



Введение
Период размножения занимает ведущее место в годовом жизненном цикле птиц. Сроки размножения любого вида животных устанавливались естественным отбором в процессе эволюции таким образом, чтобы потомство появлялось на свет, росло и развивалось в сезон года с оптимальными для этого физическими, защитными и кормовыми условиями. В средних широтах это весенне-летние месяцы, когда размножаются, за редким исключением, все птицы.
Механизмы, регулирующие сроки и продолжительность сезонных явлений годового жизненного цикла птиц, в том числе время начала размножения в текущем году и в конкретном географическом районе, представляют собой уникальную многоступенчатую систему воздействия на организм цирканных эндогенных гормональных ритмов, длины светового дня и других внешних факторов среды (Дольник, 1964; Murton, Kear, 1978; Dawson, 2008; Иовченко, 2015). Мигрирующие виды птиц, прибыв после зимовки к местам гнездования, сталкиваются с местными условиями среды. Метеорологические факторы могут влиять на птиц как напрямую, повышая затраты энергии на терморегуляцию при низких температурах, так и опосредованно, обусловливая развитие растительности и время появления беспозвоночных, служащих кормом для птиц. Раннее начало гнездования водоплавающих птиц в годы с теплой весной отмечают многие исследователи (Онно, 1975; Langford, Driver, 1979; Hammond, Johnson, 1984; Krapu, Reinecke, 1992; Greenwood et al., 1995; Avilova, 2016 и др.).
Продолжительность сезона откладки яиц определяется временем наступления фоторефрактерной стадии. У большинства уток, размножающихся в средних широтах северного полушария, откладка яиц прекращается в районе даты летнего солнцестояния (Murton, Kear, 1978).
В последние годы в связи с потеплением климата пристальное внимание исследователей привлекает проблема влияния глобальных изменений климата на экологию животных, в том числе птиц (McCarty, 2001; Crick, 2004; Соколов, 2010; Møller et al., 2010; Charmantier, Gienapp, 2014). Большинство публикаций посвящены воробьиным птицам и значительно меньше гусеобразным. В них сообщается, что изменение климата в некоторой степени обусловливает сдвиги в датах миграции и сроках начала размножения (Žalakevičius et al., 2009; Guillemain et al., 2013; Arzel et al., 2014; Clark et al., 2014; Pavón-Jordán et al., 2017).
При отклонении сроков размножения от оптимального у уток может изменяться средняя величина кладки (Krapu et al., 2004; Михантьев, Селиванова, 2008), успех гнездования (Юрлов и др., 1994; Drever, Clark, 2007; Grant, Shaffer, 2012) и выживаемость потомства (Dzus, Clark, 1998; Blums et al., 2002; Gurney et al., 2012). Это отражается на продуктивности популяции и, следовательно, на ее численности.
Цель настоящего исследования – на большом эмпирическом материале показать закономерности межгодовой вариабельности сроков гнездования кряквы на юге Западной Сибири, а именно выявить предельные параметры и возможные долговременные тренды сроков начала и длительности периода гнездования кряквы и оценить влияние локальных погодных условий (даты перехода среднесуточной температуры через 0 °С) и глобальных климатических процессов (Североатлантической осцилляции, NAO) на межгодовые вариации времени начала гнездования.
Материалы
Исследования проводили с 1970 по 2018 г. за исключением 6 лет (1976, 1977, 2008, 2012, 2013, 2014) в Северной Кулунде. Экологию водоплавающих птиц изучали на оз. Кротовая Ляга (53°43' с. ш., 77°53' в. д.) площадью 345–485 га в зависимости от обводненности, входящем в озерно-займищную систему в низовьях бассейна р. Карасук. Подробно место работы описано ранее (Михантьев, Селиванова, 2008).
В своих анализах мы использовали метеорологические данные Карасукской метеостанции, расположенной в 10 км от места работы. Значения индексов Североатлантической осцилляции (NAO) взяты с сайта американского Центра атмосферных исследований Джеймса У. Херрелла (Hurrell, 2020).
Методы
Учеты утиных гнезд проводили ежегодно с начала мая до конца июля на участках общей площадью примерно 28 га. Места учетов включали в себя массивы сплавины и ее многочисленные островки размером от одного до нескольких десятков квадратных метров на плесах в зарослях тростника (Phragmites australis) и рогоза узколистного (Typha angustifolia). Периодичность повторных учетов и проверок гнезд 7–10 дней. Обнаружено 2310 гнезд кряквы. Учтены как основные, так и, вероятно, повторные кладки. Уровень гибели кладок ежегодно оценивали процентом неуспешных от общего количества кладок с известной судьбой. Также рассчитывали долю гнезд, погибших на стадии откладки яиц. На гибель гнезд на этой стадии указывало отсутствие пуха, наличие ненасиженных яиц, скорлупы с остатками белка, желтка, подскорлуповой оболочкой свежих яиц.
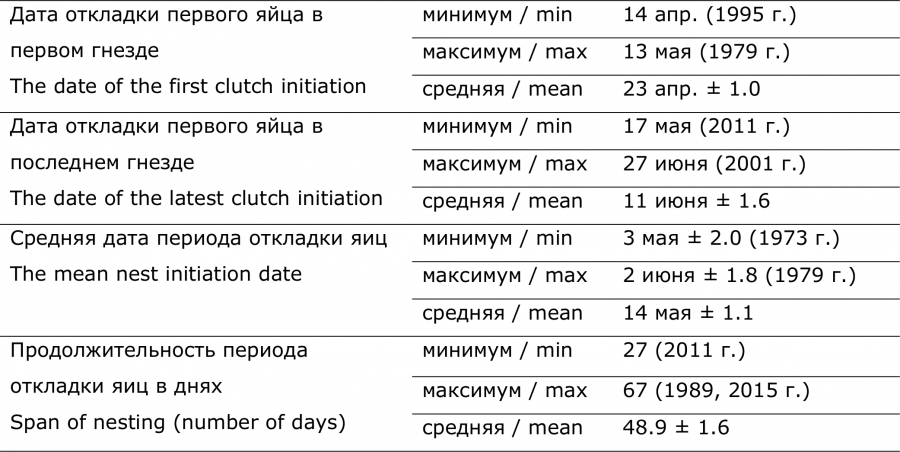
Дату откладки утками первого яйца определяли стандартными расчетами (Меднис, 1968). Для гнезд, обнаруженных во время формирования кладки, эту дату вычисляли, отнимая от даты обнаружения гнезда число яиц на этот день. Обычно утки несутся ежедневно и начинают насиживать в день откладки последнего яйца. Для гнезд, в которых были известны сроки выклева утят, дату откладки первого яйца определяли по величине кладки и средней продолжительности насиживания, которая равна для кряквы 26 дням (Исаков, 1952; Михантьев, Селиванова, 2005). В тех случаях, когда гнезда обнаруживали во время насиживания, дату откладки первого яйца вычисляли при помощи неоднократного использования методики определения степени насиженности яиц по их плавучести в воде (Westerskov, 1950; Меднис, Блум, 1976). Гнезда, разоренные или брошенные наседками задолго до их обнаружения, для характеристики сроков размножения не использовали. В общей сложности сроки начала откладки яиц были определены в 1879 гнездах кряквы, найденных на стадии откладки яиц (28.1 %), насиживания (66.2 %) и во время выклева утят (5.7 %).
Для характеристики сроков размножения в каждом конкретном году использовали дату начала формирования первой кладки, среднюю дату периода откладки яиц и дату начала формирования последней кладки. Период откладки яиц, или период появления кладок, это отрезок времени, в течение которого появлялись новые гнезда. Его продолжительность равна количеству дней между датами начала самой ранней и самой поздней кладки. Среднюю дату и длительность периода откладки яиц рассчитывали только в те годы, когда дата снесения первого яйца была установлена не менее чем в 10 гнездах. Таких было 42 года.
Календарные даты для расчетов представляли в числовом формате (1 января = 1). Статистическая оценка результатов проведена с использованием пакета программ PAST_3.17 (Hammer et al., 2001): проверка на нормальное распределение (тест Шапиро – Вилка), проверка на наличие тренда (тест Манна – Кендалла), параметрические и непараметрические анализы. Для проверки различий средних значений применяли U-критерий Манна – Уитни. Для измерения степени сопряженности различных показателей использовали метод корреляции Пирсона (r), ранговой корреляции Спирмена (rs) и метод множественной регрессии. Также для статистических анализов и построения графиков использовали Excel 2010 (Microsoft). Все средние величины приведены с ошибкой (± SE).
Результаты
Наши многолетние наблюдения показали, что период откладки яиц в разные годы может начинаться при разном фотопериоде (длина светового дня + гражданские сумерки). Самая ранняя кладка была начата в 1995 г. при фотопериоде 15 ч. 15 мин. Самое позднее начало гнездования зарегистрировано в 1979 г. при фотопериоде 17 ч. 17 мин. Средней многолетней дате начала гнездования соответствует фотопериод 15 ч. 49 мин. Таким образом, диапазон продолжительности фотопериода достигает двух часов. Такой широкий разброс свидетельствует, что время начала размножения и другие характеристики репродуктивного периода контролируются не только эндогенным ритмом с фотопериодом, но и другими факторами среды. Начало, окончание и общая продолжительность периода откладки яиц варьировали в широких пределах. Разница крайних значений достигала 30 дней (табл. 1).
Таблица 1. Сроки гнездования кряквы в Северной Кулунде (оз. Кротовая Ляга, 1970–2018 гг.)
Если рассматривать данные за весь период работы (1970–2018 гг.), то смещение сроков установления положительных весенних температур и сроков гнездования кряквы на более ранние даты не проявляется (рис. 1). Их тренды не достоверны (Z < 1.3; p > 0.2). Однако, по наблюдениям за менее продолжительный период времени, начиная с 1979 г. переход среднесуточной весенней температуры воздуха через 0 °C постепенно смещался на более раннее время (Z = 2.7; p = 0.007) в среднем на 2.5 ± 0.8 дня за каждые 10 лет. Но у сроков гнездования на этом промежутке времени такая тенденция отсутствует.
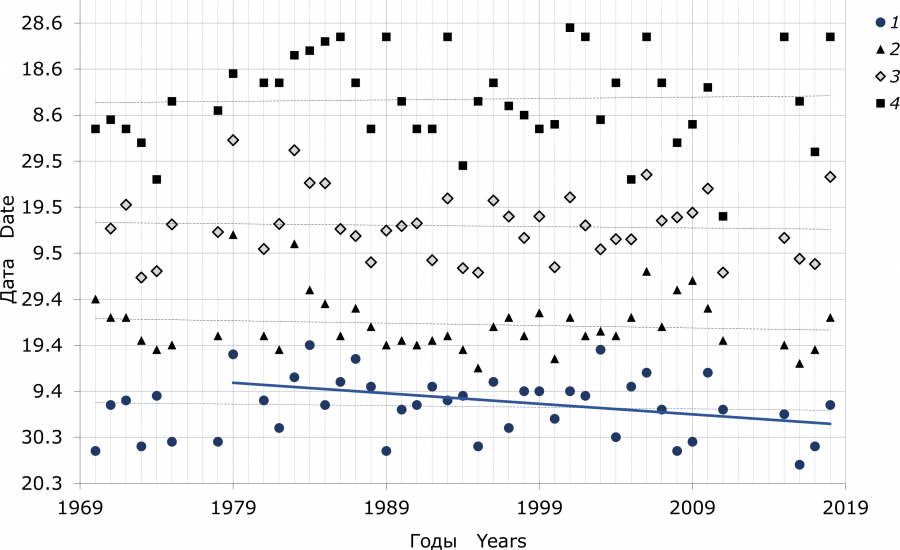
Рис. 1. Вариабельность дат периода откладки яиц кряквы и сроков установления положительных среднесуточных температур воздуха весной (оз. Кротовая Ляга, 1970–2018 гг.). Сплошная линия – достоверный тренд, пунктирные линии – недостоверные тренды. 1 – дата перехода среднесуточной весенней температуры воздуха через 0 °C, 2 – дата начала первой кладки, 3 – средняя дата периода откладки яиц, 4 – дата начала последней кладки
Fig. 1. Variation in the timing of egg-laying period of mallards, and terms of establishing positive average daily air temperatures in spring ( Lake Krotovaya Lyaga, 1970–2018). Solid line – the significant trend, dashed lines – not significant trends. 1 – date of transition of the average daily air temperature at 0 °C , 2 – the first egg-laying date, 3 – average date of the egg-laying period, 4 – date of the latest clutch initiation
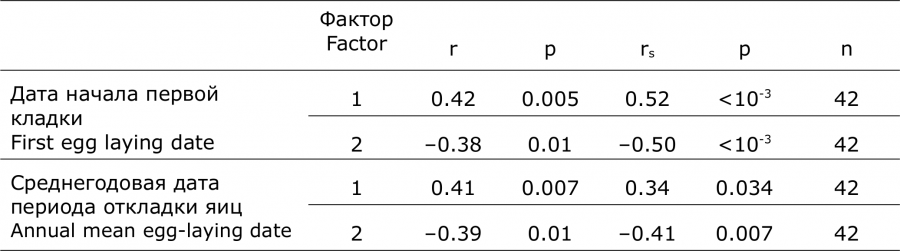
Колебания средней даты периода откладки яиц по годам зависели сильнее от даты появления первых кладок (R2 = 0.62; F[1,40] = 67.5; p < 10-3), чем последних (R2 = 0.29; F[1,40] = 16.3; p < 10-3). Анализ выявил высоко достоверную положительную связь сроков размножения с наступлением устойчивой теплой местной погоды – с датой перехода среднесуточных температур воздуха через 0 °С, и достоверно отрицательную с февральским индексом NAO. При этом местный погодный показатель и индекс NAO коррелировали между собой (r = – 0.30, p = 0.04, 1970–2018 гг.). Поскольку распределения многолетних рядов данных (индексов Североатлантической осцилляции, дат перехода среднесуточной температуры через 0 °C и средних дат периода откладки яиц) соответствуют нормальному (W > 0.95; p > 0.12), а данные по датам начала первой кладки ему не соответствуют (W = 0.89; p < 10-3), корреляционный анализ провели двумя методами: Пирсона (r) и Спирмена (rs) (табл. 2, рис. 2). Ежегодные колебания числа гнезд кряквы на озере не отражались на сроках начала ее гнездования.
Таблица 2. Коэффициенты корреляции (r Пирсона и rs Спирмена) сроков гнездования кряквы с: 1 – датой перехода среднесуточной температуры через 0 °С и 2 – февральским индексом Североатлантической осцилляции (оз. Кротовая Ляга, 1970–2018 гг.)
Мы оценили с помощью метода множественной регрессии зависимость даты появления первой кладки и средней даты периода откладки яиц от совокупного влияния двух факторов: даты перехода среднесуточной температуры через 0 °С и февральского индекса NAO. Суммарный эффект этих факторов на сроки гнездования кряквы достаточно существенный и почти одинаковый: на дату начала гнездования R2 = 0.253, F[2,39] = 6.59, p = 0.003 и на среднюю дату периода откладки яиц R2 = 0.251, F[2,39] = 6.54, p = 0.003. То есть суммарное влияние этих двух факторов обусловливает 25 % изменчивости сроков размножения кряквы.
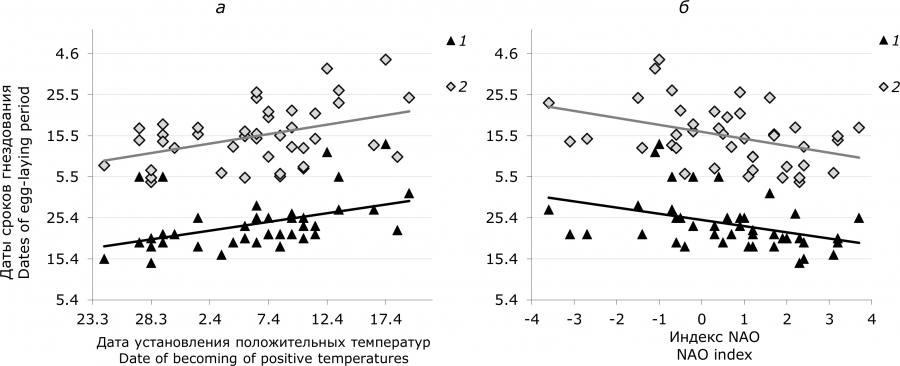
Рис. 2. Зависимость сроков гнездования кряквы от даты перехода среднесуточной весенней температуры воздуха через 0 °C (а) и февральского индекса Североатлантической осцилляции (б), оз. Кротовая Ляга, 1970–2018 гг. Прямые соответствуют линиям простой линейной регрессии. 1 – дата начала первой кладки, 2 – средняя дата периода откладки яиц
Fig. 2. The timing of egg-laying period of mallards in dependence to the date of transition of average daily spring air temperature at 0°C (a) and February North Atlantic Oscillation index (б), Lake Krotovaya Lyaga, 1970–2018. The lines indicate the simple linear regression. 1 – the first egg-laying date, 2 – average date of egg-laying period
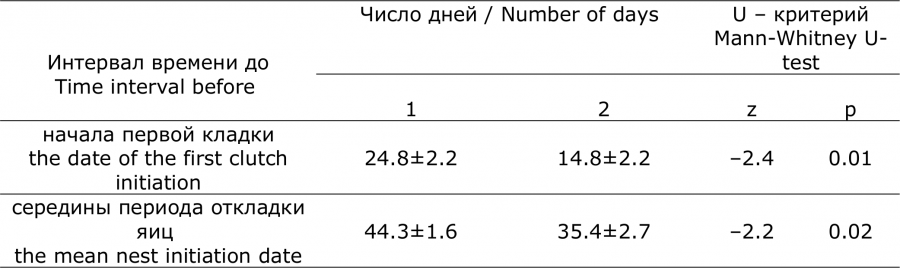
При сдвиге перехода среднесуточной температуры через 0 °C в раннюю или позднюю сторону дата появления первых гнезд и средняя дата периода откладки яиц сдвигаются на меньшее число дней. Коэффициент, отражающий угол наклона линии регрессии, меньше единицы (0.42 ± 0.11, р = 0.0008 и 0.46 ± 0.13, р = 0.006 соответственно) (см. рис. 2а). Следовательно, разница между сроками начала размножения и сроками наступления положительных среднесуточных температур уменьшается по мере запаздывания весны, и наоборот, увеличивается при раннем ее наступлении. Это правило четко проявляется при сравнении 10 лет с наиболее ранней весной, когда положительные среднесуточные температуры устанавливались в среднем 28 марта (SE = 0.7 дня) и 10 лет с наиболее поздней весной, когда положительные среднесуточные температуры устанавливались в среднем 13 апреля (SE = 1.1 дня). Временной отрезок между датой перехода среднесуточной температуры воздуха через 0 °C и датой появления первого гнезда в годы с ранней весной был достоверно продолжительнее, чем в годы с поздней весной. Такая же закономерность установлена и для временного интервала между датой перехода температуры воздуха через 0 °C и средней датой периода появления кладок (табл. 3).
Таблица 3. Сравнение временных интервалов между датой перехода среднесуточных температур воздуха через 0 °C и началом гнездования кряквы в годы с ранней (1) и поздней (2) весной (оз. Кротовая Ляга, 1970–2018 гг.)
Продолжительность видового сезона откладки яиц соответствует промежутку времени между началом самой ранней и самой поздней кладки кряквы за все время наблюдений и равна 75 дням (14 апреля – 27 июня, см. табл. 1). В каждом конкретном году длительность периода откладки яиц определялась датой начала первой кладки (rs = – 0.32, p = 0.03) и, в большей степени, но с противоположным знаком, последней (rs = 0.79, p < 10-3). Суммарное влияние других факторов на растянутость периода откладки яиц: численности гнезд на озере, температурных условий весны, гибели гнезд на стадии откладки яиц (R2 = 0.22, F[3,38] = 3.65, p = 0.02) достоверно, но из них только число гнезд имело основное влияние (rs = 0.48, p = 0.001).
Обсуждение
Мигрирующие виды птиц в течение своего годового жизненного цикла обитают в разных регионах, где подвергаются влиянию климатических и других условий среды, характерных для определенной местности. Условия существования на одном этапе цикла и их последствия для организмов мигрантов могут тем или иным образом сказываться на последующих этапах (Sedinger, Alisauskas, 2014). В частности, физическое состояние перелетных птиц в зимний период, обусловленное доступностью кормов, может повлиять на даты прилета на места размножения (Vähätalo et al., 2004; Balbontin et al., 2009; Rockwell et al., 2012) и даты откладки яиц (McKellar et al., 2013). Получение и накопление энергетических запасов для размножения обычно происходит в конце зимы или в начале весны перед размножением (Whyte et al., 1986; Dubovsky, Kaminski, 1994). Влияние состояния тела на сроки и успешность размножения отмечено у уток, гнездящихся не только в арктическом (Fox, 1996), но и в умеренном поясе (Pattenden, Boag, 1989; Alisauskas, Ankney, 1992; Dubovsky, Kaminski, 1994; Alisauskas, Devink, 2015). Самки кряквы, прилетевшие с зимовки более упитанными, начинали гнездиться примерно на 15 дней раньше, чем менее упитанные (Devries et al., 2008).
Кряквы, размножающиеся на юге Западной Сибири, мигрируют в юго-западном направлении и зимуют на юго-западе Азии и юго-востоке Европы (Veen et al., 2005). Погодно-гидрологические факторы в этих регионах, а с ними и другие условия среды, необходимые для существования и успешной подготовки крякв к миграции и последующему размножению, находятся под влиянием Североатлантической осцилляции (Нестеров, 2001; Hurrell et al., 2003). Состояние организмов уток, прибывающих в район размножения, зависит как от кормовых условий на зимовке, так и от возможности пополнять энергетические ресурсы на путях пролета. Кряква относится к видам, делающим кладку не только за счет накопленных внутренних резервов, но в значительной степени за счет ресурсов, получаемых на местах гнездования. Поэтому и доступность, и качество кормов здесь определяют скорость восполнения внутренних запасов в промежуток времени между прилетом и началом гнездования. Чем раньше и теплее весна, тем благоприятнее условия питания. Следовательно, начало откладки яиц определяется условиями существования уток и на зимовках, и в местах размножения (начало гнездования сопряжено и с индексом NAO, и с локальными погодными условиями).
Климат, как следствие глобальных геофизических процессов, изменялся во времени то в сторону похолодания, то в сторону потепления. За последние 40–50 лет происходит его потепление, оказывающее влияние планетарного масштаба на биосферу в целом и на популяции диких животных в частности. В течение следующих двух десятилетий предсказывают увеличение глобальной средней температуры приземного воздуха на 0.3–0.7 °C и увеличение осадков в средних и высоких широтах (Kirtman et al., 2014). Для многих организмов, включая птиц (Parmesan, Yohe, 2003; Соколов, 2010), в ответ на потепление климата зарегистрированы сдвиги в фенологии. У некоторых видов уток отмечают более ранний отлет с зимовок и прилет на места гнездования весной (Vähätalo et al., 2004; Murphy-Klassen et al., 2005; Rainio et al., 2006; Žalakevičius et al., 2009; Guillemain et al., 2013). Примерно в 60 % исследований долгосрочное смещение сроков размножения в раннюю сторону объясняют глобальным потеплением (Dunn, 2004). Многие виды птиц стали откладывать яйца раньше в годы с более теплой весной.
Однако по другим видам данные противоречивы. Межгодовые различия сроков наступления сезонных процессов у уток настолько велики, что приспособительный характер незначительных долгосрочных изменений во времени на фоне глобального потепления некоторые авторы считают сомнительным. Так, на севере Финляндии за 1989−2006 гг. долговременные тренды в хронологии периода гнездования кряквы и обыкновенного гоголя не обнаружены (Oja, Pöysä, 2007). Но, по данным К. В. Авиловой, в Москве за 18-летний период наблюдений (1998–2015 гг.) кряква стала приступать к гнездованию значимо раньше. Причиной этому, кроме потепления, могла стать и постоянно растущая освещенность города в темное время суток (Avilova, 2016). Есть мнение, что реакция видов на изменение климата имеет географические различия и может изменяться со временем (Dunn, 2004). В Северной Кулунде за период 1970–2018 гг. значимого смещения сроков гнездования кряквы на более раннее время не обнаружено.
Многие исследователи отмечают, что разница между временем начала размножения уток и наступлением тепла увеличивается с более ранним приходом весны, о котором судят либо по дате таяния льда, либо по дате установления положительных среднесуточных температур (Oja, Pöysä, 2007; Arzel et al., 2014; Avilova, 2016). Результаты наших наблюдений согласуются с выводами этих авторов. Чем раньше весной наступали устойчивые положительные среднесуточные температуры воздуха, тем длиннее был временной интервал от этой даты до начала размножения кряквы. До появления первых гнезд он увеличивался в среднем на 5.0 ± 1.2 дня, а до среднегодовой даты периода откладки яиц на 4.8 ± 1.4 дня на каждые 10 дней смещения даты перехода среднесуточной весенней температуры через 0 °C на ранние сроки.
Фактический ход размножения в конкретном году определяется многими причинами: возрастным составом популяции, различиями физиологического состояния отдельных особей, территориальным поведением, условиями зимовки, погодой в период, предшествующий гнездованию, и в ходе гнездования. Продолжительность периода начала кладок кряквой в годы с ранней весной и ранним началом гнездования была больше, чем в годы с поздней весной. Возможно, это связано с усилением в таких условиях разобщенности во времени гнездования взрослых и впервые размножающихся молодых самок, часто откладывающих яйца позже взрослых особей. Удлинение периода гнездования при высокой численности пар происходило, вероятно, вследствие обострения конкуренции за территорию, т. к. крякве присуще территориальное поведение, препятствующее одновременному гнездованию всей популяции, расчленяющее период гнездования на отдельные волны. Кроме этого, мы предполагали, что высокая гибель кладок может быть причиной повышения частоты повторного гнездования и, как следствие, приводить к удлинению сезона откладки яиц. Однако анализ наших данных не подтвердил эту гипотезу.
Заключение
Итак, в ходе многолетних исследований (с 1970 по 2018 г.) определены пороговые даты начала и окончания периода откладки яиц кряквы на юге Западной Сибири. Долговременного смещения сроков размножения кряквы в связи с потеплением климата мы не обнаружили. Локальные погодные условия весны и глобальные атмосферные осцилляции оказывают существенное влияние на гнездование кряквы. В годы с повышенной весенней температурой и высоким зимним индексом NAO оно начиналось достоверно раньше. В связи с прогнозом дальнейшего потепления климата исследования по фенологии птиц следует продолжить.
Библиография
Дольник В. Р. О механизме фотопериодического контроля эндогенного ритма половой цикличности птиц // Зоологический журнал. 1964. Т. 43. Вып. 5. С. 720–733.
Иовченко Н. П. Фотопериодический контроль годовых циклов у птиц: современные представления и перспективы изучения // Энергетика и годовые циклы птиц (памяти В. Р. Дольника): Материалы междунар. конф. М.: Т-во научных изданий КМК, 2015. С. 153–159.
Исаков Ю. А. Подсемейство утки // Птицы Советского Союза Т. 4 / Под ред. Г. П. Дементьев, Н. А. Гладков. М.: Советская наука, 1952. С. 344–635.
Меднис А. А. Биология гнездования уток на озере Энгурес // Экология водоплавающих птиц Латвии / Под ред. Х. А. Михельсона. Рига: Зинатне, 1968. С. 85–108.
Меднис А. А., Блум П. Н. Отлов насиживающих уток и их птенцов // Кольцевание в изучении миграций птиц фауны СССР / Под ред. В. Д. Ильичева. М.: Наука, 1976. С. 157–167.
Михантьев А. И., Селиванова М. А. Вариабельность длительности периода инкубации у уток // Актуальные вопросы изучения птиц Сибири: Материалы Сибирской орнитологической конф. Барнаул, 2005. С. 94–98.
Михантьев А. И., Селиванова М. А. Вариации величины кладки у уток // Сибирский экологический журнал. 2008. Т. 15, № 1. С. 187–194.
Нестеров Е. С. Низкочастотная изменчивость циркуляции атмосферы и уровень Каспийского моря во второй половине ХХ века // Метеорология и гидрология. 2001. № 11. С. 27–36.
Онно С. Время гнездования у водоплавающих и прибрежных птиц в Матсалуском заповеднике (Эстонская ССР) // Сообщение Прибалтийской комиссии по изучению миграций птиц. № 8. Тарту, 1975. С. 107–155.
Соколов Л. В. Климат в жизни растений и животных . СПб.: Тесса, 2010. 344 с.
Юрлов А. К., Михантьев А. И., Селиванова М. А. Влияние факторов среды на продуктивность популяций околоводных птиц на юге Западной Сибири // Сибирский экологический журнал. 1994. Т. 1, № 4. C. 347–353.
Alisauskas R. T., Ankney C. D. The cost of egg laying and its relationship to nutrient reserves in waterfowl // Ecology and Management of Breeding Waterfowl / B. D. J. Batt, A. D. Afton, M. G. Anderson, C. D. Ankney, D. H. Johnson, J. A. Kadlec, G. L. Krapu (eds.). Minneapolis: University of Minnesota Press., 1992. P. 30–61.
Alisauskas R. T., Devink J.-M. Breeding costs, nutrient reserves, and cross-seasonal effects: dealing with deficits in sea ducks // Ecology and Conservation of North American Sea Ducks. Studies in Avian Biology N. 46. CRC Press, Boca Raton, FL, 2015. P. 125–168.
Arzel C., Dessborn L., Pöysä H., Elmberg J., Nummi P., Sjöberg K. Early springs and breeding performance in two sympatric duck species with different migration strategies // Ibis. 2014. Vol. 156, № 2. P. 288–298. DOI: 10.1111/ibi.12134
Avilova K. V. The life cycle and number dynamics of urban mallard population (Anas platyrhynchos, Anseriformes, Aves) in Moscow // Biology Bulletin. 2016. Vol. 43, № 9. P. 1212–1224. DOI: 10.1134/S1062359016110029
Balbontin J., Møller A. P., Hermosell I. G., Marzal A., Reviriego M., de Lope F. Individual responses in spring arrival date to ecological conditions during winter and migration in a migratory bird // Journal of Animal Ecology. 2009. Vol. 78, № 5. P. 981–989. DOI: 10.1111/j.1365-2656.2009.01573.x
Blums P., Clark R. G., Mednis A. Patterns of reproductive effort and success in birds: path analyses of long-term data from European ducks // Journal of Animal Ecology. 2002. Vol. 71, № 2. P. 280–295. DOI: 10.1046/j.1365-2656.2002.00598.x
Charmantier A., Gienapp P. Climate change and timing of avian breeding and migration: evolutionary versus plastic changes // Evolutionary Applications. 2014. Vol. 7, № 1. P. 15–28. DOI: 10.1111/eva.12126
Clark R. G., Pöysä H., Runko P., Paasivaara A. Spring phenology and timing of breeding in short-distance migrant birds: phenotypic responses and offspring recruitment patterns in common goldeneyes // Journal of Avian Biology. 2014. Vol. 45, № 5. P. 457–465. DOI: 10.1111/jav.00290
Crick H. Q. P. The impact of climate change on birds // Ibis. 2004. Vol. 146, Suppl 1. P. 48–56.
Dawson A. Control of the annual cycle in birds: endocrine constraints and plasticity in response to ecological variability // Philosophical Transactions of the Royal Society of London Series B – Biological Sciences. 2008. № 1497. P. 1621–1633. DOI: 10.1098/rstb.2007.0004
Devries J. H., Brook R. W., Howerter D. W., Anderson M. G. Effects of spring body condition and age on reproduction in Mallards (Anas platyrhynchos) // Auk. 2008. Vol. 125, № 3. P. 618–628. DOI: 10.1525/auk.2008.07055
Drever M. C., Clark R. G. Spring temperature, clutch initiation date and duck nest success: a test of the mismatch hypothesis // Journal of Animal Ecology. 2007. Vol. 76, № 1. P. 139–148. DOI: 10.1111/j.1365-2656.2006.01183.x
Dubovsky J. A., Kaminski R. M. Potential reproductive consequences of winter-diet restriction in Mallards // Journal of Wildlife Management. 1994. Vol. 58, № 4. P. 780–786. DOI: 10.2307/3809693
Dunn P. Breeding dates and reproductive performance // Birds and Climate Change. Advances in Ecological Research. Vol. 35 / A. P. Møller, W. Fiedler, P. Berthold (eds.). San Diego: Elsevier Ltd, 2004. P. 69–87.
Dzus E. H., Clark R. G. Brood survival and recruitment of mallards in relation to wetland density and hatching date // Auk. 1998. Vol. 115, № 2. P. 311–318. DOI: 10.2307/4089189
Fox A. D. Factors affecting the reproductive output northern hemisphere migratory Anatidae: The role of female conditions (a review) // Anatidae 2000: An International Conference on the Conservation, Habitat Management and Wise Use of Ducks, Geese and Swans. Gibier Faune Sauvage. 1996. Vol. 13, № 2. P. 635–651.
Grant T. A., Shaffer T. L. Time-specific patterns of nest survival for ducks and passerines breeding in North Dakota // Auk. 2012. Vol. 129, № 2. P. 319–328. DOI: 10.1525/auk.2012.11064
Greenwood R. J., Sargeant A. B., Johnson D. H., Cowardin L. M., Shaffer T. L. Factors associated with duck nest success in the Prairie Pothole Region of Canada // Wildlife Monographs. 1995. № 128. 57 p.
Guillemain M., Pöysä H., Fox A. D., Arzel C., Dessborn L., Ekroos J., Gunnarsson G., Holm T. E., Christensen T. K., Lehikoinen A., Mitchell C., Rintala J., Møller A. P. Effects of climate change on European ducks: what do we know and what do we need to know? // Wildlife Biology. 2013. Vol. 19, № 4. P. 404–419. DOI: 10.2981/12-118
Gurney K. E. B., Clark R. G., Slattery S. M. Seasonal variation in pre-fledging survival of lesser scaup Aythya affinis: hatch date effects depend on maternal body mass // Journal of Avian Biology. 2012. Vol. 43, № 1. P. 68–78. DOI: 10.1111/j.1600-048X.2011.05490.x
Hammer Ø., Harper D. A. T., Ryan P. D. PAST: Paleontological Statistics Software Package for Education and Data Analysis // Palaeontologia Electronica. 2001. Vol. 4, № 1, art. 4. 9 p. URL: http://palaeo-electronica.org/2001_1/past/issue1_01.htm. http://folk.uio.no/ohammer/past.
Hammond M. C., Johnson D. H. Effects of Weather on Breeding Ducks in North Dakota // Fish and Wildlife Technical Report N 1. US Department of the Interior, Fish and Wildlife Service, Washington, DC, 1984. 19 p.
Hurrell J. W., Kushnir Y., Ottersen G., Visbeck M. An overview of the North Atlantic Oscillation // The North Atlantic oscillation: climate significance and environmental impact. Geophysical Monograph Series. 2003. Vol. 134. P. 1–35. DOI: 10.1029/134GM01
Hurrell J, National Center for Atmospheric Research Staff (eds). The Climate Data Guide: Hurrell North Atlantic Oscillation (NAO) Index (station-based). Retrieved from https://climatedataguide.ucar.edu/climate-data/hurrell-north-atlantic-oscillation-nao-index-station-based. Last modified 24 Apr 2020.
Kirtman S. B., Power S. B., Adedoyin A. J., Boer G. J., Bojariu R., Camilloni I., Doblas-Reyes F. J., Fiore A. M., Kimoto M., Meehl G. A. Near-term climate change: Projections and predictability // Intergovernmental Panel on Climate Change. Cambridge University Press, 2014. P. 953–1028. DOI: 10.1017/CBO9781107415324.023
Krapu G. L., Reinecke K. J. Foraging ecology and nutrition // Ecology and Management of Breeding Waterfowl / B. D. J. Batt, A. D. Afton, M. G. Anderson, C. D. Ankney, D. H. Johnson, J. A. Kadlec, G. L. Krapu (eds.). Minneapolis: University of Minnesota Press., 1992. P. 1–29.
Krapu G. L., Reynolds R. E., Sargeant G. A., Renner R. W. Patterns of variation in clutch sizes in a guild of temperate-nesting dabbling ducks // Auk. 2004. Vol. 121, № 3. P. 695–706. DOI: 10.1642/0004-8038(2004)1212.0.CO;2
Langford W. A., Driver E. A. Quantification of the relationship between Mallard nest initiation and temperature // Wildfowl. 1979. № 30. P. 31–34.
McCarty J. P. Ecological consequences of recent climate change // Conservation Biology. 2001. Vol. 15, № 2. P. 320–331. DOI: 10.1046/j.1523-1739.2001.015002320.x
McKellar A. E., Marra P. P., Hannon S. J., Studds C. E., Ratcliffe L. M. Winter rainfall predicts phenology in widely separated populations of a songbird // Oecologia. 2013. Vol. 172, № 2. P. 595–605. DOI: 10.1007/s00442-012-2520-8
Møller A. P., Fiedler W., Berthold P. (eds.). Effects of climate change on birds. Oxford University Press, 2010. 321 p.
Murphy-Klassen H. M., Underwood T. J., Sealy S. G., Czyrnyj A. A. Long-term trends in spring arrival dates of migrant birds at Delta Marsh, Manitoba, in relation to climate change // Auk. 2005. Vol. 122, № 4. P. 1130–1148. DOI: 10.1642/0004-8038(2005)1222.0.CO;2
Murton R. K., Kear J. Photoperiodism in waterfowl: phasing of breeding cycles and zoogeography // Journal of Zoology, Lond. 1978. Vol. 186, № 2. P. 243–283. DOI: 10.1111/j.1469-7998.1978.tb03368.x
Oja H., Pöysä H. Spring phenology, latitude, and the timing of breeding in two migratory ducks: implications of climate change impacts // Annales Zoologici Fennici. 2007. № 44. P. 475–485.
Parmesan C., Yohe G. A globally coherent fingerprint of climate change impacts across natural systems // Nature. 2003. Vol. 421. P. 37–42.
Pattenden R. K., Boag D. A. Effects of body mass on courtship, pairing, and reproduction in captive Mallards // Canadian Journal of Zoology. 1989. Vol. 67, № 2. P. 495–501. DOI: 10.1139/z89-072
Pavón-Jordán D., Santangeli A., Lehikoinen A. Effects of flyway-wide weather conditions and breeding habitat on the breeding abundance of migratory boreal waterbirds // Journal of Avian Biology. 2017. Vol. 48, № 7. P. 988–996. DOI: 10.1111/jav.01125
Rainio K., Laaksonen T., Ahola M., Vähätalo A. V., Lehikoinen E. Climatic response in spring migration of boreal and arctic birds in relation to wintering area and taxonomy // Journal of Avian Biology. 2006. Vol. 37, № 5. P. 507–515. DOI: 10.1111/j.0908-8857.2006.03740.x
Rockwell S. M., Bocetti C. I., Marra P. P. Carry-over effects of winter climate on spring arrival date and reproductive success in an endangered migratory bird, Kirtland’s Warbler (Setophaga kirtlandii) // Auk. 2012. Vol. 129, № 4. P. 744–752. DOI: 10.1525/auk.2012.12003
Sedinger J. S., Alisauskas R. T. Cross-seasonal effects and the dynamics of waterfowl populations // Wildfowl. 2014. Special Issue 4. P. 277–304.
Vähätalo A., Rainio K., Lehikoinen A., Lehikoinen E. Spring arrival of birds depends on the North Atlantic Oscillation // Journal of Avian Biology. 2004. Vol. 35, № 3. P. 210–216. DOI: 10.1111/j.0908-8857.2004.03199.x
Veen J., Yurlov A. K., Delany S. N., Mihantiev A. I., Selivanova M. A., Boere G. C. An atlas of movements of Southwest Siberian waterbirds. Wetlands International, Wageningen, The Netherlands, 2005. 60 p.
Westerskov K. Methods for Determining the age of game bird eggs // The Journal of Wildlife Management. 1950. Vol. 14, № 1. P. 56–67. DOI: 10.2307/3795978
Whyte R. J., Baldassarre G. A., Bolen E. G. Winter condition of mallards on the southern High Plains of Texas // The Journal of Wildlife Management. 1986. Vol. 52, № 1. P. 52–57. DOI: 10.2307/3801487
Žalakevičius M., Bartkevičienė G., Ivanauskas F., Nedzinskas V. The response of spring arrival dates of non-passerine migrants to climate change: A case study from Eastern Baltic // Acta Zoologica Lituanica. 2009. Vol. 19, № 3. P. 155–171. DOI: 10.2478/v10043-009-0029-0
Благодарности
Настоящее многолетнее исследование было бы невозможно без Карасукского озерного стационара ИСиЭЖ СО РАН, созданного профессором С. С. Фолитареком. Заведующие стационаром Д. В. Столяров и В. А. Шило и весь коллектив оказывали нам всяческую техническую помощь в работе и организации быта. Большую помощь в сборе полевого материала в разные годы оказывали студенты-дипломники и аспиранты. Всем им мы приносим свою искреннюю благодарность.
Исследование поддержано Программой фундаментальных научных исследований (ФНИ) государственных академий наук на 2013–2020 гг., проект № VI.51.1.8 (АААА-А16-116121410118-7).

 © 2011 - 2026
© 2011 - 2026