



Введение
Центрально-Черноземный район (ЦЧР) характеризуется высоким уровнем экологической напряженности (7-й ранг) (Гостева, 2010). В его больших городах изменены микроклимат, почвенный и растительный покровы, подземные воды. Во второй половине ХХ в. население Воронежа увеличилось вдвое. Часть лесных массивов из созданного в 1960-х гг. зеленого пояса (в основном лесные культуры сосны) вошла в черту город. Сосна обыкновенная очень чувствительна к техногенному стрессу (Лесные экосистемы…, 1990). Поэтому исследование экологического потенциала, адаптации и причин ослабления вида на урбанизированных территориях является актуальной задачей экологической генетики, физиологии и биохимии (Grey, Deneke, 1986; Чиркова, 2002; Brandt et al., 2016).
Целью работы является изучение на контрастном экологическом фоне структуры популяций, нормы реакции и ее сдвигов у сосны обыкновенной (Pinus sylvestris L.) по морфологическим и физиолого-биохимическим признакам генеративной и вегетативной сферы.
Материалы
Объектами исследования являлись два одновозрастных 53–55-летних насаждения сосны из экологически контрастных мест произрастания. Ступинский тест-объект (Воронежская область, Рамонской район, лесные культуры II класса бонитета, 30 деревьев, контроль) является объектом длительного мониторинга (1987–2020 гг.), типичное для лесостепной зоны по вегетативной и семенной продуктивности насаждение, прилегающее к Воронежскому биосферному заповеднику. Исследования на опытном объекте «Московский проспект» проводятся с 2007 г. Участок лесных культур находится в черте г. Воронежа (30 деревьев, опыт), вдоль крупной автотрассы М-4 «Дон», уровень эмиссионной нагрузки 6.02 т/год на км2 (Куролап, Клепиков, 2013). За годы мониторинга в регионе произошло 8 засух (1991, 1995, 2001, 2007, 2010, 2012, 2014, 2019 гг.), 3 года были холодными (1990, 1994, 2016), остальные соответствовали категории оптимальных.
Методы
Интенсивность семеношения сосны оценивалась по 5-балльной шкале В. Г. Каппера, полнозернистости семян (%) и числу семян в шишке (шт.). Полнозернистость вычисляли как соотношение числа полных семян к общему их числу (полные + пустые). Для оценки полиморфизма деревьев и определения частотного распределения генотипов диапазон их изменчивости разбит на классы продуктивности в следующих границах: по полнозернистости семян: I – 0.0–24.9 %: II – 25.0–49.9 %; III – 50.0–74.9 %; IV – 75 % и выше; по числу семян в шишке: I – 0.0–9.9; II – 10.0–19.9; III – 20.0–29.9; IV – 30.0 и больше.
Для физиолого-биохимических исследований в мае с 11 модельных деревьев отобраны побеги второго года жизни и проведен анализ средней пробы хвои. Исследования водного режима растений (дефицит влаги, общее количество влаги, содержание коллоидно-связанной воды) проводили по общепринятой методике Х. Н. Починка (1976). Содержание свободного пролина определяли в воздушно-сухих пробах методом Bates et al. (1973). Для статистической обработки данных использовали программу Statistica 6.
Результаты
Ступинский тест-объект. Район исследований относится к зоне высокопродуктивных сосновых лесов (Kuznetsova et al., 2019). В оптимальные годы интенсивность семеношения сосны стабильно высокая (4 балла). Потенциальная норма реакции по полнозернистости семян составляет 0–100 %. Реализованная норма вдвое уже и имеет асимметричный характер. Модальные значения признака (79–86 %) находятся в области высокопродуктивного IV класса. Выборка оптимальных лет имеет однотипную структуру, при которой основная часть деревьев выборки относится к продуктивным III и IV классам (рис. 1).
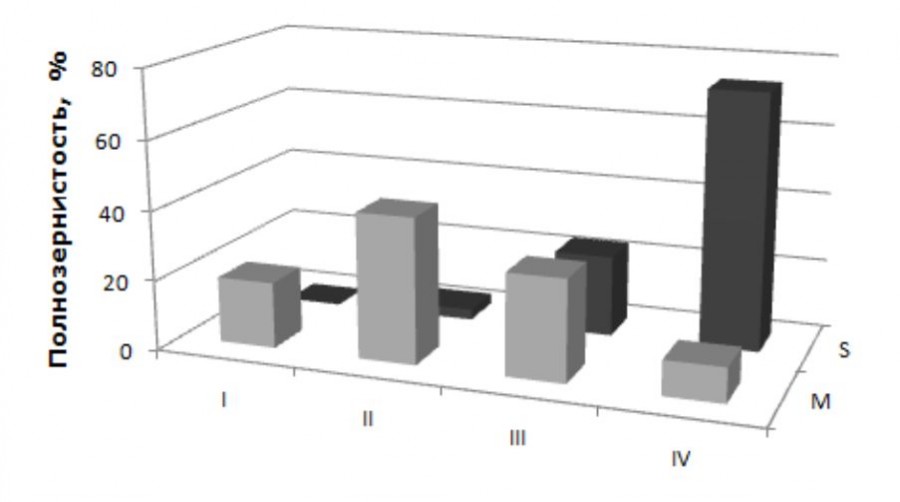
Рис. 1. Распределение деревьев сосны обыкновенной по полнозернистости семян в оптимальные годы: I класс – 0–24.9 %; II класс – 25.0–49.9 %; III класс – 50.0–74.9 %; IV класс – 75 % и выше. S – Ступинский тест-объект, M – Московский проспект
Fig. 1. Distribution of Scots pine trees by the proportion of plump seeds in optimal years: I class – 0–24.9 %; II class – 25.0–49.9 %; III class – 50.0–74.9 %; IV class – 75 % and above. S – Stupinsky test-objekt; M – Moscosky avenue
Потенциальная норма реакции по числу семян равна 0–50 семян. Модальные значения реализованной нормы признака (19.6–26.1 семени) расположены в центре вариационного ряда и, как правило, в зоне деревьев III класса продуктивности. Максимальный размах амплитуды изменчивости составляет 77.2 % (5.2–43.8 семени) от потенциальной. В структуре выборки представлены деревья всех классов – от I до IV (рис. 2). Число низкопродуктивных и высокопродуктивных деревьев невелико, но последних больше. Поскольку внутрипопуляционная выборка в оптимальные годы существенно не меняется, то групповой вклад классов в семенную продукцию года приблизительно одинаковый.
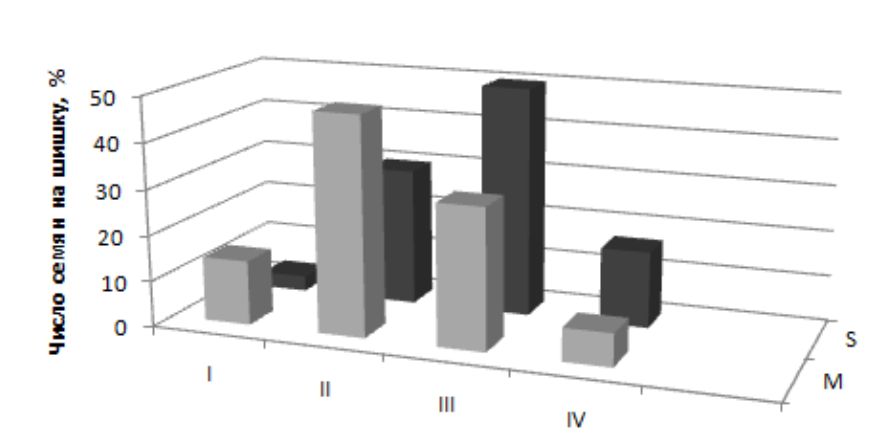
Рис. 2. Распределение деревьев сосны обыкновенной по числу семян на шишку в оптимальные годы: I класс – 0.0–9.9; II класс – 10.0–19.9; III класс – 20.0–29.9; IV класс – 30.0 семени и больше. S – Ступинский тест-объект, M – Московский проспект
Fig. 2. Distribution of Scots pine trees by the number of seeds per cone in optimal years: I class – 0.0–9.9; II class – 10.0–19.9; III class – 20.0–29.9; IV class – 30.0 and more. S – Stupinsky test-objekt; M – Moscovsky avenue
Подробный анализ физиолого-биохимических показателей ступинской популяции дан в предыдущей работе (Клушевская, Кузнецова, 2016). Здесь мы приводим краткую информацию, необходимую для сравнительного анализа. Установлено, что средние значения уровня влаги в свежесобранных образцах хвои составляют 55.3 + 1.11 %. Размах изменчивости признака небольшой – в пределах 11.6 % (50.7–62.3 %). Уровень дефицита влаги – 8.7 + 0.81 %. Амплитуда изменения признака составляет всего 6.6 % (6.0–12.6 %). Количество коллоидно-связанной воды – 19.1 + 1.65 %. Амплитуда изменчивости признака 16.9 %, максимальные значения достигают 27.7 %. Для деревьев контроля характерен в среднем невысокий уровень пролина – 2.96 + 0.27 мкМ/г, который колеблется в достаточно узких пределах (1.84–4.53 мкМ/г) (рис. 3).
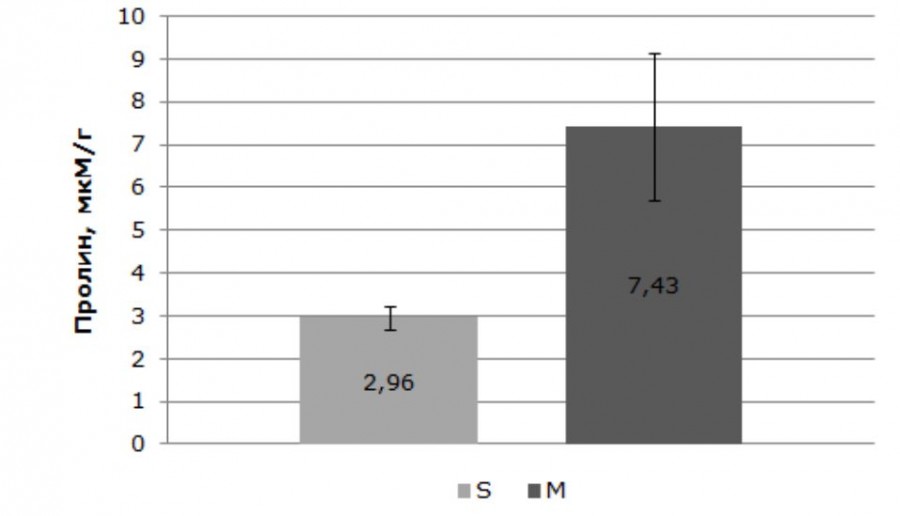
Рис. 3. Содержание пролина в хвое сосны обыкновенной в оптимальные годы. S – Ступинский тест-объект, M – Московский проспект
Fig. 3. The proline content in the needles of Scots pine in optimal years. S – Stupinsky test-objekt; M – Moscovsky avenue
Объект «Московский проспект». Ювенильный период формирования насаждения «Московский проспект» пришелся на относительно нормальные экологические условия. После того как в середине 1970-х гг. данный лесной массив вошел в городской округ Воронежа, его развитие продолжалось под прессингом постоянно нарастающего техногенного стресса.
По данным мониторинговых исследований, интенсивность семеношения большинства деревьев Московского проспекта ниже региональной нормы – 2–3 балла. Шишки, как правило, более мелкие по размеру. Средний уровень полнозернистости семян составляет 69.1 + 3.0 %. Значения варьируют в границах 62.8–78.9 %, что на 15–20 % ниже региональной нормы. Реализованная норма реакции вдвое шире ступинской и по амплитуде приблизилась к потенциальной (0.0–93.7 %). Изменилась структура выборки за счет появления низкопродуктивного I класса и резкого увеличения доли деревьев II класса (см. рис. 1). Модальные значения второго признака – число семян на шишку – ниже региональной нормы на треть: 16.4 + 1.4 шт. (в пределах 13.6–17.2 семени) и находятся в зоне II класса (см. рис. 2). Реализованная норма реакции смещена к левому пределу (0.0–37.4 семени), амплитуда изменения признака близка к Ступино (74.8 %). Здесь следует учитывать, что пустосемянность у сосны обусловлена гибелью зародышей в раннем эмбриогенезе. Редукция числа семян связана с летальностью семяпочек до оплодотворения из-за недоопыления и нарушения проэмбриогенеза.
Уровень влаги в свежесобранной хвое деревьев выше, чем на контрольном объекте, – 60.5 + 2.82 %, что, по-видимому, обусловлено его природой. В литературе описаны случаи увеличения содержания влаги в хвое сосны обыкновенной по мере приближения к источникам загрязнения как приспособительная реакция организма, направленная на детоксикацию избыточного содержания тяжелых металлов (Кизеев и др., 2009). По содержанию влаги деревья очень разнородны. Амплитуда изменчивости признака составляет 31.3 % (40.2–71.6 %), что в 2.7 раза больше, чем в Ступино (11.6 %) (см. рис. 2). Величина дефицита влаги – 16.2 + 1.83 %, что выше контроля в 1.9 раза. Причем если минимальные значения признака сопоставимы (5–6 %), то максимальные отличаются вдвое: 12.6 % в Ступино, 25.6 % – на Московском проспекте. Ранее показано, что атмосферное загрязнение способно повышать дефицит влаги растений за счет роста скорости водоотдачи и понижения водоудерживающей способности хвои сосны (Прожерина, 2006). Еще одним анализируемым показателем является содержание коллоидно-связанной воды. Ее уровень у деревьев опытного варианта резко, в 3.9 раза, повышен (75.3 + 2.37 %). Нижняя граница признака равна 58.9 %, что на 1/3 выше верхнего его предела в контроле.
Свободный пролин является стрессовым метаболитом, его концентрация свидетельствует о текущем состоянии растения и прямо зависит от факторов окружающей среды (Кузнецов, Шевякова, 1999; Verbruggen, Hermans, 2008). Установлено, что содержание пролина в хвое деревьев с Московского проспекта превышает контроль в 2.5 раза (7.43 + 1.73 мкМ/г). Для них характерна широкая область распределения признака (1.45–15.38 мкМ/г) (см. рис. 3). Ранее для сосны и ели показано увеличение пролина в хвое при атмосферном загрязнении (Дорофеева, 1981).
Обсуждение
В оптимальные годы ступинская популяция находится в состоянии, близком к равновесию, характеризуется упорядоченностью процессов, оптимальной изменчивостью и устойчивостью. Особенностью таких лесных экосистем является то, что они всегда после возмущения (в пределах адаптационных возможностей вида) возвращаются в исходное положение – региональной нормы. Для поддержания упорядоченности им необходима энергия. В зоне высокопродуктивных сосновых лесов, куда входит район исследований, энергетических ресурсов у сосны в системах «вид – среда» и «генотип – среда» достаточно для сбалансированного развития вегетативной и генеративной сферы, что подтверждается экспериментальными данными.
На техногенно загрязненной территории Московского проспекта биологические и экологические требования сосны обыкновенной и параметры среды обитания совпадают лишь частично. Растительные сообщества тратят часть своей энергии на детоксикацию внутренней среды и сохранение гомеостаза. Поэтому они вынуждены не просто приспосабливаться к хронической стрессовой нагрузке, но использовать наименее энергозатратные пути выживания и жизнеобеспечения. Один из таких путей – переход в качественно иное жизненное состояние – слабо неравновесной системы. Для неравновесных систем при отсутствии внешнего воздействия характерен более низкий уровень упорядоченности. При половом воспроизведении избыточная изменчивость частично нивелируется путем элиминации наиболее отклоняющихся генотипов, что проявляется в виде пустосемянности и уменьшения числа семян в шишках.
При смене одного устойчивого состояния на другое неизбежно происходит преобразование предшествующей структуры и функций (Раутиан, 1993). Во многом это происходит посредством реорганизации метаболических процессов, усиления репаративной способности растений, расширения нормы реакции (Чудинова, Орлова, 2006). При частичном дисбалансе генотипы имеют больше степеней свободы, нормы реакции перекрываются, что обеспечивает их устойчивость к дополнительной стрессовой нагрузке. Однако переход в данное функциональное состояние сопряжен с падением качества как лесных экосистем, так и их семенного потомства. При нормализации среды данное состояние обратимо. Возвращение сосны к динамическому равновесию происходит в ходе полового размножения в системе «родители – потомки» (Кузнецова, Машкина, 2011).
Заключение
Деревья и популяционные выборки насаждений сосны из экологически благоприятных (Ступинский тест-объект) и техногенно загрязненных (Московский проспект) мест обитания достоверно отличаются по экологически зависимым морфологическим и физиолого-биохимическим показателям, за исключением содержания влаги. При сравнении данных объектов наблюдается общая тенденция – расширение границ варьирования признаков в неблагоприятных условиях.
Морфологические признаки у анализируемых объектов имеют разную структурированность и адаптивную норму реакции. Повышенная изменчивость и тип трансформации структуры выборки в череде оптимальных лет не меняется, что свидетельствует о стабильности данных состояний. Вероятно, при хроническом техногенном загрязнении стрессоустойчивость вида обеспечивается переходом растений на менее энергозатратный путь развития – слабо неравновесной системы. Высвобождающаяся при этом энергия идет на поддержание жизнеспособности и выживание растений в аномальных условиях.
Библиография
Гостева С. Р. Экологическая безопасность России и устойчивое развитие // Вестник ТГТУ. 2010. Т. 16. № 3. С. 704–718.
Дорофеева Л. М. Индивидуальная изменчивость сосны обыкновенной по термостойкости // Исследование форм внутривидовой изменчивости растений. Свердловск: ИЭРиЖ УНЦ АН СССР, 1981. С. 81–90.
Кизеев А. Н., Жиров В. К., Никанов А. Н. Влияние промышленных эмиссий предприятий Кольского полуострова на ассимиляционный аппарат сосны // Экология человека. 2009. № 1. С. 9–14.
Клушевская Е. С., Кузнецова Н. Ф. Оценка устойчивости сосны обыкновенной к засухе по физиологическим характеристикам хвои // Лесоведение. 2016. № 3. С. 216–222.
Кузнецов В. В., Шевякова Н. И. Пролин при стрессе: биологическая роль, метаболизм, регуляция // Физиология растений. 1999. Т. 46. № 2. С. 321–336.
Кузнецова Н. Ф., Машкина О. С. Реакция на стресс и ее последействие у сосны обыкновенной в онтогенезе и при смене поколений // Хвойные бореальной зоны. 2011. Т. XXVIII. № 1–2. С. 83–90.
Куролап С. А., Клепиков О. В. Интегральное медико-экологическое зонирование как основа региональной стратегии устойчивого развития Воронежского региона // Вестник ТГУ. 2013. Т. 18. Вып. 2. С. 516–519.
Лесные экосистемы и атмосферное загрязнение / Под ред. А. К. Алексеева. Л.: Наука, 1990. 200 с.
Починок Х. Н. Методы биохимического анализа растений // Методы исследования водного режима растений. Киев: Наукова Думка, 1976. С. 318–324.
Прожерина Н. А. Адаптация хвойных к аэротехногенному загрязнению в районе Архангельской промышленной агломерации // Вестник Северного (Арктического) федерального университета. Сер.: Естественные науки. 2006. № 2. С. 77–82.
Раутиан А. С. О природе генотипа и наследственности // Журнал общей биологии. 1993. Т. 54. № 2. С. 13–148.
Чиркова Т. В. Физиологические основы устойчивости растений . СПб.: СПбГУ, 2002. 244 с.
Чудинова Л. А., Орлова Н. В. Физиология устойчивости растений . Пермь: Изд-во Пермского университета, 2006. 124 с.
Bates L. S., Waldren R. P., Teare I. D. Rapid determination of free proline for water stress studies // Plant Soil. 1973. Vol. 39. № 1. P. 205–207. DOI: 10.1007/BF00018060.
Brandt L., Levis A. D., Fahey R., Scott L., Darling L., Swanston C. A framework for adapting urban forests to climate change // Environment Science and Policy. 2016. Vol. 66. P. 393–402. DOI: 10.1016/j.envsci.2016.06.005.
Grey G. W., Deneke F. J. Urban Forestry. 2nd Ed. New York: John Wiley and Sons, 1986. 299 p.
Kuznetsova N. F., Semenov M. A., Sautkina M. Yu. Pine forests of East European plain / Pinus: Growth, Distribution and Uses. New York: Nova Science Publ., 2019. P. 1–47.
Verbruggen N., Hermans C. Proline accumulation in plants: a review // Amino Acids. 2008. № 35. P. 753–759. DOI: 10.1007/s00726-008-0061-6.
Благодарности
Исследования выполнены в рамках госзадания Всероссийского института лесной генетики, селекции и биотехнологии.

 © 2011 - 2026
© 2011 - 2026