



Введение
Зимняя спячка присутствует у представителей широкого филогенетического диапазона мелких и средних млекопитающих. Характеризуется длительными периодами гипотермического оцепенения, которые регулярно прерываются краткими интервалами нормотермии, пробуждениями (Melvin and Andrews, 2009). Пробуждения – по-прежнему загадка зимней спячки, но их значение таково, что они встречаются у подавляющего большинства зимоспящих млекопитающих. При пробуждениях фаза быстрого саморазогревания включает в себя несократительный термогенез, возникающий в специализированном «органе саморазогревания», уникальном для млекопитающих и называемом бурой жировой тканью (Калабухов, 1985; Ballinger and Andrews, 2018). Предполагается, что пробуждения жизненно необходимы мелким зимоспящим млекопитающим для возобновления транскрипции, обновления внутриклеточных структур и активации иммунной системы для борьбы с патогенами. Синтез макромолекул, таких как РНК и белок, более эффективен и проходит в более высоком темпе, при высоких температурах тела (Van Breukelen and Martin, 2001, 2002; Prendergast et al., 2002; D'Alessandro et al., 2017; Wiersma et al., 2018).
При периодических спонтанных пробуждениях процесс саморазогревания происходит в сжатые сроки и сопровождается быстрым ростом уровня метаболизма и температуры тела. Скорость роста температуры тела – показатель способности организма зимоспящего животного быстро продуцировать и выделять достаточное количество энергии для саморазогревания. В представленном сообщении в сравнительном плане проанализирована динамика температуры тела и скорость роста температуры при регулярных периодических пробуждениях у особей пяти зимоспящих видов семейств Sciuridae и Erinaceidae.
Материалы
В работе использованы материалы, полученные от животных, постоянно проживавших в Республиканском зоопарке «Орто-Дойду»: черношапочных сурков (Marmota camtschatica Pallas, 1811) и арктических (берингийских) сусликов (Spermophilus parryii Richardsony, 1827). Родина этих животных – отроги Верхоянского хребта в Якутии. Работа с бурундуками (Tamias sibiricus jacutensis Ognev, 1935) и длиннохвостыми сусликами (Spermophilus undulatus Pallas, 1778) проводилась со зверьками, отловленными в окрестностях г. Якутска в безморозный период. Проанализированы данные, полученные в 2010–2019 гг. Сравнительные материалы по белогрудым ежам (Erinaceus roumanicus Barrett-Hamilton, 1900) получены при обработке первичных материалов и совместном написании статей по зимней спячке ежей (Рутовская и др., 2019а, б). В зимний период в подвальном помещении ИБПК СО РАН, где зимовали животные, поддерживали температурный режим, близкий к естественному на горизонтах почв коренных местообитаний животных. Зимой 2018/19 г. спячка длиннохвостых сусликов проходила в штольне подземной лаборатории Института мерзлотоведения СО РАН (г. Якутск) (Ануфриев, Ядрихинский, 2019). Белогрудые ежи зимовали на экспериментальной базе ИПЭЭ РАН зимой 2017/18 г. в Подмосковье в заранее подготовленных искусственных норах. Характеристика экспериментальных животных представлена в табл. 1.
Методы
Представителям сем. Беличьи до начала спячки, в конце августа – сентябре, внутрибрюшинно имплантировали приборы длительной регистрации температуры тела (температурные накопители DS-1922L, подробное описание прибора и основные характеристики можно найти на сайтах: www.elin.ru, http://www.thermochron.ru/, http://www.ibdl.ru/).
Таблица 1. Основные параметры экспериментальных животных
| Вид | Число особей | Период исследований | Пол животных | Масса тела перед спячкой, г |
| T. sibiricus | 3 | 2010/11; 2019/20 | 2♂; 1♀ | 97–120 |
| S. undulatus | 3 | 2018/2019 | 3♂ | 860–960 |
| S. parryii | 3 | 2009/10; 2014/15 | 2♂; 1♀ | 840–1180 |
| M. camtschatica | 3 | 2011/2012 | 2♂; 1♀ | 3200–3800 |
| E. roumanicus | 3 | 2017/18 | 3♂ | 980–1200 |
Приборы были запрограммированы на измерение температуры с частотой 1 раз в 60 мин. Аналогичными приборами проводили регистрацию температуры окружающей среды. Бурундукам, из-за их относительно небольших размеров, приборы имплантировали под кожу, в переднюю часть тела (в подмышечную впадину). Белогрудым ежам до начала спячки в те же сроки внутрибрюшинно были имплантированы измерители Петровского (Петровский и др., 2008) (термонакопители ДТН4-28 «ЭМБИ РЕСЕРЧ», Новосибирск). Измерения проводились с частотой 1 раз в 20 мин.
Проанализировано по 6–7 пробуждений в период «глубокой» спячки (декабрь – март) у трех особей каждого вида. У бурундуков и ежей – в диапазоне температуры среды от -3...5 до +3...5о. У двух видов сусликов и черношапочных сурков в диапазоне от -10...8 до +2...4о. В этих температурных диапазонах проходит зимняя спячка исследованных видов в естественой среде. У всех видов расчет скорости температуры тела и времени разогревания проводили при достижении скорости роста температуры 0.01–0.02 о/мин. Окончанием саморазогревания считали снижение скорости роста температуры также 0.01–0.02 о/мин. Сравнивались максимальные и средние скорости роста температуры тела при пробуждении, время пробуждений у пяти зимоспящих видов. Статистическую обработку проводили, используя стандартный пакет анализа MS Excel, при сравнении средних значений измеряемых величин использовали критерий Стьюдента. Работу с животными проводили с учетом рекомендаций национального стандарта, по принципам надлежащей лабораторной практики Российской Федерации, ГОСТ З 53434-2009.
Результаты
За время зимней спячки представители изучаемых видов многократно погружались в состояние гипотермии, периодически прерываемое пробуждениями. Анализ полученных и имеющихся материалов показал, что число пробуждений за сезон зимней спячки у разных видов различно. Так, два вида сусликов (Sp. undulatus и Sp. parryii) за почти шестимесячный период спячки при оптимальных для спячки температурах среды в основном пробуждались от 12 до 15 раз, максимальная продолжительность гипотермии составляла 400 ч и более. Бурундук, обладающий наименьшей массой тела среди сравниваемых видов, за спячку пробуждался 20 раз и более с максимальной продолжительностью до 229 ч. Черношапочный сурок, наиболее крупный представитель зимоспящих, за спячку пробуждался около 20 раз, максимальная продолжительность гипотермии превышала 400 ч. Наибольшее число пробуждений за спячку отмечено у белогрудого ежа, до 35. Максимальная продолжительность гипотермии у белогрудых ежей также около 400 ч (рис. 1). У черношапочного сурка и белогрудого ежа календарная продолжительность спячки в естественной среде выше, чем у остальных видов (Ануфриев, 2008; Ануфриев, Ядрихинский, 2019; Рутовская и др., 2019а).

Рис. 1. Типичные примеры хода зимней спячки (по температуре тела) у бурундука (А), длиннохвостого (Б) и арктического (В) сусликов, черношапочного сурка (Г) и белогрудого ежа (Д) (Рутовская и др., 2019)
Fig. 1. Typical examples of the course of hibernation (body temperature) in the Chipmunk (A), Long-tailed (Б) and Arctic (B) ground squirrels, Black-capped marmot (Г), and White-breasted hedgehog (Д) (Rutkovskaya et al., 2019)
У всех экспериментальных животных продолжительность активной части саморазогреваний составляла в среднем 4–6 ч (табл. 2), а температура тела у окончательно пробудившихся зверьков имела значения 33–35 оС, в редких случаях до 36 оС. Анализ графика температурной зависимости показал, что повсеместно присутствует S-образность температурного графика саморазогревания (рис. 2).
Таблица 2. Скорость и время саморазогревания при пробуждении у пяти зимоспящих видов
| Вид / параметр | T. sibiricus (M ± m) Me Min – Max |
S. undulatus
(M ± m) Me Min – Max |
S. parryii
(M ± m) Me Min – Max |
M. camtschatica (M ± m) Me Min – Max |
E. roumanicus (M ± m) Me Min – Max |
| Vмах оС/мин | (0.33 ± 0.02)0.37 0.15–0.45 | (0.22 ± 0.017)0.23 0.13–0.37 | (0.2 ± 0.007)0.21 0.17–0.24 | (0.2 ± 0.01)0.16 0.133–0.28 |
(0.35 ± 0.02)0.34 0.26–0.45 |
| Vсредн. оС/мин | (0.15 ± 0.01)0.14 0.12–0.2 | (0.12 ± 0.01)0.11 0.09–0.154 | (0.106 ± 0.03)0.1 0.084–0.122 | (0.096 ± 0.006)0.09 0.055–0.12 | (0.12 ± 0.004)0.12 0.092–0.148 |
| Длит. пробужд. ч | (4.4 ± 0.27)4.0 3.0–6.0 |
(5.7 ± 0.18)5.6 5.0–7.0 |
(5.4 ± 0.16)5.2 5.0–6.0 | (5.9 ± 0.19)6.0 5.0–8.0 |
(4.8 ± 0.27)4.5 3.7–6.0 |
Примечание. m – количество проанализированных пробуждений, у всех видов, по 20; Ме – медиана.
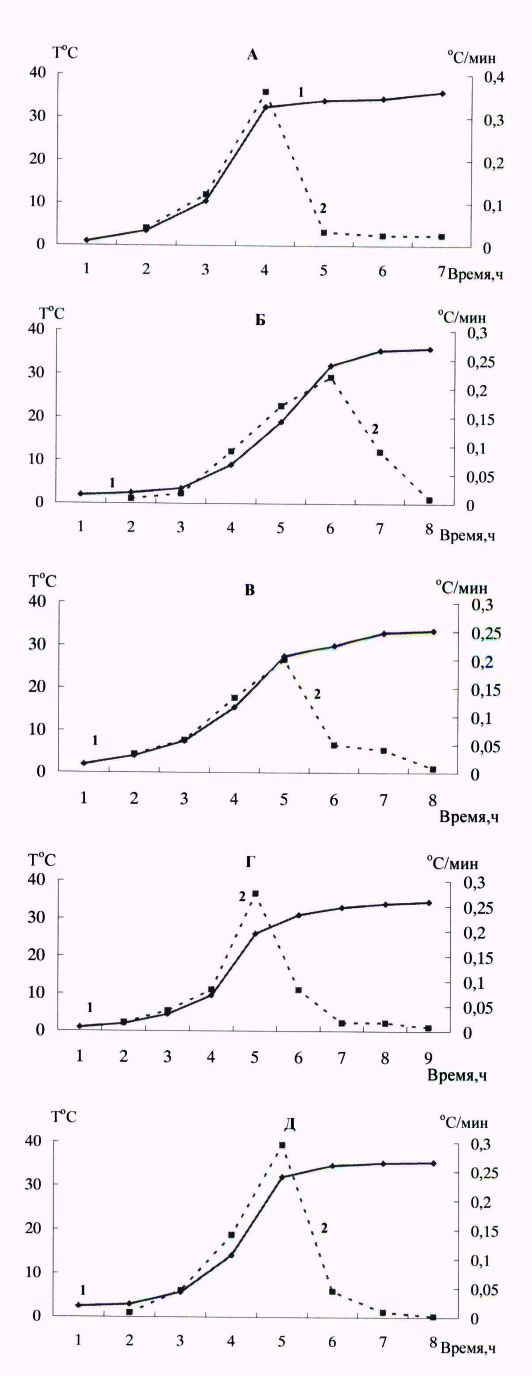
Рис. 2. Динамика температуры тела при пробуждении у сибирского бурундука (А), арктического суслика (Б), длиннохвостого суслика (В), черношапочного сурка (Г) и белогрудого ежа: 1 – температура тела; 2 – скорость роста температуры тела
Fig. 2. Dynamics of body temperature on arousal in the Siberian chipmunk (A), Arctic ground squirrel (B), Long-tailed souslik (В), Black-capped marmot (Г) and White-breasted hedgehog: 1 – body temperature; 2 – rate of body temperature growth
Это означает, что саморазогревание – процесс активный и имеет кумулятивную направленность. Скорость роста температуры тела у животных различна на разных этапах саморазогревания. На начальном этапе температура тела растет медленно, почти линейно. Затем наступает фаза быстрого роста, которая у всех животных происходит в диапазоне температуры тела от 10–12 до 22–25 оС. Скорость роста температуры достигает максимума, после чего резко снижается. Имеется точка «перелома» графика скорости роста температуры тела, указывающая на окончание активного процесса саморазогревания. В начальный период саморазогревания и при максимальной скорости продуцирования тепла происходит разогрев и аккумуляция тепла в передней части тела, которая разогревается значительно быстрее задней. Это фаза несократительного термогенеза. Более быстрый разогрев передней части тела обусловлен локализацией в ней запасов бурого жира, расположенного в подмышечных впадинах, а также покрывающего тонким слоем сердечную сумку и внутреннюю часть грудного отдела позвоночника. При саморазогревании длиннохвостого суслика разница температуры в ротовой полости и в прямой кишке превышает 10 оС (Ануфриев, 2008). После прохождения пика максимальной скорости роста температуры тела выработанное тепло перераспределяется по всему организму, скорость роста температуры тела снижается и становится нулевой. Эта часть саморазогревания сопровождается выраженным «дрожанием» животных – фаза сократительного термогенеза. У представителей всех пяти рассматриваемых видов пиковая скорость роста температуры тела составляла в среднем 0.2–0.35 град./мин. А средняя скорость роста температуры – 0.1–0.15 град./мин. Чем продолжительнее время саморазогревания, тем ниже скорость роста температуры тела. У белогрудого ежа и сибирского бурундука среднее время саморазогревания близко по значениям, максимальная скорость роста температуры также близка.
Температура окружающей среды непосредственно влияет на общее время пробуждения. Во время спячки при температурах среды ниже минус 5–6 оС температура тела у рассматриваемых животных, кроме сибирского бурундука, могла иметь значения ниже 0 оС (см. рис. 1) (Ануфриев, 2008; Рутовская, 2019б). В этом случае увеличивалась продолжительность начальной стадии саморазогревания, предшествующей «быстрому» разогреванию. Ее длительность зависела от температуры окружающей среды, начальной температуры тела и, вероятно, массы зверька. Так, у длиннохвостого суслика, пробуждение которого началось с температуры в полости тела минус 1 оС, начальная стадия пробуждения, когда температура тела росла медленно и почти линейно, длилась несколько часов. За это время она подросла на 3–4о. Затем при достижении температуры 2–3 оС начиналось обычное пробуждение с характерным быстрым ростом температуры, и за 6 ч зверек приобретал температуру около 34–35 оС (рис. 3). Стрелками показаны начало и окончание активной части пробуждения. На рис. 3 пробуждение взрослого самца 6–7 марта, это тот же зверек на рис. 1. У исследованных животных, спячка которых проходила с температурой тела 2–3 оС и выше, быстрый рост температуры начинался после непродолжительного предварительного этапа.

Рис. 3. Динамика роста температуры тела у длиннохвостого суслика при пробуждении в спячке с температурой тела ниже 0 оС: 1 – температура тела; 2 – температура среды. Стрелками обозначено начало и окончание процесса разогревания
Fig. 3. Dynamics of body temperature growth in a long-tailed ground squirrel upon interbout arosal with body temperature below 0 °C: 1 – body temperature; 2 – ambient temperature. Arrows indicate the beginning and end of the heating process
У исследованных видов на протяжении периода «глубокой» спячки при пробуждениях основные характеристики «активного» саморазогревания (время, скорость) изменялись незначительно (рис. 4).
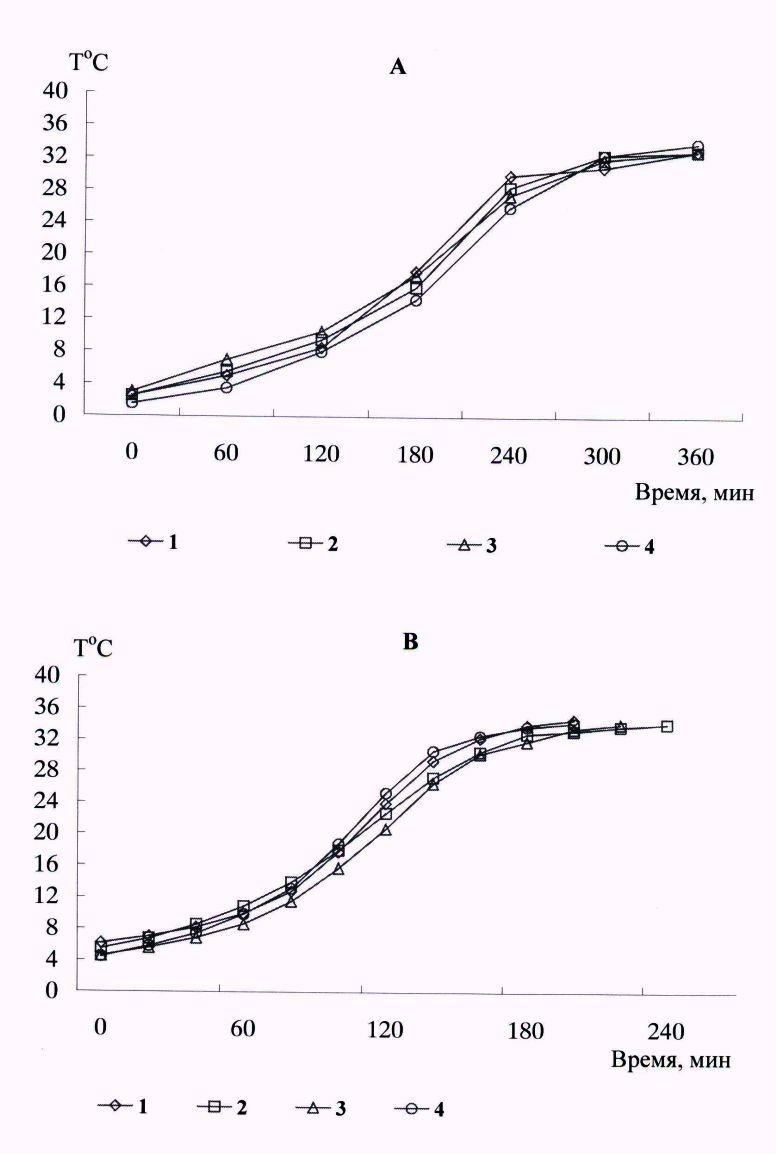
Рис. 4. Динамика температуры тела при пробуждении у длиннохвостого суслика (А) и белогрудого ежа (В) на протяжении спячки: 1 – декабрь; 2 – январь; 3 – февраль; 4 – март
Fig. 4. Dynamics of body temperature on awakening in a long-tailed ground squirrel (A) and white-breasted hedgehog (B) during hibernation: 1 – December; 2 – January; 3 – February; 4 – March
Обсуждение
С переходом зимоспящих животных к норной жизни и потери связи с поверхностью почвы количество воздействующих на них внешних факторов, синхронизирующих эндогенные ритмы с экзогенными, значительно снижается. Исчезает суточный периодизм изменения освещенности, температуры и влажности окружающей среды. Основными факторами, позволяющими синхронизировать эндогенные ритмы с сезонными изменениями внешней среды, по крайней мере для зимоспящих северо-востока Сибири, остаются лишь температура, в меньшей степени – атмосферное давление и влажность почвы (Ануфриев, 2008). Регуляция ритмов зимней спячки в значительной степени связана с температурой внешней среды. Так, для длиннохвостого суслика еще в начале 90-х годов прошлого века была выявлена зависимость продолжительности периодов гипотермии и уровня метаболизма от температуры среды. Позднее сходная зависимость была получена для арктического суслика с Аляски (Ануфриев, Ахременко, 1990; Buck, Barnes, 2000). Аналогичные зависимости выявлены для сибирского бурундука, черношапочного сурка и белогрудого ежа (Ахременко и др., 1998; Ануфриев, 2008; Рутовская и др., 2019). У каждого зимоспящего вида в период спячки имеется температурный диапазон, в котором продолжительности периодов гипотермии максимальны, а уровень обмена минимален. Этот диапазон температуры среды близок к температурам почвы, месторасположениям зимовочных гнезд коренных местообитаний видов, зонам «оптимума» спячки (Ануфриев, 2008). Частота пробуждений связана с температурными условиями протекания спячки и косвенно подтверждает гипотезу о метаболической природе пробуждений (Калабухов, 1985). Температура среды, при которой проходит спячка, влияет на продолжительность саморазогревания животных. Так, у длиннохвостого суслика в начальный период спячки при температуре среды 3–4 оС саморазогревание при пробуждении проходит за 4 ч, в период глубокой спячки около 6 ч. У белогрудого ежа в начальный период спячки при относительно высоких температурах среды также наблюдается высокая частота пробуждений, а время саморазогревания значительно короче. Анализ динамики температуры тела у длиннохвостого и золотистого сусликов показал, что чем выше начальная температура тела, тем меньше продолжительность пробуждений (Соломонов и др., 1987; Utz et al., 2007). Поэтому мы рассматриваем пробуждения после установления стабильного ритма спячки, который мы условно называем «глубокая» спячка. Ранее анализ пробуждений длиннохвостого суслика показал, что самопроизвольные (спонтанные) пробуждения по характеру графической зависимости не отличались от спровоцированных (вызванных) пробуждений. При пробуждениях животных с температурой тела ниже 5 оС скорость разогревания достигает максимума в диапазоне температуры тела 20–23 оС. Если пробуждение проходит при более высоких температурах, уменьшается время разогревания, максимальная скорость разогревания – при более высоких температурах тела (Ануфриев, 2008).
У изучаемых видов «активная» часть саморазогревания начинается при температурах тела около 2–3 оС, у ежей чуть выше. У этих видов, за исключением бурундука, отмечена спячка с температурой тела ниже 0 оС. При спячке с температурой тела ниже 0 оС перед началом «активного» саморазогревания температура тела гибернирующих животных возрастает практически линейно, продолжительность этой части пробуждения зависит от температуры тела, следовательно, и от температуры среды и может продолжиться несколько часов.
Периодические пробуждения мелких зимоспящих животных энергетически необычайно расточительны. Это отмечал еще Н. И. Калабухов (1985), они имеют метаболическую природу. Основным источником тепла в мышцах разогревающихся зимоспящих животных является свободное несопряженное окисление, в основе которого лежит разобщение окисления и фосфорилирования (Нейфах, Даудова, 1964; Скулачев, 1972). И действительно, наибольшая часть энергозатрат в период спячки связана именно с периодическими пробуждениями. Прямое измерение массы тела T. sibiricus и Sp. undulatus показало, что при каждом пробуждении потери массы тела у бурундуков составляют 0.15, а у сусликов 0.1 г/жив./ч. Наиболее энергозатратной частью является саморазогревание, потребление кислорода при максимальной скорости роста температуры тела у длиннохвостого суслика в три раза выше, чем у окончательно пробудившегося зверька. Графическая кривая потребления кислорода при пробуждении длиннохвостого суслика и график скорости роста температуры тела очень похожи (Ануфриев, 2008).
Саморазогревание у всех видов происходит в сжатые сроки с быстрым продуцированием тепла. У австралийской ехидны (Tachyglossus aculeatus) бурый жир отсутствует, температура тела при пробуждении растет практически линейно и продолжается свыше десяти часов (Augee, Ealeu, 1968). Среди рассматриваемых беличьих сибирский бурундук имеет размеры и массу тела на порядок меньшие по сравнению с остальными видами. Продолжительность пробуждения у бурундука, по сравнению с остальными видами беличьих, меньше только на 25–29 %, максимальная скорость разогревания выше на 60 %, а средняя скорость на 25–30 % (P < 0.05). У рукокрылых, например северного кожанка с массой тела около 10 г, при температурах среды от 0 до минус 2 оС саморазогревание при пробуждении занимает около 30 мин, а весь цикл – пробуждение и залегание в спячку – около 1.5 ч. Длительность оцепенений в естественной среде в период «глубокой» спячки, в январе – феврале, у кожанка составляет 8–11 сут и сопоставима с продолжительностью периодов гипотермии у бурундука (Ануфриев, Ревин, 2006; Кириллин и др., 2018).
У ежовых, имеющих массу тела близкую к двум видам сусликов, пробуждения проходят в более сжатые сроки и с более высоким продуцированием тепла в единицу времени. Причем время пробуждения на 15–20 % меньше, максимальная скорость на 60 %, а средняя выше на 15–18 % (P < 0.05). Вместе с тем графические зависимости динамики температуры тела и скорости роста температуры тела сходны со сравниваемыми видами (см. рис. 2). Имеется еще ряд сходных черт зимней спячки у представителей семейств Беличьи и Ежовые. Все исследованные виды объединяет способность находиться в состоянии зимней спячки при окружающих температурах среды ниже нуля. У трех видов беличьих описана способность находиться в состоянии гипотермии с температурой тела (внутрибрюшинно и под кожей) ниже 0 оС (Ануфриев, 2008). Совсем недавно было показано, что два вида ежей сем. Erinaceus, имеющих обширные ареалы на Европейском континенте (E. roumanicus и E. europaeus), могут находиться в спячке с температурой в полости тела ниже 0 оС. У всех рассматриваемых видов число пробуждений за спячку напрямую связано с температурой окружающей среды. Наиболее продолжительные периоды оцепенений отмечены при оптимальных для спячки каждого вида температурах среды, которые в коренных местообитаниях видов поддерживаются на большей части зимовки и спячки (Рутовская и др., 2019а; б; Ануфриев, 2020).
На протяжении сезона зимней спячки у особей исследованных видов в эксперименте поддерживались относительно постоянные температурные условия зимовки, сходные с естественными. Крайние значения температурного диапазона спячки поддерживались непродолжительное время. Зверьки находились в спячке в гнездах из пакли и сухой травы, уменьшая воздействие внешней температуры. На протяжении периода «глубокой» спячки, декабрь – март, не отмечено существенных изменений основных характеристик пробуждений (скорости роста температуры, увеличения или уменьшения времени саморазогревания) (см. рис. 4). Но в начале зимовок, когда температура среды всегда была относительно высокой, частота пробуждений была выше, температура тела зверьков составляла 6–7 оС, соответственно, время разогревания было менее продолжительным.
Заключение
Форма графических кривых динамики температуры тела, скорости роста температуры тела и, вероятно, механизм продуцирования тепла при пробуждениях сходны у рассмотренных видов. Имеются отличия в скорости роста температуры тела и продолжительности процесса разогревания, отражающие экологические особенности видов. Более крупные зимоспящие беличьи затрачивают на пробуждения больше времени, средняя и максимальная скорость роста температуры тела ниже. Белогрудый еж демонстрирует скорость роста температуры тела большую, а время разогрева меньшее по сравнению с близкими по размерам и массе тела беличьими. Возможно, это отличие является одной из особенностей энергетики зимней спячки ежовых.
Библиография
Ануфриев А. И. Механизмы зимней спячки мелких млекопитающих Якутии . Новосибирск: Изд-во СО РАН, 2008. С. 1–157.
Ануфриев А. И. Температурная регуляция ритмов зимней спячки // Природные ресурсы Арктики и Субарктики. 2020. Т. 25. № 1. С. 60–67.
Ануфриев А. И., Ахременко А. К. Энергетическая стоимость зимней спячки длиннохвостого суслика // Экология. 1990. № 5. С. 68–72.
Ануфриев А. И., Ревин Ю. В. Биоэнергетика зимней спячки летучих мышей (Chiroptera, Vespertilionidae) в Якутии. // Plecotus et al. 2006. № 9. С. 8–18.
Ануфриев А. И., Ядрихинский В. Ф. Температурная регуляция процессов зимней спячки у длиннохвостого суслика Spermophilus undulatus Pallas, 1778 // Принципы экологии. 2019. Т. 8. № 3. С. 8–17.
Ахременко А. К., Ануфриев А. И., Соломонов Н. Г., Соломонова Т. Н., Васильев В. Н. Зимняя спячка при температуре ниже нуля // Сибирский экологический журнал. 1998. № 3–4. С. 347–352.
Калабухов Н. И. Спячка млекопитающих . М.: Наука, 1985. 264 с.
Кириллин Р. А., Соломонов Н. Г., Ануфриев А. И., Охлопков И. М. Зимовка северного кожанка (Eptesicus nilssonii, Chiroptera, Vespertilionidae) в окрестностях г. Якутска (Центральная Якутия) // Зоологический журнал. 2018. Т. 97. № 9. С. 1171–1175.
Нейфах С. А., Даудова Г. М. Разобщение окислительного фосфорилирования в печени в момент пробуждения зимоспящего животного // Биохимия. 1964. Т. 29. № 5. С. 1003–1008.
Петровский Д. В., Новиков Е. А., Мошкин М. П. Динамика температуры тела обыкновенной слепушонки (ELLOBIUS TALPINUS, RODENTIA, CRICETIDAE) в зимний период // Зоологический журнал. 2008. Т. 87. № 12. С. 1504–1508.
Рутовская М. В., Диатроптов М. Е., Кузнецова Е. В., Ануфриев А. И., Феоктистова Н. Ю., Суров А. В. Динамика температуры тела белогрудого ежа (Erinaceus roumanicus Barrett-Hamilton, 1900) во время зимней спячки // Зоологический журнал. 2019а. № 5. С. 556–566.
Рутовская М. В., Диатроптов М. Е., Кузнецова Е. В., Ануфриев А. И., Феоктистова Н. Ю., Суров А. В. Феномен снижения температуры тела до отрицательных значений у ежей рода Erinaceus во время зимней спячки // Журнал эволюционной биохимии и физиологии. 2019б. Т. 55. Вып. 6. С. 463–464.
Скулачев В. П. Трансформация энергии в биологических мембранах . М: Наука, 1972. 448 с.
Соломонов Н. Г., Ахременко А. К., Ануфриев А. И. Динамика энергетических субстратов в тканях пробуждающихся сусликов // Механизмы зимней спячки. Пущино, 1987. С. 48–56.
Augee M. L., Ealeu E. N. Torpor of the echidna, Tachyglossus aculeatus // J. Mammol. 1968. Vol. 49. № 3. P. 446–464.
Ballinger M. A., Andrews M. T. Nature’s fat-burning machine: brown adipose tissue in a hibernating mammal // J. Exp. Biol. 2018. Vol. 221. DOI: 10.1242/jeb.162586.
Buck С. L., Barnes B. М. Effects of ambient temperature on metabolic rate, respiratory quotient, and torpor in an arctic hibernator // Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000. Vol. 279. № 1. P. 255–262.
D’Alessandro A., Nemkov T., Bogren L. K., Martin S. L. and Hansen K. C. Comfortably numb and back: plasma metabolomics reveals biochemical adaptations in the hibernating 13-lined ground squirrel // J. Proteome Res. 2017. Vol. 16. P. 958–969.
Melvin R. G. and Andrews M. T. Torpor induction in mammals: recent discoveries fueling new ideas // Trends Endocrinol. Metab. 2009. № 20. Р. 490–498.
Prendergast B. J., Freeman D. A., Zucker I. and Nelson R. J. Periodic arousal from hibernation is necessary for initiation of immune responses in ground squirrels // Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002. Vol. 282. P. 1054–1062.
Utz J. C., Velickovska V., Shmereva A., Van Breukelen F. Temporal and temperature effects on the maximum rate of rewarming from hibernation // Journal of Thermal Biology. 2007. Vol. 32. P. 276–281.
Van Breukelen F., Martin S. L. Translational initiation is uncoupled from elongation at 18 degrees C during mammalian hibernation // Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001. Vol. 281. P. 1374–1379.
Van Breukelen F., Martin S. L. Reversible depression of transcription during hibernation // J. Comp. Physiol. 2002. Vol. 172. P. 355–361.
Wiersma M., Beuren T. M. A., de Vrij E. L., Reitsema V. A., Bruintjes J. J., Bouma H. R., Brundel B. J., Henning R. H. Torpor-arousal cycles in Syrian hamster heart are associated with transient activation of the protein quality control system // Comp. Biochem. Physiol. (B). Biochem. Mol. Biol. 2018. Vol. 223. P. 23–28.
Благодарности
Работа выполнена в рамках темы Государственного задания ИБПК ЯНЦ СОРАН: «Структура и динамика популяций и сообществ животных холодного региона Северо-Востока России в современных условиях глобального изменения климата и антропогенной трансформации северных экосистем: факторы, механизмы, адаптации, сохранение» АААА-А17-117020110058-4.

 © 2011 - 2026
© 2011 - 2026