



Введение
Одной из основных целей интродукции растений в районы Крайнего Севера является улучшение качества городской среды. В последние десятилетия Российская Арктика все больше подвергается антропогенному воздействию вследствие активного развития промышленности, транспорта и логистики, разведки углеводородов (Abakumov et al., 2017). В связи с этим в северных городах необходимо формирование устойчивых зеленых насаждений с использованием видов, способных адаптироваться к экстремальным климатическим условиям и антропогенному загрязнению среды. Ряд интродуцентов, в частности из рода Syringa L., устойчивы к загазованности и задымлению воздуха (Кищенко, Потапова, 2014), они аккумулируют большое количество пыли и газов без потери декоративности (Полякова, 2011). Одним из таких видов является Syringa josikaea (сирень венгерская), она хорошо переносит городские условия, высокоустойчива к атмосферному загрязнению, обладает способностью к биоаккумуляции тяжелых металлов, проявляет к ним устойчивость, что свидетельствует о ее высоком адаптационном потенциале (Шлапак и др., 2017). S. josikaеа занимает одно из первых мест в ассортименте кустарников для озеленения Мурманской области, в то время как S. vulgaris (сирень обыкновенная) в условиях полярного дня не образует репродуктивных органов и не цветет. С целью интродукции саженцы сирени венгерской были впервые завезены в Полярно-альпийский ботанический сад в 1936 г. из Ботанического сада г. Ленинграда (Гонтарь и др., 2011) и уже в 1940 г. S. josikaеа стала использоваться для озеленения Мурманска и Мончегорска. Большая партия сирени была высажена в Мурманске в 1947 г. В настоящее время S. josikaea составляет основу зеленых насаждений г. Мурманска. Доля участия сирени в посадках на улицах и в скверах города в 1977 г. составляла 21 %, в 2012 г. – 28 % (Гонтарь и др., 2013).
Интегральным показателем, характеризующим адаптацию растений к условиям среды, является сезонный ритм развития (Логинов, 1980). В последние несколько десятилетий весенние фенофазы древесных растений, такие как появление листьев и цветение, стали более ранними по всему земному шару (Chambers et al., 2013; Wang et al., 2015), хотя изменения осенних фенофаз менее выражены (Gill et al., 2015). Фенофаза растений является важным биологическим индикатором климата и его воздействия на наземные экосистемы. Смещение фенофаз может привести к изменению продуктивности, репродукции, несоответствию между растениями и опылителями (DeLucia et al., 2012). Несмотря на большой ассортимент дендроинтродуцентов в городах Евро-Арктической зоны России, исследования особенностей их роста и развития в условиях океанического климата немногочисленны (Бабич и др., 2008; Гонтарь и др., 2013; Василевская, Морозова, 2016; Шлапак и др., 2017; Морозова, 2018; Vasilevskaya, Morozova, 2018; Vasilevskaya, Domachina, 2018).
Материалы
Объект исследования – Syringa josikaea Jacq. fil. (сирень венгерская) относится к секции сирени волосистые (Villosae C. K. Schneid), семейству Маслинные (Oleaceae Link.). S. josikaea – эндемичный кустарник Западных Карпат, имеющий узкий ареал и находящийся под угрозой исчезновения (Lendvay et al., 2013). В настоящее время является очень редким видом, произрастает в естественных условиях только в Украинских Карпатах и Западных Румынских Карпатах (Dihoru, Negrean, 2009). По последним данным (Lendvay et al., 2013), несколько локальных ценопопуляций существуют на небольших участках в долинах вдоль ручьев, часто представленные всего лишь несколькими особями и разделенные между собой большими расстояниями. S. josikaea – декоративный, быстрорастущий, высоко засухоустойчивый и морозостойкий вид, его экологическая пластичность определила успешную интродукцию в северных районах европейской части России (Гонтарь и др., 2010). В условиях интродукции является самым зимостойким видом по сравнению с другими представителями рода Syringa L. (Семенюк, Долганова, 2008; Мартынов, 2013).
Исследования проведены в г. Мурманске – самом большом в мире незамерзающем порту за полярным кругом. Город расположен в северной части Кольского полуострова, на берегу Кольского залива Баренцева моря (68°58' с. ш., 33°4' в. д.), на узкой, всхолмленной равнине, вытянутой с севера на юг на 25 км (рисунок). В Мурманске выделяют 3 внутригородские территории: Ленинский, Октябрьский и Первомайский районы, изолированные друг от друга крупными сопками. Наиболее характерными являются иллювиально-гумусовые подзолистые почвы (Переверзев, 2011). Город расположен в подзонах субарктической тундры (южный вариант) и лесотундры (Walker et al., 2018). Находится в атлантико-арктической зоне умеренного климата. На климат города значительное влияние оказывает теплое Северо-Атлантическое течение Баренцева моря. Среднегодовая температура воздуха +1.1 °С, средняя температура зимы –7.2 °С, лета +11.1 °С (Демин, 2012). Полярная ночь в Мурманске начинается 2 декабря и продолжается до 11 января, полярный день – с 22 мая по 22 июля (Богданец и др., 1998). Продолжительность вегетационного периода 120–130 дней. Ветер имеет муссонный характер. Относительная влажность воздуха колеблется в пределах 61–80 %. Годовое количество осадков составляет в среднем 495 мм. Снег в Мурманске лежит в среднем 210 дней.
2014 и 2015 гг., в которые проводились исследования, в основном соответствовали средней погодной норме региона. Есть различия по средней температуре в период, предшествующий началу вегетации. По многолетним данным, средняя температура мая составляет 4.0 °С, в 2014 г. ее значения соответствовали норме (4.2 °С), в 2015 г. значительно выше (6.9 °С). Средняя температура вегетационного сезона – 11.6 °С в 2014 г. и 10.6 °С в 2015 г., что близко к среднестатистической (11.0 °С). Различия отмечены по температурам июля. По многолетним данным, среднемесячная температура июля составляет 12.8 °С, в июле 2014 г. она была выше (13.6 °С), в 2015 г. – значительно ниже нормы (9.9 °С).
Основными источниками загрязнения г. Мурманска являются Мурманская ТЭЦ, Мурманский морской торговый порт, завод по термической обработке твердых бытовых отходов, 35-й судоремонтный завод АО ЦС «Звездочка» и база РТП «Атомфлот». Экологическая ситуация в городе в последние годы стала резко ухудшаться за счет повышения объемов перевалки угля открытым способом и его дробления Мурманским морским торговым портом, использования мазута низкого качества местными ТЭЦ. В 2015 г. в Мурманске отмечен высокий индекс загрязнения атмосферы (ИЗА = 2.84) (Доклад..., 2015).
Методы
Исследования проводились в г. Мурманске летом 2014–2015 гг. Пробные площади (см. рисунок) заложены с севера на юг и отличаются высотой над уровнем моря, климатическими условиями: ПП1 – сквер возле торгового центра «Мир»; ПП2 – сквер на ул. Ленинградской; ПП3 – посадки сирени около автобусной остановки «Автопарк»; ПП4 – посадки сирени около автобусной остановки «Ул. Шевченко». В качестве контрольной площадки (КП) выбран сквер около Музея Военно-воздушных сил Северного флота в поселке Сафоново (19 км на северо-восток от г. Мурманска). Сквер расположен на берегу Кольского залива Баренцева моря, вдали от автомобильных дорог и промышленных производств.
Скверы Мурманска, в которых проводились исследования, созданы в 40–80-е гг. ХХ в. Сквер на ул. Ленинградской (ПП2) является самым старым ботаническим объектом города, созданным в довоенные годы. Здесь еще в 1940 г. были высажены первые саженцы сирени, в настоящее время доля посадок S. josikaeа составляет 38 % от всех интродуцированных кустарников (Гонтарь и др., 2016). Посадки в сквере у торгового центра «Мир» (ПП1) датируются 1968 г. (Бродер, 1975). Здание Администрации Первомайского округа построено в 1979 г., в этот же год был заложен сквер (ПП4 – остановка «Ул. Шевченко»). В поселке Сафоново (контрольная площадка) посадки в сквере сделаны после создания музея Военно-воздушных сил Северного флота в 1976 г. На всех пробных площадках S. josikaeа является одним из доминирующих видов. Она представлена в основном старовозрастными (41–60 лет и более) и средневозрастными (до 40 лет) особями. На всех пробных площадках преобладают растения с раскидистыми кронами высотой 3–4 м, многие из них ослаблены в связи с отсутствием ухода и возрастом.
В работе использована методика фенологических наблюдений М. С. Александровой (1978), Н. Е. Булыгина и В. Т. Ярмишко (2001). Наблюдения велись за 10 экземплярами S. josikaеа на каждой пробной площадке с частотой 1–2 раза в неделю (с мая по октябрь) в течение двух лет. Отмечались следующие фенологические фазы: ПЧ – развертывание почек; ПБ1 – рост побегов (начало измерений); ПБ2 – окончание роста побегов; О1 – частичное одревеснение побегов; О2 – полное одревеснение побегов; Ц1 – начало цветения; Ц2 – окончание цветения; ПЛ – завязывание плодов; Л1 – расцвечивание листьев; Л2 – начало опадения листьев.
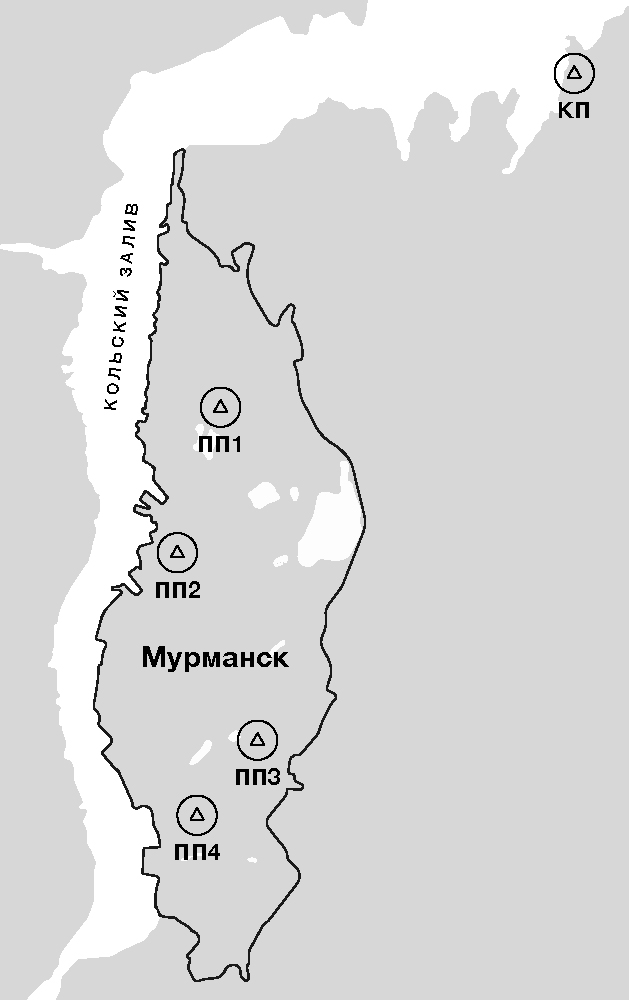
Схема расположения пробных площадок в г. Мурманске. КП – контрольная площадка в п. Сафоново; ПП1 – сквер возле торгового центра «Мир»; ПП2 – сквер на ул. Ленинградской; ПП3 – сквер около автобусной остановки «Автопарк»; ПП4 – сквер около автобусной остановки «Ул. Шевченко»
Lay out of sample plots in the city of Murmansk. КП – control plot in Safonovo; ПП1 – square near the shopping center "Mir"; ПП2 – square on the Leningradkaya Street; ПП3 – square near the bus stop "Autopark"; ПП4 – square near the bus stop "Shevchenko Street"
В течение двух вегетационных сезонов 2014–2015 гг. изучалась динамика роста побегов сирени. Еженедельно измерялись приросты 5 побегов на 10 маркированных растениях (N = 50) на каждой пробной площадке. Для исследования сезонного ритма развития S. josikaea рассчитаны суммы активных температур (САТ), при этом суммировались среднесуточные температуры выше 0 °С. В г. Мурманске только одна метеостанция, поэтому данных по климатическим различиям районов города нет. Показания температур получены по данным ГМС Мурманск ВМО 22113 (широта 68.95°, долгота 33.05°, высота над уровнем моря 51 м).
Результаты
В результате двух лет фенологических исследований Syringa josikaea получено, что вегетация сирени начинается в Мурманске 24 мая – 2 июня (табл. 1). Развертывание почек в 2014 г. происходило при сумме положительных температур 81.8–147.7 °С, в 2015 г. при 186.7–236 °С. Синхронность пробуждения объясняется очень коротким периодом покоя у сиреней (Пенкина, Ткаченко, 1974).
Таблица 1. Даты наступления фенофаз Syringa josikaea в г. Мурманске в 2014 и 2015 гг.
| № ПП | Год | ПЧ | ПБ1 | ПБ2 | О1 | О2 | Ц1 | Ц2 | ПЛ | Л1 | Л2 |
| КП | 2014 | 23.05 | 03.06 | 5.07 | 28.06 | 05.07 | 05.07 | 20.07 | 17.07 | 21.09 | 5.10 |
| 2015 | 01.06 | 03.06 | 5.07 | 28.06 | 11.07 | 11.07 | 18.07 | 17.07 | 25.09 | 8.10 | |
| ПП1 | 2014 | 24.05 | 03.06 | 6.07 | 28.06 | 05.07 | 06.07 | 21.07 | 17.07 | 20.09 | 6.10 |
| 2015 | 01.06 | 03.06 | 6.07 | 28.06 | 11.07 | 11.07 | 18.07 | 17.07 | 25.09 | 8.10 | |
| ПП2 | 2014 | 01.06 | 03.06 | 12.07 | 29.06 | 05.07 | 08.07 | 20.07 | 16.07 | 14.09 | 3.10 |
| 2015 | 29.05 | 03.06 | 7.07 | 27.06 | 12.07 | 10.07 | 20.07 | 16.07 | 25.09 | 4.10 | |
| ПП3 | 2014 | 02.06 | 03.06 | 12.07 | 29.06 | 06.07 | 10.07 | 22.07 | 19.07 | 20.09 | 4.10 |
| 2015 | 29.05 | 03.06 | 7.07 | 27.06 | 12.07 | 10.07 | 21.07 | 19.07 | 27.09 | 4.10 | |
| ПП4 | 2014 | 02.06 | 03.06 | 12.07 | 29.06 | 06.07 | 10.07 | 22.07 | 19.07 | 14.09 | 4.10 |
| 2015 | 29.05 | 03.06 | 7.07 | 27.06 | 12.07 | 10.07 | 21.07 | 19.07 | 27.09 | 4.10 |
Рост вегетативных побегов S. josikaea отмечен через несколько суток после развертывания почек. В оба года мониторинга наблюдалось синхронное начало линейного роста побегов во всех ценопопуляциях г. Мурманска и в пос. Сафоново – 3 июня (табл. 1, 2), в 2014 г. при сумме положительных температур 163 °С, в 2015 г. – 248 °С.
Таблица 2. Динамика прироста побегов Syringa josikaea в г. Мурманске (2014–2015 гг.) (в см)
| Даты | КП | ПП1 | ПП2 | ПП3 | ПП4 |
| 03.06.2014 03.06.2015 | 2.4 ± 0.1 2.4 ± 0.1 |
2.6 ± 0.1 2.6 ± 0.1 |
2.3 ± 0.1 2.2 ± 0.1 |
3.6 ± 0.1 3.6 ± 0.1 |
2.9 ± 0.1 2.9 ± 0.1 |
| 08.06.2014 8.06.2015 | 5.5 ± 0.2 3.8 ± 0.1 |
5.9 ± 0.2 6.9 ± 0.2 |
5.1 ± 0.3 4.9 ± 0.3 |
7.2 ± 0.2 7.2 ± 0.2 |
5.4 ± 0.2 5.4 ± 0.2 |
| 14–15.06.2014 16.06.2015 | 7.5 ± 0.3 7.5 ± 0.4 |
9.8 ± 0.3 9.8 ± 0.3 |
7.1 ± 0.3 6.9 ± 0.3 |
10.9 ± 0.3 11.8 ± 0.5 | 8.5 ± 0.3 10.5 ± 0.4 |
| 21–22.06.2014 21.06.2015 | 7.5 ± 0.3 12.0 ± 0.6 | 12.2 ± 0.4 12.5 ± 0.4 |
8.3 ± 0.3 8.2 ± 0.3 |
12.6 ± 0.4 13.3 ± 0.6 | 9.8 ± 0.3 12.2 ± 0.4 |
| 28–29.06.2014 28.06.2015 | 10.3 ± 0.6 14.1 ± 0.7 | 14.8 ± 0.5 15.1 ± 0.5 |
10.2 ± 0.3 9.9 ± 0.3 |
13.1 ± 0.4 14.5 ± 0.6 | 11.4 ± 0.5 13.0 ± 0.4 |
| 05–06.07.2014 06.07.2015 | 13.0 ± 0.8 15.5 ± 0.7 | 16.0 ± 0.5 15.7 ± 0.6 |
14.5 ± 0.3 14.1 ± 0.7 | 16.6 ± 0.5 16.7 ± 0.7 |
13.5 ± 0.4 15.7 ± 0.6 |
Окончание роста побегов S. josikaea происходит с 5 по 12 июля (см. табл. 1, 2). В первый год наблюдений разница в окончании фенофазы составляла между площадками около недели, во второй год мониторинга – 1–2 дня. При этом сумма активных температур в период окончания фенофазы значительно различается по годам. Если в 2014 г. она составила 429–445 °C, то в 2015 г. – 553–567 °С. Продолжительность фазы роста вегетативных побегов – 33–35 дней, только в 2014 г. в Октябрьском (ПП2) и Первомайском районах г. Мурманска (ПП3, ПП4) этот показатель был выше – 40 дней (табл. 3). Продолжительность роста побегов характеризует морозоустойчивость вида. Чем раньше заканчивается эта фенофаза, тем больше времени для запасания питательных веществ и подготовки к зимнему периоду покоя (Пенкина, Ткаченко, 1974).
Длина годичных приростов S. josikaea в конце вегетации варьирует по площадкам города в диапазоне 13.5–16.6 см в 2014 г., 14.1–16.7 см в 2015 г. Наибольшие годичные приросты в оба года мониторинга (см. табл. 2) отмечены в скверах у ТЦ «Мир» (ПП1 – Ленинский район) и на ост. «Автопарк» (ПП3 – Первомайский район). В Октябрьском районе, расположенном в центре города (сквер на ул. Ленинградской – ПП2) выявлено ингибирование роста побегов по сравнению с другими ценопопуляциями S. josikaea и контролем. Экспериментальная площадка расположена в районе с высоким уровнем техногенной нагрузки: рядом с автомагистралью и железной дорогой, а также Мурманским морским торговым портом, где происходит перевалка и дробление угля открытым способом.
Таблица 3. Продолжительность фенологических фаз Syringa josikaea в г. Мурманске (2014–2015 гг.)
| № ПП | Год | ПЧ | ПБ | О | Ц | Л | Продолж. вегетации |
| КП | 2014 | 12 | 33 | 8 | 16 | 15 | 136 |
| 2015 | 3 | 33 | 15 | 8 | 14 | 130 | |
| ПП1 | 2014 | 11 | 34 | 8 | 16 | 17 | 136 |
| 2015 | 3 | 34 | 15 | 8 | 14 | 130 | |
| ПП2 | 2014 | 3 | 40 | 7 | 13 | 20 | 125 |
| 2015 | 6 | 35 | 16 | 11 | 10 | 128 | |
| ПП3 | 2014 | 2 | 40 | 8 | 13 | 15 | 125 |
| 2015 | 6 | 35 | 16 | 12 | 8 | 129 | |
| ПП4 | 2014 | 2 | 40 | 8 | 13 | 21 | 125 |
| 2015 | 6 | 35 | 16 | 12 | 8 | 129 |
Частичное одревеснение побегов сирени отмечено 27–29 июня в оба года наблюдений, полное одревеснение 5–12 июля, продолжительность фенофазы составляет от 7 до 8 суток в 2014 г. и 15–16 в 2015 г. Одревеснение побегов S. josikaea и окончание их роста приходится в г. Мурманске на фазу начала цветения (см. табл. 1, 3), это значительно раньше, чем в других регионах России, поскольку у многих видов сирени окончание роста побегов практически совпадает с фазой окончания цветения (Полякова, 2011). Эта особенность развития S. josikaea в условиях арктического климата, по-видимому, связана с тем, что побеги, рано окончившие рост, успевают достаточно одревеснеть до наступления пониженных температур.
Критерием успешной интродукции является цветение и плодоношение. Фенофаза цветения в исследованных ценопопуляциях сирени венгерской начинается в Мурманске 6–11 июля. В других северных городах России, но на более низких широтах, цветение данного вида отмечается во второй половине мая – середине июня. На родине в Карпатах S. josikaea зацветает в конце мая – начале июня (Пенкина, Ткаченко, 1974). В 2014 г. цветание S. josikaea в Мурманске отмечено 5–10 июля при сумме активных температур 429.1–500.2 °С. Летом 2015 г. цветение началось практически синхронно на всех экспериментальных площадках 10–11 июля, при этом САТ составила 595–602 °С, его окончание отмечено 18–22 июля. Продолжительность данной фенофазы является одной из важнейших характеристик декоративности сиреней (Fiala, 2008). Продолжительность цветения S. josikaea в условиях арктического Мурманска составляет 8–16 суток (см. табл. 3), в первый год мониторинга оно было более длительным (13–16 суток), во второй – более коротким (8–12 дней). Исследования флорального органогенеза S. josikaea показало, что заложение и дифференциация зачатков цветков и соцветий в изучаемых ценопопуляциях г. Мурманска происходит в разное время: в контроле, на площадках ПП2 и ПП4 – осенью 2014 г., а на ПП2 и ПП3 – весной 2015 г. (Василевская, Морозова, 2017). При этом поливариантность флорального органогенеза не влияет на сроки начала и окончания цветения, его продолжительность.
Завязывание плодов у S. josikaea отмечено 16–19 июля (см. табл. 1, 3). Изменение окраски плодов от зеленого к коричневатому оттенку свидетельствует об их созревании, оно наблюдается в г. Мурманске значительно позже, к окончанию листопада. Листья держатся на побегах долго, расцвечивание начинается с появления коричневатого или желтоватого оттенка. Для большинства видов секции Villosae C. K. Schneid (сирени волосистые), к которой относится сирень венгерская, характерно только частичное пожелтение листьев. Начало фазы расцвечивания листьев (см. табл. 1) различается по годам: в 2014 г. – 14–20 сентября, в 2015 г. – 25–27 сентября. Листопад S. josikaea отмечен 3–8 октября при стабильно низких положительных температурах. Полное опадение листьев у S. josikaea происходит после выпадения осадков в виде снега, при этом листья очень долго могут находиться на побегах, нередки случаи, когда единичные листья могут оставаться на побегах в течение зимы.
Исследования показали, что продолжительность вегетационного периода S. josikaea на широте г. Мурманска 125–136 суток (см. табл. 3). Это значительно меньше, чем в других северных городах России. У зимостойких видов сезонное развитие протекает в более короткие сроки и, как правило, завершается в период с температурой выше +5 °С (Бабич и др., 2008). По-видимому, количество календарных дней безморозного периода вполне достаточно S. josikaea для прохождения всех этапов вегетативного и репродуктивного развития.
Обсуждение
С введением растений в новые места обитания изменяется не только их жизненная форма, но и период вегетации, сдвигаются сроки начала и окончания определенных фенологических фаз. Мониторинг роста и развития сирени венгерской в г. Мурманске показал, что в условиях холодного и короткого арктического лета происходит сокращение периода вегетации, значительное укорочение ряда фенофаз, их смещение на более поздние сроки.
Известно, что срок наступления вегетации – очень изменчивый признак, в значительной степени зависящий от условий среды (Бабич и др., 2008). В Мурманске вегетация S. josikaea начинается в конце мая – начале июня, что значительно позже, чем в городах России, расположенных в подзонах северной и средней тайги. В 2014 г. распускание почек происходило при более низких значениях суммы активных температур, чем в 2015 г. При этом на ряде пробных площадок сирень начинала вегетировать при значениях САТ 81.8–87.3 °С. В г. Тюмени развертывание почек S. josikaea отмечено в разные годы при суммах активных температур 109–191 °С (Герасимова, 2015). Это подтверждает данные о том, что в северных широтах деревья и кустарники могут проходить начальные фенофазы при более низких температурах, чем в более южных регионах (Бабич и др., 2008). Некоторые авторы (Видякина, Семенова, 2009) считают, что различия в сроках развертывания почек по годам связаны с колебаниями весенних среднесуточных температур. Исследования Syringa vulgaris в Европе показали, что величина изменчивости температуры перед началом вегетации является существенным фактором, влияющим на весеннюю фенологию (Wang et al., 2018). Возможным механизмом является то, что растения ослабляют свои фенологические реакции, чтобы избежать риска замерзания в климате с более высоким уровнем изменчивости весенних температур.
Начало линейного роста побегов S. josikaea происходит синхронно 3 июня на всех пробных площадках г. Мурманска в оба года мониторинга, что на две – три недели позже, чем в городах, расположенных в подзонах средней и северной тайги. В 2014 г. эта фенофаза началась при сумме положительных температур 163 °С, в 2015 г. при 248 °С. В Тюмени побеги сирени венгерской трогаются в рост при САТ 196–288 °С (Герасимова, 2015). Н. А. Бабич (2008) отмечает, что начало роста побегов интродуцентов определяется особенностями их биологии и суммой весенних эффективных температур. По мнению Н. В. Поляковой (2009), наступление фенофаз начала развертывания почек и роста побегов у сиреней в наименьшей степени контролируется генетическими особенностями вида.
Фаза роста побегов S. josikaea на пробных площадках г. Мурманска отличается своей непродолжительностью (33–40 суток) по сравнению с другими северными регионами. Сроки окончания роста побегов сирени, по данным Н. В Поляковой (2011), зависят от видовых особенностей и мало связаны с условиями года. Факт интенсивного роста вегетативных побегов S. josikaea в условиях арктического города представляет особый интерес, поскольку при достаточно низких температурах вегетационного сезона за короткий промежуток времени побеги дают большой прирост. Исследованиями сотрудников Полярно-альпийского ботанического сада установлено, что для интродуцированных зимостойких кустарников на Кольском полуострове характерна неустойчивая и высокая скорость роста годичных побегов (Жиров, Гонтарь, 2009), обусловленная круглосуточным освещением в условиях полярного дня и непрерывным фотосинтезом в течение всего вегетационного периода. В условиях Кольской Арктики S. josikaea отличается высокой газоустойчивостью (Салтан и др., 2017), и даже при высоком уровне техногенного загрязнения окружающей среды в ее ассимилирующих органах наблюдается увеличение фотосинтетической активности. Выявленное ингибирование роста побегов сирени в центре г. Мурманска (ул. Ленинградская – ПП2) свидетельствует о снижении показателей вегетативного роста у старовозрастных экземпляров в условиях антропогенного загрязнения среды. Сквер на ул. Ленинградской расположен рядом с железнодорожным транспортным узлом, автомагистралью и Мурманским морским торговым портом, где осуществляется дробление и перевалка угля на экспорт. Аналогичные результаты получены при исследовании динамики роста побегов Sorbus gorodkovii (Василевская, Лебедевич, 2016). Многие виды сиреней являются фотопериодически нейтральными (Körner, Basler, 2010), именно эта особенность сирени венгерской обусловила ее успешное цветение в условиях арктического полярного дня. Цветение S. josikaea в г. Мурманске начинается в первой декаде июля, что на две – три недели позже, чем в городах, расположенных в подзонах северной и средней тайги (Северодвинск, Сыктывкар, Петрозаводск), и на полтора месяца позже, чем в подзоне южной тайги (Тюмень). В Мурманске в 2014 г. цветение S. josikaea началось при сумме активных температур 429–445 °С, в то время как в 2015 г. синхронное цветение на всех пробных площадках отмечено при САТ 595–602 °С. В Тюмени сирень венгерская зацветает при САТ 612–727 °С в зависимости от года (Герасимова, 2015). Показано, что у сиреней величина температурной чувствительности начала цветения уменьшается по мере продвижения в более высокие широты (Cleland et al., 2012; Wang et al., 2015). Длительность цветения S. josikaea в г. Мурманске составляет 8–16 суток, что близко к данным ряда авторов (Бабич и др., 2008; Мартынов, 2013; Кищенко, Потапова, 2014). Несмотря на наличие неблагоприятных факторов, влияющих на рост и развитие сирени в районах ее интродукции, они практически не сказываются на цветении (Чащухина, 1970). В условиях Севера длительность периода цветения интродуцированных видов не меньше, чем в более благоприятных климатических условиях (Бабич и др., 2008).
Листопад у сирени венгерской на широте Мурманска начинается в начале октября. На основе дисперсионного анализа Н. В. Поляковой (2011) показано, что равное влияние на начало опадения листьев у представителей рода Syringa L. оказывают как видовые особенности, так и климатические факторы. При сохраняющейся генетической обусловленности наступления этой фенофазы значительно возрастает роль климатических факторов, непосредственно влияющих на длительность сохранения листвы (перепады температур, осадки, интенсивность ветра).
Известно, что период вегетации интродуцированных растений на Севере значительно короче, чем на их родине и в других географических районах (Бабич и др., 2008). В г. Мурманске период вегетации S. josikae непродолжителен и составляет 126–136 суток. Укороченность вегетационного периода S. josikaea в Мурманске обусловлена непродолжительным и холодным арктическим летом на данной приполярной широте. При этом сирень венгерская способна пройти все фенологические фазы, несмотря на сокращение вегетационного периода.
Заключение
В условиях атлантико-арктического климата у сирени венгерской происходит значительное укорочение фенофаз, их смещение на более поздние даты по сравнению с другими городами севера России. Развертывание почек, рост побегов, начало и окончание цветения, листопад S. josikaea в условиях холодного лета и полярного дня осуществляются практически синхронно в разных районах г. Мурманска, при суммах активных температур, значительно различающихся по годам. При этом фенофазы могут проходить при минимальных значениях суммы активных температур. Это может свидетельствовать о том, что процессы развития S. josikaea в значительной степени детерминированы генетически. Сроки прохождения растениями основных фенофаз зависят не только от климатических условий, но в равной степени от биологических особенностей видов, сложившихся под влиянием естественной среды обитания и географического происхождения последних (Пенкина, 1978; Сунцова, Иншаков, 2007). Известно, что S. josikaеа – реликтовый вид, в естественных условиях произрастающий в горных районах. В. Л. Комаров (1940) предполагал, что Syringa L. является родом третичного периода, который в свое время был широко распространен в Старом Свете, в период повышенной геологической активности ареал рода был оттеснен на юг. Согласно М. Г. Попову (1949), виды рода Syringa L. сформировались в пребореальных мезотермальных лесах, образованных на северной окраине арктотретичной флоры, отступающих к югу под влиянием плейстоценового ледника. Сирени венгерской свойственны высокая экологическая пластичность и устойчивость, она является видом с широкой нормой реакции, поскольку успешно вегетирует и цветет в условиях полярного дня и низких температур арктического региона.
Библиография
Александрова М. С., Головкин Б. Н. Переселение деревьев и кустарников на Крайний Север . Л.: Наука, 1978. 114 с.
Бабич Н. А., Залывская О. С., Травникова Г. И. Интродуценты в зеленом строительстве северных городов . Архангельск: Изд-во АГТУ, 2008. 143 с.
Богданец Т. П., Василевская Н. В., Коммандер Я., Шевченко А. В. Экология Мурманской области с основами общей экологии . Мурманск: Милори, 1998. 188 c.
Бродер Б. М. Некоторые итоги озеленения г. Мурманска // Флористические исследования и зеленое строительство на Кольском полуострове. Апатиты: Изд-во Кольского филиала АН СССР, 1975. С. 148–161.
Булыгин Н. Е., Ярмишко В. Т. Дендрология . М.: МГУЛ, 2001. 528 с.
Василевская Н. В., Лебедевич И. Е. Воздействие техногенного загрязнения г. Мурманска на рост и развитие Sorbus gorodkovii Pojark // Естественные и технические науки. 2016. № 4 (94). С. 23–28.
Василевская Н. В., Морозова Д. А. Тератоморфизм пыльцы Syringa josikaea Jacq. при интродукции на урбанизированных территориях Российской Арктики // Ученые записки Петрозаводского государственного университета. Сер.: Биологические науки. 2016. № 8 (161). С. 7–13.
Василевская Н. В., Морозова Д. А. Динамика флорального органогенеза Syringa josikaea Jacq. при интродукции в условиях арктического климата г. Мурманска // Ученые записки Петрозаводского государственного университета. Серия: Биологические науки. 2017. № 4 (165). С. 14–21.
Видякина А. А., Семенова М. В. Фенологические наблюдения за развитием вегетативных и генеративных побегов Syringa josikaea Jacq. в различных районах г. Тюмени // Вестник экологии, лесоведения и ландшафтоведения. 2008. № 9. С. 142–145.
Герасимова А. А. Фенологический мониторинг древесно-кустарниковой растительности г. Тюмени: Дис. ... канд. биол. наук . Тюмень, 2015. 273 с.
Гонтарь О. Б., Жиров В. К., Казаков Л. А., Святковская Е. А., Тростенюк Н. Н. Зеленое строительство в городах Мурманской области . Апатиты: Изд-во КНЦ РАН, 2010. 292 с.
Гонтарь О. Б., Святковская Е. А., Тростенюк Н. Н., Жиров В. К. Этапы создания и анализ особенностей ассортимента декоративных древесных растений для оптимального озеленения урбанизированных территорий Крайнего Севера // Вестник МГТУ. 2011. Т. 14. № 3. С. 577–582.
Гонтарь О. Б., Святковская Е. А., Тростенюк Н. Н., Корoбейникова Н. М., Шлапак Е. П., Носатенко О. Ю. Мониторинг состояния древесных насаждений на некоторых объектах озеленения в центральной части города Мурманска // Известия Самарского научного центра РАН. 2013. Т. 15. № 3 (2). С. 621–625.
Гонтарь О. Б., Макарова О. А., Дудакова Е. Ф. Скверы города Мурманска – экологические маршруты для изучения природы и истории родного края // Экологическое краеведение. Ишим, 2016. С. 146–152.
Демин В. И. Основные климатические тенденции на Кольском полуострове за период инструментальных метеорологических измерений // Труды Кольского НЦ РАН. 2012. Т. 1. С. 99–110.
Доклад о состоянии и об охране окружающей среды Мурманской области в 2015 году . URL: http://mpr.gov-murman.ru (дата обращения: 12.02.2020).
Жиров В. К., Гонтарь О. Б. Интродукция растений как приоритетное направление научной и практической деятельности Полярно-альпийского ботанического сада-института // Вестник Кольского НЦ РАН. 2009. № 1. С. 34–44.
Кищенко И. Т., Потапова М. Н. Развитие интродуцированных видов Syringa (Oleacea) в условиях Карелии // Ученые записки Петрозаводского государственного университета. 2014. № 2. С. 16–18.
Комаров В. Л. Учение о виде у растений . М.; Л.: Изд-во АН СССР, 1940. 212 с.
Логинов В. Б. К методике построения частных теорий интродукции // Теории и методы интродукции растений и зеленого строительства. Киев, 1980. С. 58–60.
Мартынов Л. Г. Интродукция видов рода Сирень (Syringa) в условиях ботанического сада подзоны средней тайги // Известия Коми научного центра УрО РАН. 2013. № 4. С. 25–31.
Морозова Д. А. Фенология роста и развития Syringa josikaea Jacq. в условиях арктического города (на примере г. Мурманска) // Научное сообщество студентов XXI столетия. Естественные науки. Новосибирск, 2018. № 11 (69). С. 21–28.
Обзор метеорологических условий // Мурманское управление по гидрометеорологии и мониторингу окружающей среды. URL: http://www.kolgimet.ru (дата обращения: 01.09.2016).
Пенкина И. Г. Сирени в Чуйской долине . Фрунзе: ИЛИМ, 1978. 56 с.
Пенкина И. Г., Ткаченко В. И. Рост и развитие видов рода Syringa L. в Ботаническом саду г. Фрунзе // Известия АН Киргизской ССР. 1974. № 2. С. 67–73.
Переверзев В. Н. Почвы и почвенный покров Кольского полуострова: история и современное состояние исследований // Вестник Кольского НЦ РАН. 2011. № 1. С. 39–43.
Полякова Н. В. Интродукция сирени в Башкирском Предуралье // Естественные науки. 2009. № 2. С. 48–53.
Полякова Н. В. Сезонный ритм развития видов рода Syringa L. в г. Уфа // Вестник Иркутской государственной сельскохозяйственной академии. 2011. № 2 (44). С. 120–125.
Полякова Н. В., Путенихин В. П., Вафин Р. В. Сирени в Башкирском Предуралье. Интродукция и биологические особенности . Уфа: Гилем, 2010. 164 с.
Попов М. Г. Очерк растительности и флоры Карпат . М.: МОИП, 1949. 303 с.
Салтан Н. В., Шлапак Е. П., Жиров В. К., Святковская Е. А., Гонтарь О. Б., Тростенюк Н. Н. Изучение пластичности фотосинтетического аппарата листьев дендроинтродуцентов в условиях урбаносреды Кольской Арктики // Известия Самарского научного центра РАН. 2017. Т. 19. № 2 (3). С. 545–549.
Семенюк Н. Б., Долганова З. В. Сохранение и расширение генофонда сирени, создание зимостойких сортов для условий Западной Сибири // Достижения науки и техники АПК. 2008. № 7. С. 22–24.
Сунцова Л. Н., Иншаков Е. М. Древесные растения в условиях техногенной среды // Хвойные бореальной зоны. 2007. Т. 24. № 1. С. 95–99.
Чащухина А. А. Фенологические наблюдения над декоративными деревьями и кустарниковыми растениями // Бюллетень Ботанического сада (1957–1960). Владивосток: Изд-во ДВ НЦ АН СССР, 1970. С. 114–124.
Шлапак Е. П., Салтан Н. В., Святковская Е. А., Гонтарь О. Б., Жиров В. К. Морфологические аспекты адаптации древесных интродуцентов в урбанизированной среде Кольской Арктики [Morphological aspects of adaptation of tree introducers in the urbanized environment of the Kola Arctic // Экологическая, промышленная и энергетическая безопасность. Севастополь, 2017. С. 1572–1575.
Abakumov E., Shamilishviliy G., Yurtaev A. Soil polychemical contamination on Beliy island as key background and reference plot for Yamal region // Polish Polar Research Journal. 2017. Vol. 30. № 3. P. 313–332.
Chambers L. E., Altwegg R., Barbraud C., Barnard P., Beaumont L. J., Crawford R. J. M. Phenological changes in the Southern Hemisphere // PLoS One. 2013. Vol. 8. № 10. P. 1–12.
Cleland E. E., Allen J. M., Crimmins T. M., Dunne J. A., Pau S., Travers S. E. Phenological tracking enables positive species responses to climate change // Ecology. 2012. Vol. 93. № 8. P. 1765–1771.
DeLucia E. H., Nabity P. D., Zavala J. A. Climate change: Resetting plant-insect interactions // Plant Physiology. 2012. Vol. 160. P. 1677–1685.
Dihoru G., Negrean G. The Red book of vascular plants of Romania // Romanian Journal of Biology. 2009. Vol. 54. № 1. Р. 101–114.
Fiala N. Measuring sustainability: Why the ecological footprint is bad economics and bad environmental science // Ecological Economics. 2008. Vol. 67. № 4. P. 519–525.
Gill A. L., Gallinat A. S., Sanders-DeMott R., Rigden A. J., Gianotti D. J. S., Mantooth J. A., Templer P. H. Changes in autumn senescence in Northern Hemisphere deciduous trees: A meta-analysis of autumn phenology studies // Annals of Botany. 2015. Vol. 116. № 6. P. 875–888.
Körner C., Basler D. Phenology under global warming // Science. 2010. Vol. 327. № 5972. P. 1461–1462.
Lendvay B., Pedryc A., Holm M. Characterization of nuclear microsattelite markers for the narrow endemic Syringa josikaea Jacq. fil ex Rchb. // Notulae Botanicae Horti Arrobotanici cluy – Napoka. 2013. Vol. 41 № 1. P. 301–305.
Vasilevskaya N. V., Morozova D. A. Polyvariance of floral organogenesis of Syringa josikaea Jacq. (Oleaceae) in the conditions of the Arctic urbanized territory // Ecology and Sustainable Development. 2018. Vol. 1. № 1. P. 31–39.
Vasilevskaya N. V., Domachina A. D. Teratomorphism of pollen of Larix sibirica Ledeb. (Pinaceae Lindl.) in the Arctic urbanized territory // Czech Polar Reports. 2018. Vol. 8. № 1. P. 24–36.
Wang H., Ge. Q., Dai J., Tao Z. Geographical pattern in first bloom variability and its relation to temperature sensitivity in the USA and China // International Journal of Biometeorology. 2015. Vol. 59. P. 961–969.
Wang H., Dai J., Rutishauser T., Gonsamo A. Trends and variability of lilac flowering phenology // Journal of Geophysical Research: Biogeoscience. 2018. Vol. 123. № 3. P. 807–817.
Walker D. A., Deniels J. A., Matveeva N. V. et al. Circumpolar Arctic vegetation classification // Phytocoenologia. 2018. Vol. 48. № 2. P. 181–201.

 © 2011 - 2026
© 2011 - 2026