей и 34–41 таксон беспозвоночных), наименьшее – в р. Большая Чичатка (29 и 5). Биофонд Амазарского водохранилища создавали водоросли и беспозвоночные р. Амазар. Показатели численности и биомассы были очень низкими и не превышали 131.24 тыс. кл./л и 272.44 мг/м3 для фитопланктона, 0.94 тыс. экз./м3 и 22.38 мг/м3 для зоопланктона. Наибольшая концентрация гидробионтов отмечалась в р. Амазар и водохранилище, наименьшая – в реках Большая Чичатка и Крестовая. Основу фитоценозов формировали преимущественно диатомовые водоросли, зооценозов – коловратки и младшевозрастные стадии копепод. Анализ влияния экологических факторов на уровень развития планктона показал, что наибольшее влияние на них оказывают гидрологические факторы, определяющие глубину водных объектов, а также физические факторы, такие как температура, прозрачность, мутность, электропроводность, pH, минерализация.
Библиография
Авакян А. Б., Салтанкин В. П., Шарапов В. А. Водохранилища . М.: Мысль, 1987. 325 с.
Афонина Е. Ю., Зыкова Е. Х. Видовой состав и количественные показатели коловраток и ракообразных среднего и нижнего течений реки Амазар (Забайкальский край) // Амурский зоологический журнал. 2019. Т. XI, № 2. С. 95–102. DOI: 10.33910/2686-9519-2019-11-2-95-102.
Балушкина Е. Б., Винберг Г. Г. Зависимость между массой и длиной тела у планктонных животных // Общие основы изучения водных экосистем / Под ред. Г. Г. Винберга. Л.: Наука, 1979. С. 169–172.
Вайнштейн Б. А. Об оценке сходства между биоценозами // Биология, морфология и систематика водных организмов. Л.: Наука, 1976. С. 156–164.
География Забайкальского края: Учебное пособие / Под ред. В. С. Кулакова. Чита: Экспресс-издательство, 2009. 308 с.
Ермолаева Н. И. Водные экосистемы. Особенности формирования зоопланктона водохранилищ: Аналитический обзор // ГПНТБ СО РАН. 2008. № 88. С. 69.
Замана Л. В., Михеев И. Е. Оценка воздействия проектируемого Амазарского водохранилища на ихтиофауну Верхнеамурского бассейна // Научные основы экологического мониторинга водохранилищ: Материалы Всерос. науч.-практ. конф. Хабаровск: ИВЭП ДВО РАН, 2005. С. 135–138.
Зыкова Е. Х. Зоопланктон реки Амазар в районе проектируемого целлюлозного завода // Кулагинские чтения: Тезисы XI Междунар. науч.-практ. конф. Чита: ЗабГУ, 2011. С. 135–137.
Казыкина С. М. Зообентосное сообщество р. Амазар в районе впадения р. Большая Чичатка // Кулагинские чтения: Тезисы XI Междунар. науч.-практ. конф. Чита: ЗабГУ, 2011. С. 137–139.
Киселев И. А. Планктон морей и континентальных водоемов . Т. 1. Л.: Наука, 1969. 658 с.
Киселева В. А. Формирование гидробиологического режима Усть-Каменогорского водохранилища : Автореф. дис. ... канд. биол. наук. Алма-Ата, 1967. 23 с.
Крылов А. В., Мэндсайхан Б., Аюушсурен Ч., Цветков А. И. Зоопланктон русловых участков разнотипных водохранилищ аридной зоны: влияние уровенного режима и метеорологических условий // Трансформация экосистем. 2019. Т. 2, № 1. С. 59–74.
Кутикова Л. А. Коловратки фауны СССР (Rotatoria) . Л.: Наука, 1970. 744 с.
Матафонов Д. В. Прогноз изменения структуры донных сообществ реки Амазар в связи со строительством Амазарского целлюлозного завода // Научные основы экологического мониторинга: Материалы Всерос. науч.-практ. конф. Хабаровск: ИВЭП ДВО РАН, 2005. С. 23.
Медведева Л. А., Никулина Т. В., Сиротский С. Е. Оценка состояния водной экосистемы Бурейского водохранилища по данным анализа фитопланктонных сообществ // Водные ресурсы. 2015. Т. 42, № 2. С. 199–211.
Мухутдинов В. Ф., Бутакова Е. А. Продукционные характеристики фитопланктона в первые годы существования Юмагузинского водохранилища // Биология внутренних вод. 2012. № 4. С. 25–30.
Определитель пресноводных беспозвоночных России и сопредельных территорий / Под ред. С. Я. Цалолихина. Т. 2: Ракообразные. СПб.: Наука, 1995. 632 c.
Полева А. О., Шкундина Ф. Б. Фитопланктон Павловского водохранилища на р. Уфа // Вестник ОГУ. 2008. № 12. С. 15–19.
Попов П. А. К прогнозу формирования ихтиоценоза Эвенкийского водохранилища // Мир науки, культуры, образования. 2009. Т. 3, № 15. С. 18–25.
Пирожников П. Л. Состояние и перспективы развития рыбного хозяйства на озерах, реках и водохранилищах СССР // Рыбное хозяйство внутренних водоемов СССР. М., 1963. 203 с.
Ресурсы поверхностных вод СССР. Гидрологическая изученность / Под ред. С. Д. Шабалина. Т. 18. Дальний Восток. Вып. 1. Амур. Л.: Гидрометеорологическое изд-во, 1966. 488 с.
Садчиков А. П. Методы изучения пресноводного фитопланктона . М.: Университет и школа, 2003. 157 с.
Саушкин Ю. Г. Географическая наука в прошлом, настоящем и будущем . М.: Просвещение, 1980. 269 с.
Смирнов Н. Н. Chydoridae фауны мира . (Новая серия / АН СССР. Зоол. ин-т. Фауна СССР. Ракообразные; № 101; Т. 1. Вып. 2). Л.: Наука, 1971. 531 с.
Ташлыкова Н. А. Экологические особенности развития фитопланктона дельтовых проток реки Селенги и сора Черкалово (оз. Байкал) : Дис. ... канд. биол. наук. Улан-Удэ, 2009. 207 с.
Шипунов А. Б., Балдин Е. М., Волкова П. А., Коробейников А. И., Назарова С. А., Петров С. В., Суфиянов В. Г. Наглядная статистика, используем R! . М.: ДМК Пресс, 2014. 298 с.
Agawin N. S. R., Duarte C. M., Agusti S. Nutrient and temperature control of the contribution pf picoplankton to phytoplankton biomass and production // Limnology and Oceanography. 2000. № 45. P. 591–600.
Bell T., Kalff J. The contribution of picoplankton in marine and freshwater system of different tropic status and depth // Limnology and Oceanography. 2001. № 46. P. 1243–1248.
Callieri C., Stockner J. G. Freshwater autotropic picophytoplankton: A review // Journal of Limnology. 2002. № 6. P. 1–14.
Condie S. A., Bormans M. The influence of density stratification on particle settling, dispersion and population growth // Journal of Theoretical Biology. 1997. Vol. 187, № 1. P. 65–75.
Descy J.-P. Phytoplankton composition and dynamics in the river Meuse (Belgium) // Archiv fur Hydrobiologie. 1987. Vol. 78. P. 225–245.
Diehl S. Phytoplankton, light, and nutrients in a gradient of mixing depths: Theory // Ecology. 2002. Vol. 83, № 2. P. 386–398.
Guiry M. D., Guiry G. M. AlgaeBase. URL: http://www.algaebase.org (дата обращения: 02.10.2018).
Jäger C. G., Diehl S., Schmidt G. M. Influence of water‐column depth and mixing on phytoplankton biomass, community composition, and nutrients // Limnology and oceanography. 2008. Vol. 53, № 6. P. 2361–2373.
Huisman J. Population dynamics of light-limited phytoplankton: Microcosm experiments // Ecology. 1999. Vol. 80. P. 202–210.
Huisman J., Arrayas M., Ebert U., Sommeijer B. How do sinking phytoplankton species manage to persist? // American Naturalist. 2002. Vol. 159. P. 245–254.
Huisman J., Thi N. N. P., Karl D. M. Sommeijer B. Reduced mixing generates oscillations and chaos in the oceanic deep chlorophyll maximum // Nature. 2006. Vol. 439. P. 322–325.
O’Brien K. R., Ivey G. N., Hamilton D. P., Waite A. M., Visser P. M. Simple mixing criteria for the growth of negatively buoyant phytoplankton // Limnology and oceanography. 2003. Vol. 48. P. 1326–1337.
Piirsoo K., Pall P., Tuvikene A., Viik M. Temporal and spatial patterns of phytoplankton in a temperate lowland river (Emajõgi, Estonia) // Journal of Plankton Research. 2008. Vol. 30, № 11. P. 1285–1295.
Potapova M., Prygiel J., Whitton B. A., Bukowska J. Use of algae for monitoring rivers in Russia // Use of Algae for Monitoring Rivers, III. Douai Agence de ĺEau Artois-Picardie, France, 1999. P. 96–101.
Ruttner-Kolisko A. Suggestions for biomass calculation of plankton rotifers. Archiv für Hydrobiologie Beihefte Ergebnisse der // Limnologie. 1977. Bd. 8. P. 71–76.
Visser P. M., Massut L., Huisman, J., Mur L. R. Sedimentation losses of Scenedesmus in relation to mixing depth // Arch. Hydrobiol. 1996. Vol. 136. P. 289–308.
WoRMS: World Register of Marine Species. 2022. URL: http://www.marinespecies.org (дата обращения: 18.11.2021).
Благодарности
Работа выполнена в рамках ФНИ (№ госрегистрации 121032200070-2) «Геоэкология водных экосистем Забайкалья в условиях современного климата и техногенеза, основные подходы к рациональному использованию вод и их биологических ресурсов». Авторы благодарят сотрудников ИПРЭК СО РАН к. г. н. И. Е. Михеева, к. б. н. Б. Б. Базарову, к. б. н. А. П. Куклина за помощь в отборе планктонных проб.

 © 2011 - 2026
© 2011 - 2026