



Введение
Ареалы ужеобразных змей рода Natrix Laurenti, 1768 охватывают все расположенные на территории Волжского бассейна 39 субъектов Российской Федерации и две области Казахстана (Табачишин, Табачишина, 2002; Чугуевская, 2005; Бакиев и др., 2009). Имеются обширные сведения по морфологии, распространению, численности, плотности, размерно-половой структуре популяций, размножению, сезонной и суточной активности, термобиологии, питанию, паразитам и потребителям ужей рода Natrix в Волжском бассейне (Bakiev et al., 2011; Litvinov et al., 2011; Кленина, Бакиев, 2015; Кленина и др., 2015). Установлено, что змеи имеют вполне развитую кроветворную и иммунную системы и демонстрируют реакции на весь спектр экологических факторов, характерных для среды их обитания (Davis et al., 2008; Kobolkuti et al., 2012; Lisicic et al., 2013). Кровь змей имеет много общего, с одной стороны, с кровью рыб и амфибий, с другой – птиц и млекопитающих (Cooper et al., 1985), что делает их уникальным объектом для экологических исследований. Известно, что клеточные механизмы неспецифической резистентности у рептилий представлены набором лейкоцитов: моноцитами, базофилами, эозинофилами и гетерофилами, реакции адаптивного иммунного ответа обеспечиваются лимфоцитами (Davis et al., 2008; Arican, Cicek, 2010; Павлов, Юсупов, 2015; Васильев, 2016 и др.). Получены популяционные лейкоцитарные формулы крови ужа обыкновенного и ужа водяного Самарской области. Выявлены межвидовые различия, происходящие в крови этих змей до и после откладывания яиц, связанные с активацией естественного иммунитета животных (Романова и др., 2015). Выявлены различия в лейкоцитарном составе крови ядовитых и неядовитых змей (Романова и др., 2017). В настоящее время антропогенная трансформация местообитаний имеет глобальный характер, при этом индивидуальная аккомодация на изменяющиеся экологические условия среды во многом определяется способностью организма нормализовать иммуногематологическую систему гомеостаза. В этой связи актуальность и дальнейшие перспективы изучения адаптивных реакций системы крови и механизмов регуляции защитных функций организма змей не вызывают сомнений и необходимы как для теории, так и для решения практических природоохранных вопросов в области экологии отдельных видов.
Целью работы являлась сравнительная оценка показателей лейкоцитарной системы крови ужа водяного (Natrix tessellata Laurenti, 1768) популяций Северного и Восточного Прикаспия в разные периоды сезонной активности.
Материалы
Отлов ужа водяного и лабораторные исследования велись на территории Северного (РФ, Астраханская область) и Восточного (Казахстан, Мангистауская область) Прикаспия (рис. 1). Все работы со змеями проводили в соответствии с «Международными рекомендациями по проведению медико-биологических исследований с использованием животных» (International Recommendations..., 2012). В исследованиях использовали только половозрелых змей. Объем материала и краткая характеристика мест отлова представлены в табл. 1.
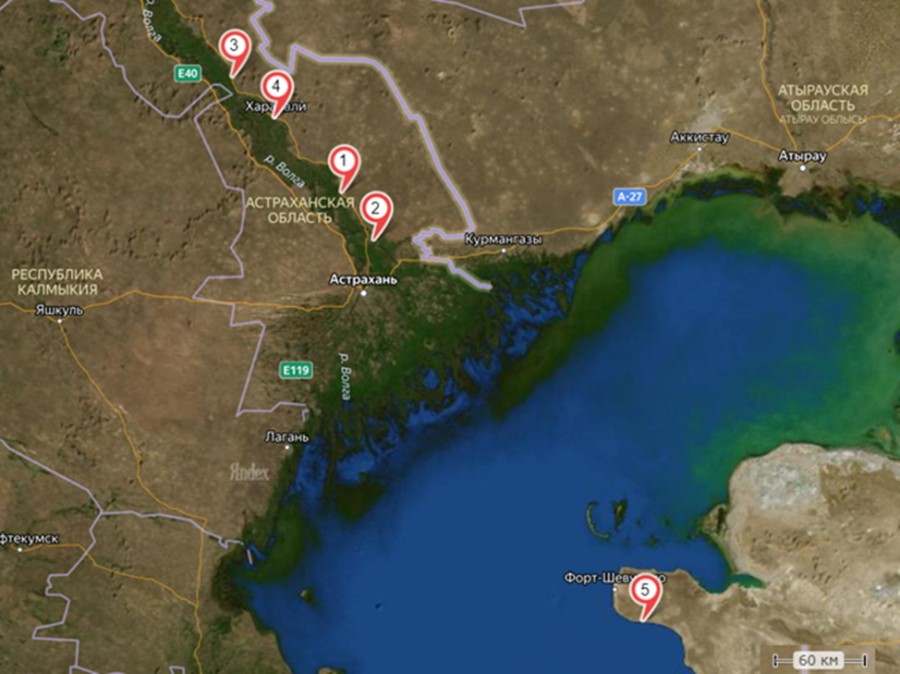
Рис. 1. Карта-схема мест отлова ужа водяного. Северный Прикаспий: 1 – Астраханская область, Красноярский район, пос. Комсомольский, с. Лапас; 2 – Астраханская область, Красноярский район, р. Ахтуба, пос. Белячий, с. Ясын-Сокан; 3 – Астраханская область, Харабалинский район, пос. Бугор, с. Михайловка; 4 – Астраханская область, Харабалинский район, г. Харабали. Восточный Прикаспий: 5 – Казахстан, Мангистауская область, окрестности с. Кызылозен
Fig. 1. Schematic map of Natrix tessellata trapping stations. Northern Pre-Caspian: 1 – village Lapas, settlement Komsomol'sky, Krasnoyarsky region, Astrahanskaya oblast'; 2 – v. Jasyn-Sokan, s. Belyachy, Krasnoyarsky region, Astrahanskaya oblast'; 3 – v. Mihaylovka, s. Bugor, Harabalinsky region, Astrahanskaya oblast'; 4 – Harabali city, Harabalinsky region, Astrahanskaya oblast'. Eastern Pre-Caspian: 5 – v. Kyzylozen, Mangistauskaya oblast', Kazahstan
Таблица 1. Краткая эколого-географическая характеристика мест отлова ужа водяного Natrix tessellata (Laurenti, 1768)
| Время отлова | Место отлова | Самки (экз.) | Самцы (экз.) |
| Районы Северного Прикаспия | |||
| 23.08.17– 24.08.17 | Астраханская область, Красноярский район, между пос. Комсомольский и с. Лапас (координаты: 46°55'42.2"N, 47°51'45.6"E) (место водопоя скота и рыбалки местных жителей) | 17 | 8 |
| 25.08.17 | Астраханская область, Красноярский район, р. Ахтуба, между пос. Белячий и с. Ясын-Сокан (координаты: 46°40'48.4"N, 48°04'12.9"E) (частое место рыбной ловли и проезда автотранспорта) | 10 | 6 |
| 26.08.17 | Астраханская область, Харабалинский район, между пос. Бугор и с. Михайловка (координаты: 47°36'18.5"N, 46°53'15.2"E) (запруженный участок, частое место отдыха местных жителей) | 3 | 1 |
| 26.08.17 | Астраханская область, Харабалинский район, южные окрестности г. Харабали (координаты: 47°22'50.2"N 47°17'08.4"E) (близкое расположение населенного пункта, частый проезд автотранспорта) | 2 | 2 |
| Всего | 33 | 17 | |
| Район Восточного Прикаспия | |||
| 01.05.19– 2.05.19 | Казахстан, Мангистауская область, окрестности с. Кызылозен (координаты 44.17.55 с. ш., 50.30.16 в. д.) (каменистый берег Каспийского моря, редко посещаемый местным населением) | 27 | 27 |
| Всего | 27 | 27 | |
| Итого | 104 экз. | ||
В местах отлова ужей на территории Астраханской области антропогенное влияние было более выражено по сравнению с территорией Казахстана.
Методы
Для получения образцов крови животных обездвиживали путем захвата и делали пункцию верхнечелюстной вены иглой, смоченной в растворе гепарина, для взятия крови и приготовления мазков. Все работы проводились в соответствии с «Международными рекомендациями по проведению медико-биологических исследований с использованием животных» (International Recommendations..., 2012). Для каждой особи готовили по два мазка крови. После фиксации в смеси спирт : эфир (1 : 1) мазки окрашивали по Романовскому – Гимзе в течение 20 мин. (Ромейс, 1954). Дифференцированный подсчет лейкоцитов проводили на бинокулярном микроскопе Meiji Techno серии МТ 4000 с иммерсией (×1500). Всего проанализировано 206 препаратов и просмотрено 2060 лейкоцитов. С учетом морфологических особенностей определяли шесть типов лейкоцитарных клеток (в %): гранулоциты (гетерофилы, базофилы, эозинофилы) и агранулоциты (азурофилы, моноциты, лимфоциты) (Campbell, 2006; Хайрутдинов, Соколина, 2010; Павлов, 2019). На основании лейкоцитарной формулы крови рассчитаны интегральные лейкоцитарные индексы (отн. ед.):
- Индекс сдвига лейкоцитов (ИСЛ), отн. ед.: = ∑ гранулоцитов / ∑ агранулоцитов;
- Индекс соотношения лимфоцитов и эозинофилов (ИСЛЭ), отн. ед.: = Л/Э;
- Индекс соотношения гетерофилов и эозинофилов (ИСГЭ), отн. ед.: = Г/Э ;
- Лимфоцитарно-гранулоцитарный индекс (ИЛГ), отн. ед.: = Л • 10/(Э + Г + Б);
- Индекс соотношения гетерофилов и лимфоцитов (ИСГЛ), отн. ед.: = Г/Л,
где Л – лимфоциты; Э – эозинофилы; Г – гетерофилы; Б – базофилы.
Статистическая обработка. Анализ полученных данных проводили методами непараметрической статистики с расчетом критериев: Краскела – Уоллиса (H) (при множественном сравнении независимых групп по одному признаку), Манна – Уитни (U) и Данна (D) (при попарном сравнении групп) в пакете прикладных программ Statistica. За величину статистической значимости принимали a = 0.05. При проведении множественных сравнений производилась коррекция критического уровня значимости. С учетом вида распределения центральные тенденции и рассеяние изученных показателей описывали медианой (Ме) и интерквартильным размахом (IQR) (значения 25-го и 75-го процентилей) (Реброва, 2006).
Результаты
Исследование качественного и количественного состава лейкоцитов крови рептилий связано с рядом трудностей, обусловленных как особенностями морфологии клеток, диффузностью кроветворения, так и внешними факторами (Павлов, 2019). Для поддержания гомеостаза организма в специфичных условиях среды обитания животные используют набор универсальных эффекторных клеток. У змей это зернистые лейкоциты (гранулоциты), характеризующиеся наличием крупного сегментированного ядра и присутствием в цитоплазме специфических гранул, представлены базофилами, эозинофилами и гетерофилами (Campbell, 2006). Мононуклеарные клетки (азурофилы, моноциты и лимфоциты), в отличие от агранулоцитов млекопитающих, имеют многочисленные мелкие гранулы в цитоплазме. Моноциты – крупные клетки с бобовидным ядром, смещенным от центра клетки, являются фагоцитами, обрабатывают антигены, высвобождают цитокины – гуморальные регуляторы межклеточных взаимодействий (Arican, Cicek, 2010). Азурофилы имеют меньшие размеры по сравнению с моноцитами, крупное округлое ядро, также характеризуются высокой фагоцитарной активностью (Salakij et al., 2002). Лимфоциты – клетки округлой или овальной формы с округлым ядром, обеспечивают реализацию специфических реакций иммунной системы организма (Davis et al., 2008).
Лимфоцитарно-гранулоцитарный состав лейкограмм периферической крови ужа водяного Северного и Восточного Прикаспия характеризовался преобладанием агранулоцитов (70–75 %), доля гранулоцитов составляла 25–30 % (табл. 2). Значимые различия в лейкограммах исследованных выборок ужа водяного выявлены по содержанию как гранулоцитов (гетерофилов), так и агранулоцитов (моноцитов и лимфоцитов).
Таблица 2. Лейкоцитарный состав периферической крови ужа водяного Natrix tessellata Северного и Восточного Прикаспия
| Статистичес-кие показатели | Гетерофилы, % | Базофилы, % | Эозинофилы, % | Азурофилы, % | Моноциты, % | Лимфоциты, % |
| Северный Прикаспий (n = 50) | ||||||
| Me | 5.00 | 9.50 | 9.00 | 8.00 | 6.50 | 62.25 |
| IQR | 2.00 | 4.00 | 4.00 | 2.00 | 2.00 | 7.00 |
| Восточный Прикаспий (n = 54 ) | ||||||
| Me | 12.00 | 9.50 | 8.25 | 8.00 | 11.25 | 51.75 |
| IQR | 4.25 | 5.25 | 5.25 | 4.25 | 5.50 | 8.50 |
| U | 6.68 | 1.43 | 0.93 | 1.28 | 6.23 | 5.65 |
| p – уровень значимости | < 0.01 | 0.15 | 0.35 | 0.20 | < 0.01 | < 0.01 |
Примечание. Жирным шрифтом выделены статистически значимые различия (a = 0.05).
Сравнительный анализ лейкограмм не выявил межполовых различий в морфологическом составе периферической крови ужа водяного Восточного Прикаспия. В лейкоцитарном профиле крови N. tessellata Северного Прикаспия межполовые различия наблюдались только в отношении азурофилов, доля которых в крови самок была выше по сравнению с самцами (D = 2.64, р = 0.05). Механизм таких различий между полами не ясен, но можно предположить, что в зависимости от сезонных изменений у полов наблюдаются разные колебания в количестве азурофилов.
Определение лейкоцитарной формулы является необходимой составной частью общего анализа крови. Однако отдельные ее показатели не дают целостного представления о реакциях системы крови и организма на воздействие различных факторов. Поэтому для интегральной оценки состояния системы крови используют лейкоцитарные интегральные индексы, отражающие взаимосвязи клеток крови и позволяющие оценить работу эффекторных механизмов иммунной системы.
Сравнительный анализ интегральных показателей выявил в популяции N. tessellata Восточного Прикаспия повышенное значение индексов ИСЛ, ИСГЭ, ИСГЛ и более низкое значение ИЛГ по сравнению с популяцией вида Северного Прикаспия (табл. 3).
Таблица 3. Значение интегральных индексов периферической крови ужа водяного Natrix tessellata
Северного и Восточного Прикаспия
| Статистические показатели | ИСЛ | ИСЛЭ | ИСГЭ | ИЛГ | ИСГЛ |
| Северный Прикаспий (n = 50) | |||||
| Me | 0.34 | 7.09 | 0.61 | 23.63 | 0.09 |
| IQR | 0.07 | 3.25 | 0.38 | 8.89 | 0.03 |
| Восточный Прикаспий (n = 54) | |||||
| Me | 0.40 | 6.57 | 1.43 | 18.08 | 0.25 |
| IQR | 0.16 | 4.38 | 1.04 | 8.12 | 0.11 |
| U | 3.25 | 0.58 | 6.11 | 4.37 | 6.78 |
| р – уровень значимости | < 0.01 | 0.56 | < 0.01 | < 0.01 | < 0.01 |
Примечание. Жирным шрифтом выделены статистически значимые различия (a = 0.05).
Полученные результаты свидетельствовали о перераспределении соотношения гранулоцитов (U = 2.95, p = 0.003) к агранулоцитам (U = 1.96, p = 0.04) в крови исследованных популяций. В иммунологических показателях крови особей Восточного Прикаспия преобладала неспецифическая защитная составляющая. Напротив, усредненная лейкограмма особей популяции ужей Северного Прикаспия имела ярко выраженный лимфоцитарный профиль, что свидетельствовало о возрастании специфической реактивности организма (рис. 2). Отметим, что активация специфических реакций иммунной системы (содержание лимфоцитов) прослеживалась в лейкоцитарном составе крови у самцов и самок популяции Северного Прикаспия.
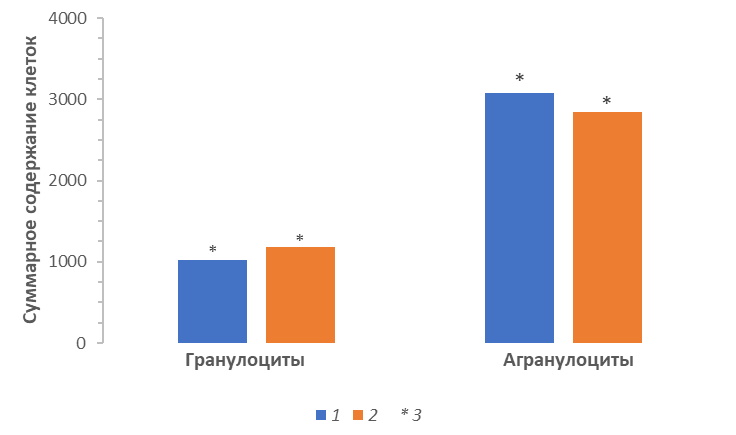
Рис. 2. Перераспределение соотношения гранулоцитов к агранулоцитам в составе периферической крови особей ужа водяного Прикаспийской низменности. 1 – Северный Прикаспий; 2 – Восточный Прикаспий; 3 – статистически значимые различия
Fig. 2. Redistribution of the ratio of granulocytes to agranulocytes in the periferal blood of N. tessellata of the Caspian Lowland. 1 – Northern Pre-Caspian; 2 – Eastern Pre-Caspian; 3 – statistically significant differences between groups
Особенностью морфологической картины и самок, и самцов ужа водяного Восточного Прикаспия являлось повышенное содержание в крови гетерофилов и моноцитов, которое статистически значимо превышало аналогичные показатели самок и самцов ужа водяного в популяции Северного Прикаспия (табл. 4).
Таблица 4. Лейкограммы крови самок и самцов ужа водяного Natrix tessellata
Прикаспийской низменности
| Статистичес-кие показатели | Гетерофилы, % | Базофилы, % | Эозинофилы, % | Азурофилы, % | Моноциты, % | Лимфоциты, % |
| Северный Прикаспий, самки (n = 33) | ||||||
| Me | 5.00 | 9.00 | 10.00 | 9.00 | 7.00 | 59.00 |
| IQR | 1.50 | 3.50 | 4.00 | 3.00 | 1.50 | 6.50 |
| Восточный Прикаспий, самки (n = 27) | ||||||
| Me | 12.50 | 10.00 | 8.00 | 8.00 | 11.00 | 51.50 |
| IQR | 3.50 | 8.50 | 4.00 | 4.50 | 5.00 | 10.00 |
| U | 5.16 | 0.13 | 2.15 | 0.49 | 3.99 | 3.44 |
| p – уровень значимости | <0.001 | 1.00 | 0.19 | 1.00 | 0.0004 | 0.003 |
| Северный Прикаспий, самцы (n = 17) | ||||||
| Me | 5.00 | 10.00 | 8.00 | 7.00 | 6.00 | 65.50 |
| IQR | 1.75 | 1.75 | 3.50 | 1.00 | 1.75 | 5.50 |
| Восточный Прикаспий, самцы (n = 27) | ||||||
| Me | 11.50 | 9.00 | 8.50 | 8.00 | 11.50 | 52.00 |
| IQR | 5.25 | 4.50 | 5.50 | 5.00 | 5.25 | 8.50 |
| U | 4.41 | 2.33 | 1.18 | 2.84 | 4.88 | 4.88 |
| p – уровень значимости | < 0.001 | 0.12 | 1.00 | 0.03 | < 0.001 | < 0.001 |
Примечание. Жирным шрифтом выделены статистически значимые различия (a = 0.05)
Количество гетерофилов в крови рептилий может зависеть от наличия заболеваний или повреждения тканей (Campbell, 2006). В мазках крови ужа водяного Восточного Прикаспия токсических гетерофилов (появление которых связано с бактериальными инфекциями) не наблюдалось. Повышенные доли гетерофилов и моноцитов в крови особей этой выборки ужа могли быть связаны со строго индивидуальными факторами, такими как восприимчивость отдельной змеи к стрессу, наличие микроорганизмов в окружающей среде или некоторые другие внутренние и внешние факторы.
Интегральные индексы (ИСГЭ, ИЛГ, ИСГЛ) выявили однотипный характер изменчивости параметров лейкоцитарной системы крови самок и самцов, обитающих в районах Прикаспийской низменности. Так, например, индексы соотношения гетерофилов к лимфоцитам (ИСГЛ) и гетерофилов к эозинофилам (ИСГЭ) были статистически значимо выше у самцов и у самок ужа водяного Восточного Прикаспия по сравнению с особями Северного Прикаспия (рис. 3).
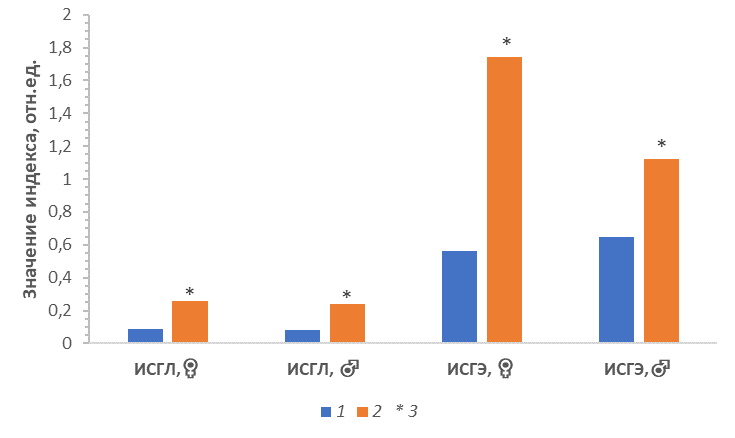
Рис. 3. Значение индекса отношения гетерофилов к лимфоцитам (ИСГЛ) и эозинофилам (ИСГЭ) для самок и самцов ужа водяного Северного и Восточного Прикаспия. 1 – Северный Прикаспий; 2 – Восточный Прикаспий; 3 – статистически значимые различия
Fig. 3. Values of the index of the ratio of heterophiles to lymphocytes in males and females of N. tessellata from the Northern and Eastern Pre-Caspian. 1 – Northern Pre-Caspian; 2 – Eastern Pre-Caspian; 3 – statistically significant differences
Обсуждение
Подводя итог проведенному исследованию, отметим, что лимфомиелоидный комплекс рептилий достаточно гетерогенен и фактически соответствует лимфоидной системе высокоорганизованных позвоночных животных. Рептилии подвергаются атаке самых разнообразных инфекционных и неинфекционных агентов, действию которых препятствуют две формы иммунного реагирования: неспецифический (врожденный) иммунитет и специфический (адаптивный, приобретенный) иммунитет. Врожденная система характеризуется более высоким развитием, быстро и эффективно реагирует на широкий спектр патогенных антигенов среды обитания по сравнению с адаптивными ответами. Иммунная система рептилий (наземных эктотермных животных) способна реагировать в широком диапазоне температур, но наиболее высокие ответы наблюдаются при определенной, оптимальной для конкретного вида температуре, поэтому исследование иммунных реакций требует учета временных и климатических факторов. Стратегия иммунной защиты определяется не только онтогенетическими особенностями рептилий, но и путем поступления, величиной и длительностью воздействия антигенов (Coico, 2003). Хорошо известно, что на проявление лейкоцитарного профиля крови и иммунный статус организма змей оказывают влияние возраст, пол, физиологическое состояние животных, а также сезоны года и изменение абиотических факторов среды (Павлов, Юсупов, 2015). Перечисленные факторы накладываются друг на друга и оказывают комбинированное действие на организм. Выявленные различия в параметрах лейкоцитарной системы крови между выборками из популяций ужа водяного в первую очередь связаны с сезонной динамикой физиологической активности животных. Уж водяной Северного Прикаспия был отловлен ближе к концу сезона физиологической активности, особи Восточного Прикаспия, напротив, в брачный период, связанный со спариванием и максимальной сезонной активностью. Большинство изменений в организме являются иммуноопосредованными или же напрямую детерминированы влиянием иммунологических факторов. Взаимоотношения организма со средой обитания являются результатом воздействия, с одной стороны, антигенов (ксенобиотиков), а с другой – распознавания и реагирования на эти антигены иммунной системы организма. Любые экологические факторы, изменяющие иммунологическую реактивность организма животных, могут нарушать формирование сложных механизмов обеспечения гомеостатического равновесия в системе «организм – среда обитания».
Заключение
В процессе адаптации к среде обитания и разных сроков онтогенетической активности у змей изменяются показатели клеточной и гуморальной защиты, что отражается на морфологическом составе крови. Вследствие этого особенно актуален поиск популяционных маркеров, позволяющих оценить работу эффекторных механизмов иммунной системы, а также уровень иммунологической реактивности, определяющие процесс формирования адаптивных реакций. Использованные в работе интегральные лейкоцитарные индексы обладали информативностью, иллюстрировали активацию специфической составляющей иммунитета особей ужа водяного Северного Прикаспия при подготовке к зимовке. Неспецифическая защитная система крови змей быстрее и эффективнее реагирует на широкий спектр патогенных антигенов среды обитания по сравнению с адаптивными ответами, поэтому в период спаривания именно врожденная система защиты, характеризующаяся более высоким развитием у эктотермных животных, обеспечивала максимальную защиту и устойчивое функционирование организма.
Библиография
Бакиев А. Г., Маленев А. Л., Зайцева О. В., Шуршина И. В. Змеи Самарской области . Тольятти: ООО «Кассандра», 2009. 170 с.
Васильев Д. Б. Ветеринарная герпетология . М.: Аквариум, 2016. 392 с.
Кленина А. А., Бакиев А. Г. Объем яиц в кладках обыкновенного ужа Natrix natrix и водяного ужа N. tessellata: работа над ошибками // Принципы экологии. 2015. Т. 4, № 4. C. 11–21.
Кленина А. А., Гордеев Д. А., Прилипко С. К. Питание ужей рода Natrix в Волгоградской области // Известия Самарского научного центра Российской академии наук. 2015. Т. 17, № 4 (4). С. 718–720.
Павлов А. В. Ключевые моменты гематологии рептилий: особенности оценки лейкоцитарной части крови // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2019. № 1 (25). С. 138–152. DOI: 10.21685/2307-9150-2019-1-14.
Павлов А. В., Юсупов Р. Х. Система крови // Гадюки (Reptilia: Serpentes: Viperidae: Vipera) Волжского бассейна. Ч. 1. Тольятти: Кассандра, 2015. С. 137–155.
Реброва О. Ю. Статистический анализ медицинских данных . М.: МедиаСфера, 2006. 312 с.
Романова Е. Б., Соломайкин Е. И., Бакиев А. Г., Горелов Р. А., Кленина А. А. Иммуногематологические показатели ядовитых и неядовитых змей на территориях Волжского бассейна с разной антропогенной трансформацией // Известия Самарского НЦ РАН. 2017. Т. 19, № 2. С. 54–61.
Романова Е. Б., Николаев В. Ю., Бакиев А. Г., Кленина А. А. Особенности лейкоцитарного состава крови самок обыкновенного ужа (Natrix natrix) и водяного ужа (N. tessellata) (Reptilia: Colubridae) Самарской области // Современная герпетология. 2015. Т. 15. Вып. 1/2. С. 69–76.
Ромейс Б. Микроскопическая техника / Пер. с нем. проф. В. Я. Александрова и З. И. Крюкова; Под ред. И. И. Соколова. М.: Изд-во иностр. лит., 1954. 719 с.
Табачишин В. Г., Табачишина И. Е. Распространение и особенности экологии обыкновенного ужа (Natrix natrix) на севере Нижнего Поволжья // Поволжский экологический журнал. 2002. № 2. С. 179–183.
Хайрутдинов И. З., Соколина Ф. М. Характеристика крови рептилий и ее связь с условиями среды обитания . Казань: Казанский университет, 2010. 44 с.
Чугуевская Н. М. Ужи (Serpentes, Colubridae, Natrix) Волжского бассейна: Экология и охрана : Автореф. дис. … канд. биол. наук. Тольятти, 2005. 20 с.
Arican H., Cicek K. Morphology of peripheral blood cells from various species of Turkish herpetofauna // Acta Herpetologica. 2010. Vol. 5, No 2. P. 179–198.
Bakiev A., Kirillov A., Mebert K. Diet and Parasitic Helminths of Dice Snakes from the Volga Basin, Russia // The Dice Snake, Natrix tessellata: Biology, Distribution and Conservation of a Palaearctic Species / Mertensiella. 2011. № 18. P. 325–329.
Campbell T. W. Clinical pathology of reptiles // Reptile medicine and surgery. 2nd edition. St. Louis (MO): Saunders Publishing, 2006. P. 453–470.
Cooper E. L., Klempau A. E., Zapata A. G. Reptilian Immunity // Biol. Reptilia. New York, 1985. Vol. 14. P. 601–636.
Coico R. Immunology. A Short Course. Hoboken; NJ: Wiley-Liss Publications, 2003. 237 p.
Davis A. K., Maney D. L., Maerz J. C. The use of leukocyte profiles to measure stress in vertebrates: a review for ecologists // Functional Ecology. 2008. Vol. 22. P. 760–767.
International Recommendations for Biomedical Research Using Animals. 2012. URL: https://grants.nih.gov/grants/olaw/guiding_principles_2012.pdf
Kobolkuti L., Cadar D., Czirjak G., Niculae M. et al. The Effects of Environment and Physiological Cyclicity on the Immune System of Viperinae // The Scientific World Journal. 2012. Vol. 4. DOI: 10.1100/2012/574867
Lisicic D., Dikic D., Benkovic V., Knezevic A. H. et al. Biochemical and hematological profiles of a wild population of the nose-horned viper Vipera ammodytes (serpentes: Viperidae) during autumn, with a morphological assessment of blood cells // Zoological studies. 2013. Vol. 52, № 1. P. 11–20.
Litvinov N., Bakiev A., Mebert K. Thermobiology and Microclimate of the Dice Snake at its Northern Range Limit in Russia // The Dice Snake, Natrix tessellata: Biology, Distribution and Conservation of a Palaearctic Species / Mertensiella. 2011. № 18. P. 330–335.
Salakij C., Salakij J., Chanhome L. Comparative hematology, morphology and ultrastructure of blood cells in Monocellate cobra (Naja kaouthia), Siamese spitting cobra (Naja siamensis) and Golden spitting cobra (Naja sumatrana) // Kasetsart J. (Nat. Sci.). 2002. Vol. 36. P. 291–300.
Благодарности
Авторы признательны к. б. н., доценту, старшему научному сотруднику Волжско-Камского государственного природного биосферного заповедника Алексею Владиленовичу Павлову за ценные советы при проведении дифференцированного анализа лейкоцитарного состава крови.

 © 2011 - 2026
© 2011 - 2026