



Введение
Исследование изменчивости как общебиологического явления имеет ряд основных аспектов. Исторически наиболее традиционными являются исследования морфологического разнообразия организмов, связанные с проблемами систематики и таксономии. Исследования внутривидовой изменчивости в связи с антропогенными изменениями среды обитания особенно широко развиваются последние десятилетия. Одним из новых направлений таких исследований является изучение флуктуирующей асимметрии билатеральных признаков. Особенностью этого направления является анализ особой формы изменчивости – внутрииндивидуального разнообразия как проявления случайной изменчивости развития (Захаров, 1987). Уровень флуктуирующей асимметрии билатеральных признаков является характеристикой общей стабильности развития, понимая под этим наличие стабилизированного потока, траектории развития (Waddington, 1940; Mather, 1953). Являясь показателем случайных отклонений в развитии, т. е. стабилизированного развития (Mather, 1953), флуктуирующая асимметрия одновременно выступает неспецифическим показателем условий развития, что дает возможность использовать ее для оценки условий существования как естественных, так и искусственных популяций (Захаров, 1987). Одним из популярных современных подходов оценки популяционного разнообразия является оценка уровня флуктуирующей асимметрии, которая позволяет определить не только гетерогенность популяций из разных участков города, но и стабильность развития особей (Захаров, 1987).
В настоящем исследовании проведен статистический анализ асимметрии билатерально симметричных признаков фолидоза головы и конечностей плоскохвостого домового геккона Hemidactylus platyurus на территории крупных городов Юго-Восточной Азии (ЮВА) (Вьентьян, Пномпень, Бангкок).
Материалы
При выполнении настоящего исследования использованы данные, полученные в результате обработки материала, собранного авторами в 2011–2014 гг. на территории ЮВА. Всего анализировались 765 экземпляров из трех локалитетов: г. Бангкок (Таиланд) – 130 самок, 168 самцов и 73 ювенильных особей; г. Вьентьян (Лаос) – 82 самки, 79 самцов и 36 ювенильных особей; г. Пномпень (Камбоджа) – 83 самки, 91 самец и 23 ювенильных особей. В каждом городе выборки сделаны в пяти точках (рис. 1): первая – условном центре города (за него принимался район с многоэтажной застройкой, удаленный от края города и приблизительно совпадающий с географическим центром) и четыре точки в условных окраинах (район на окраине с малоэтажной застройкой, граничащий с незастроенными территориями: пустырями с древесной или травянистой растительностью или сельскохозяйственными культурами, промышленные зоны), расположенными по сторонам горизонта от центральной точки (север, юг, запад, восток). Материал собирался с учетом обитания на стенах построек и стволах деревьев.
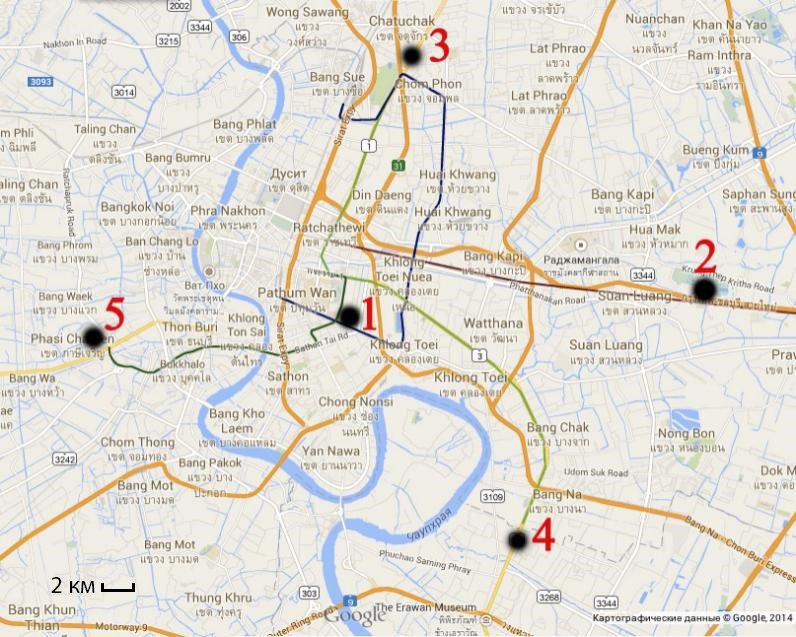
Рис. 1. Пример расположения точек сбора в городе (Бангкок). Условные обозначения: 1 – центр, 2 – восточная окраина, 3 – северная окраина, 4 – южная окраина, 5 – западная окраина
Fig. 1. An example of the location of sampling points in the city (Bangkok). Legend: 1 – center, 2 – eastern outskirts, 3 – northern outskirts, 4 – southern outskirts, 5 – western outskirts
В качестве изучаемых билатеральных признаков использовался фолидоз головы и число подпальцевых пластинок пальцев: Sl. – количество верхнегубных щитков; P. sl. – количество щитков следующего ряда, соприкасающихся с верхнегубными; Il. – количество нижнегубных щитков; G. il. – количество щитков следующего ряда, соприкасающихся с нижнегубными; Im – количество нижнечелюстных щитков; G. im. – количество следующего ряда щитков за нижнечелюстными; Ds – количество парных подпальцевых пластинок; Ads – количество непарных подпальцевых пластинок; Lm – количество подпальцевых пластинок.
Подсчет меристических признаков производился с применением бинокуляра МБС–10. Для удобства подсчета и дальнейшего анализа подпальцевых пластинок пальцам лап были присвоены порядковые номера: для передних – 1–5, для задних – 6–10.
Методы
Математическая обработка данных проведена с использованием электронных таблиц Microsoft Excel 2016 и пакета статистических программ STATISTICA, версия 10.0 (StatSoft, Inc., 2010, США). Проверка нормальности распределения переменных осуществлялась по критерию Колмогорова – Смирнова. Чтобы убедиться во флуктуирующем характере исследуемых признаков и возможности включения этих признаков в систему интегральной оценки ФА, был проведен статистический анализ: изучение направленности асимметрии признака, тест на идеальную ФА, изучение степени скоррелированности величины асимметрии разных признаков (Гелашвили и др., 2004). Для всех статистических тестов был установлен уровень значимости 0.05. Расчет интегрального индекса ФА производили по формуле (Васильев и др., 2006):
TA2 = [ ∑ (Li – Ri)] / k
DA2 = [ ∑ (Li – Ri)]2 / k2
FA2 = TA2 – DA2,
где L – значение признака слева, R – значение признака справа, k – число признаков, TA – дисперсия общей асимметрии, DA – направленная асимметрия, FA – флуктуирующая асимметрия.
Результаты
Проверка нормальности распределения
Для выбора статистических методов исследования на предварительном этапе был проведен анализ вида распределения асимметрии изучаемых признаков (L – R). Отклонения от нормального распределения зафиксированы для всех изучаемых признаков (табл. 1). Поэтому в дальнейшем использовались непараметрические методы статистического анализа.
Таблица 1. Проверка нормальности распределения значений асимметрии по критерию Колмогорова – Смирнова (KS)
| Признак | KS | p | Признак | KS | p | Признак | KS | p |
| Ds1 | 0.42 | <0.01 | Ds8 | 0.36 | <0.01 | Ds6 | 0.40 | <0.01 |
| Ads1 | 0.36 | <0.01 | Ads8 | 0.31 | <0.01 | Ads6 | 0.37 | <0.01 |
| Lm1 | 0.41 | <0.01 | Lm8 | 0.38 | <0.01 | Lm6 | 0.43 | <0.01 |
| Ds2 | 0.36 | <0.01 | Ds9 | 0.36 | <0.01 | Ds10 | 0.38 | <0.01 |
| Ads2 | 0.34 | <0.01 | Ads9 | 0.30 | <0.01 | Ads10 | 0.33 | <0.01 |
| Lm2 | 0.40 | <0.01 | Lm9 | 0.38 | <0.01 | Lm10 | 0.37 | <0.01 |
| Ds3 | 0.34 | <0.01 | Ds4 | 0.35 | <0.01 | Sl. | 0.27 | <0.01 |
| Ads3 | 0.31 | <0.01 | Ads4 | 0.33 | <0.01 | P.sl | 0.11 | <0.01 |
| Lm3 | 0.40 | <0.01 | Lm4 | 0.39 | <0.01 | Il. | 0.28 | <0.01 |
| Ds7 | 0.35 | <0.01 | Ds5 | 0.36 | <0.01 | G.il. | 0.13 | <0.01 |
| Ads7 | 0.33 | <0.01 | Ads5 | 0.33 | <0.01 | Im. | 0.40 | <0.01 |
| Lm7 | 0.40 | <0.01 | Lm5 | 0.38 | <0.01 | G.im. | 0.18 | <0.01 |
Проверка направленности асимметрии
Отсутствие направленности асимметрии выражается в том, что распределение различий признака на левой и правой сторонах является статистически однородным и приблизительно симметрично расположенным относительно нулевого значения. Проверку направленности ФА проводили с использованием рангового критерия сдвига Уилкоксона, т. е. в случае принятия нулевой гипотезы о статистической однородности показателей ФА (р > 0.05) принималось предположение о флуктуирующем характере асимметрии, а при альтернативе делался вывод о том или ином типе ее направленности (Гелашвили и др., 2007). Анализ направленности асимметрии признаков фолидоза H. platyurus показал (табл. 2), что статистически значимые различия между величинами признака на левой и правой сторонах тела наблюдаются по следующим признакам: количество непарных подпальцевых пластинок на 5-м пальце передней лапы (Ads5), количество подпальцевых пластинок на 1-м пальце задней лапы (Lm6), количество парных подпальцевых пластинок на 2-м пальце задней лапы (Ds7), количество щитков следующего ряда, соприкасающихся с верхнегубными (P. sl.), количество нижнегубных щитков (Il). Для остальных изучаемых признаков имеют место незначительные отклонения от билатеральной симметрии.
Таблица 2. Анализ направленности асимметрии признаков фолидоза H. platyurus (по критерию Уилкоксона)
| Признак | Z | p | Признак | Z | p | Признак | Z | p |
| Ds1 | –0.09 | 0.92 | Ds5 | –0.98 | 0.32 | Ds9 | –0.06 | 0.95 |
| Ads1 | –0.95 | 0.34 | Ads5 | –2.01 | 0.04 | Ads9 | –0.17 | 0.86 |
| Lm1 | –1.22 | 0.22 | Lm5 | –0.90 | 0.36 | Lm9 | –0.38 | 0.70 |
| Ds2 | –1.36 | 0.17 | Ds6 | –0.24 | 0.80 | Ds10 | –1.53 | 0.12 |
| Ads2 | –1.91 | 0.05 | Ads6 | –1.22 | 0.22 | Ads10 | –1.48 | 0.13 |
| Lm2 | –0.86 | 0.39 | Lm6 | –3.90 | <0.01 | Lm10 | –0.47 | 0.63 |
| Ds3 | –0.37 | 0.71 | Ds7 | –3.04 | <0.01 | Sl | –0.65 | 0.51 |
| Ads3 | –0.98 | 0.32 | Ads7 | –0.24 | 0.80 | P.sl | –3.21 | <0.01 |
| Lm3 | –1.95 | 0.05 | Lm7 | –0.75 | 0.44 | Il | –4.17 | <0.01 |
| Ds4 | –0.68 | 0.49 | Ds8 | –0.59 | 0.55 | G.il | –1.92 | 0.05 |
| Ads4 | –1.00 | 0.31 | Ads8 | –1.40 | 0.15 | Im | –0.43 | 0.66 |
| Lm4 | –0.58 | 0.56 | Lm8 | –0.95 | 0.33 | G.im | –1.01 | 0.31 |
Примечание. Жирным шрифтом обозначены статистически значимые значения.
Проверка на идеальную ФА
Тест на идеальную ФА включает проверку гипотезы о наличии у признаков явления антисимметрии. Явления ФА и антисимметрии не являются антагонистами и нечетко различимы в пределах одного и того же признака, однако выделение именно ФА имеет важное экологическое значение для оценки нормы генотипической гетерогенности организмов и стабильности их развития.
Индикатором антисимметрии служит отрицательный эксцесс (k) распределения различий между сторонами (L–R). При k ≥ 0 предположение о наличии антисимметрии отклоняется и принимается гипотеза о флуктуирующем характере асимметрии. Для всех исследуемых признаков значения эксцесса положительны (табл. 3), что позволяет исключить возможность проявления антисимметрии по выбранным признакам и считать, что наблюдаемая асимметрия носит флуктуирующий характер.
Таблица 3. Значения эксцесса распределения различий между сторонами (L – R) признаков фолидоза H. platyurus
| Признак | N | k | Признак | k | Признак | k |
| Ds1 | 765 | 3.4 | Ds5 | 11.1 | Ds9 | 14.5 |
| Ads1 | 765 | 2.0 | Ads5 | 0.5 | Ads9 | 0.8 |
| Lm1 | 765 | 4.3 | Lm5 | 12.6 | Lm9 | 21.1 |
| Ds2 | 765 | 6.4 | Ds6 | 4.4 | Ds10 | 3.4 |
| Ads2 | 765 | 0.9 | Ads6 | 2.1 | Ads10 | 1.5 |
| Lm2 | 765 | 26.9 | Lm6 | 9.6 | Lm10 | 19.9 |
| Ds3 | 765 | 11.4 | Ds7 | 8.6 | Sl. | 3.3 |
| Ads3 | 765 | 0.3 | Ads7 | 0.3 | P.sl | 4.1 |
| Lm3 | 765 | 15.1 | Lm7 | 28.9 | Il. | 1.2 |
| Ds4 | 765 | 12.8 | Ds8 | 11.5 | G.il. | 1.1 |
| Ads4 | 765 | 0.9 | Ads8 | 0.6 | Im. | 125.8 |
| Lm4 | 765 | 21.0 | Lm8 | 36.4 | G.im. | 1.2 |
Проверка корреляции
Для более точной и объективной оценки ФА предпочтительнее использовать не один какой-либо признак, а их ортогональный комплекс. Для оценки ортогональности изучаемых признаков необходимо выявить уровень их взаимной коррелированности между собой: чем меньше коррелируют эти признаки, тем более четкую и обоснованную оценку можно дать уровню обобщенной ФА. В случае если выявлена сильная корреляция между ними, необходим подбор других признаков для обобщенной оценки ФА. Для проверки корреляционной связи асимметрии признаков использовался ранговый коэффициент корреляции Спирмена. По большинству признаков была выявлена корреляция с разной степенью силы (рис. 2). Признаки, у которых сила корреляции превышала значение 0.7 и была статистически значимой, исключались из дальнейшего анализа. Превышение отмечено для следующих признаков: количество непарных подпальцевых пластинок на 1–5-м пальцах передней лапы (Ads1, Ads2, Ads3, Ads4, Ads5); количество непарных подпальцевых пластинок на 1–5-м пальцах задней лапы (Ads6, Ads7, Ads8, Ads9, Ads10); количество подпальцевых пластинок на 1-м и 4-м пальцах передней лапы (Lm1, Lm4); количество подпальцевых пластинок на 3, 4 и 5-м пальцах задней лапы (Lm8, Lm9, Lm10).
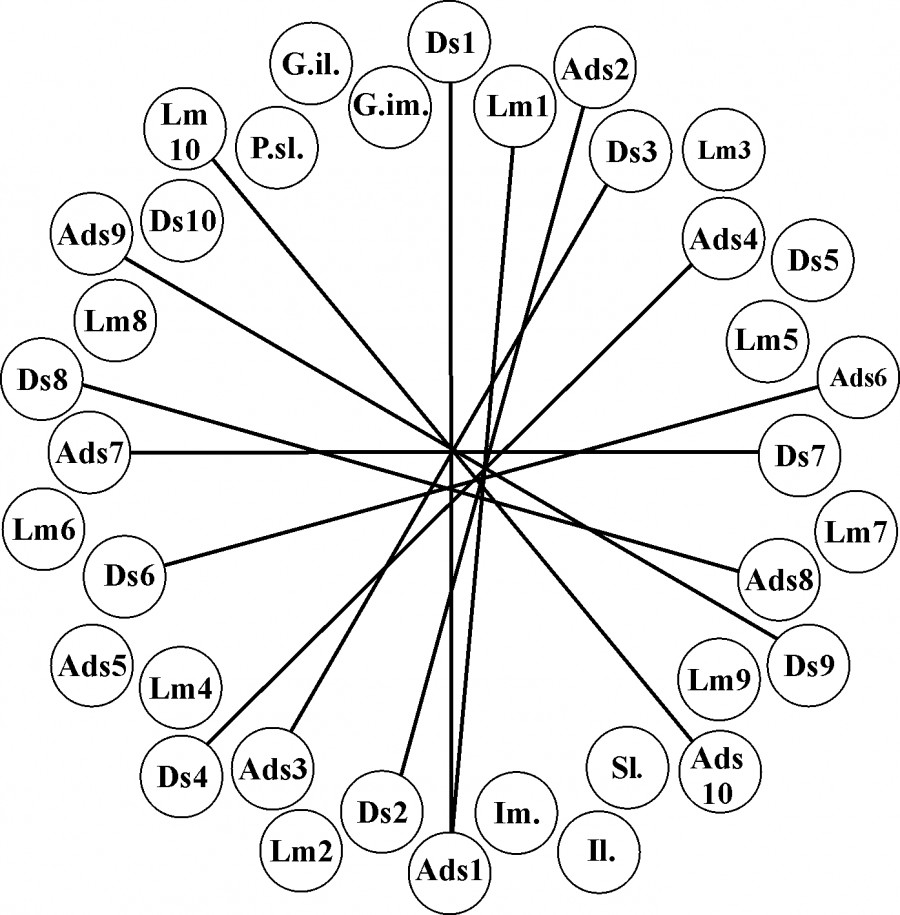
Рис. 2. Коррелограмма величины асимметрии признаков фолидоза |L – R|. Линиями обозначены сильно коррелирующие признаки (p < 0.05)
Fig. 2. Сorrelogram of the asymmetry of the folidosis signs | L – R |. The lines indicate strongly correlating traits (p < 0.05)
На основе проведенного статистического анализ исследуемых признаков фолидоза в систему оценки интегрального индекса ФА были включены следующие признаки: количество парных подпальцевых пластинок на 2-м и 5-м пальцах задней лапы (Ds8, Ds10), количество непарных подпальцевых пластинок на 2-м пальце задней лапы (ADs8), количество подпальцевых пластинок на 2, 3, 4 и 5-м пальцах передней лапы (Lm2, Lm3, Lm4, Lm5), количество подпальцевых пластинок на 1, 2, 3 и 4-м пальцах задней лапы (Lm6, Lm7, Lm8, Lm9), количество верхнегубных щитков (Sl.), количество щитков следующего ряда, соприкасающихся с верхнегубными (P. sl.), количество нижнегубных щитков (Il.), количество щитков следующего ряда, соприкасающихся с нижнегубными (G. il.), количество нижнечелюстных щитков (Im.), количество следующего ряда щитков за нижнечелюстными (G. im.).
Вклад отдельных признаков
Чтобы выяснить, какой вклад вносят отдельные признаки в интегральные показатели асимметрии H. platyurus из разных выборок, проанализировали частоту встречаемости особей, имеющих асимметрию по тому или иному признаку (табл. 4).
Таблица 4. Асимметричное проявление признаков
| Признак | Бангкок | Пномпень | Вьентьян | |||
| n | % | n | % | n | % | |
| Ds1 | 47 | 12.6 | 37 | 18.7 | 37 | 18.7 |
| Ds2 | 73 | 19.6 | 66 | 33.5 | 69 | 35.0 |
| Lm2 | 56 | 15.0 | 57 | 28.9 | 43 | 21.8 |
| Ds3 | 92 | 24.8 | 66 | 33.5 | 82 | 41.6 |
| Lm3 | 70 | 18.8 | 50 | 25.3 | 43 | 21.8 |
| Ds4 | 97 | 26.1 | 54 | 27.4 | 74 | 37.5 |
| Ds5 | 84 | 22.6 | 59 | 29.9 | 64 | 32.4 |
| Lm5 | 91 | 24.5 | 48 | 24.3 | 45 | 22.8 |
| Ds6 | 74 | 19.9 | 38 | 19.2 | 38 | 19.2 |
| Lm6 | 78 | 21.0 | 35 | 17.7 | 27 | 13.7 |
| Ds7 | 112 | 30.1 | 62 | 31.4 | 64 | 32.4 |
| Lm7 | 61 | 16.4 | 55 | 27.9 | 42 | 21.3 |
| Ds8 | 93 | 25.0 | 58 | 29.4 | 73 | 37.0 |
| Ds9 | 98 | 26.4 | 53 | 26.9 | 75 | 38.0 |
| Ds10 | 81 | 21.8 | 51 | 25.8 | 64 | 32.4 |
| Il. | 155 | 41.7 | 87 | 44.1 | 107 | 54.3 |
| G.il. | 266 | 71.7 | 152 | 77.1 | 154 | 78.1 |
| Im. | 56 | 15.0 | 69 | 35.0 | 23 | 11.6 |
| G.im. | 240 | 64.6 | 130 | 65.9 | 120 | 60.9 |
Для особей из Бангкока чаще характерна асимметрия: количество парных подпальцевых пластинок на 2-м пальце задней лапы (Ds7) – 30.1 %, количество нижнегубных щитков (Il.) – 41.7 %, количество щитков следующего ряда, соприкасающихся с нижнегубными (G. il), – 71.7 %, количество следующего ряда щитков за нижнечелюстными (G. im) – 64.6 %. В Пномпене: количество парных подпальцевых пластинок на 2-м пальце передней лапы (Ds2) – 33.5 %, количество парных подпальцевых пластинок на 3-м пальце передней лапы (Ds3) – 33.5 %, количество парных подпальцевых пластинок на 2-м пальце задней лапы (Ds7) – 31.4 %, количество нижнегубных щитков (Il.) – 44.4 %, количество щитков следующего ряда, соприкасающихся с нижнегубными (G. il), – 77.16 %, количество нижнечелюстных щитков (Im) – 35.0 %, количество следующего ряда щитков за нижнечелюстными (G. im) – 65.9 %. Во Вьентьяне: количество парных подпальцевых пластинок на 2-м пальце передней лапы (Ds2) – 35.0 %, количество парных подпальцевых пластинок на 3-м пальце передней лапы (Ds3) – 41.6 %, количество парных подпальцевых пластинок на 4-м пальце передней лапы (Ds4) – 37.5 %, количество парных подпальцевых пластинок на 5-м пальце передней лапы (Ds5) – 32.4 %, количество парных подпальцевых пластинок на 2-м пальце задней лапы (Ds7) – 32.4 %, количество парных подпальцевых пластинок на 3-м пальце задней лапы (Ds8) – 37.0 %, количество парных подпальцевых пластинок на 4-м пальце задней лапы (Ds9) – 38.0 %, количество парных подпальцевых пластинок на 5-м пальце задней лапы (Ds10) – 32.4 %, количество нижнегубных щитков (Il.) – 54.3 %, количество щитков следующего ряда, соприкасающихся с нижнегубными (G. il), – 78.1 %, количество следующего ряда щитков за нижнечелюстными (G. im) – 60.9 %.
В целом в трех изучаемых выборках у H. platyurus чаще встречается асимметрия количества нижнегубных щитков (Il.), количества щитков следующего ряда, соприкасающихся с нижнегубными (G. il), и количества следующего ряда щитков за нижнечелюстными (G. im).
Анализ интегрального показателя ФА
Возрастные и половые различия
Обоснованная интегральная оценка уровня ФА на всем массиве данных возможна при отсутствии влияния факторов, вызывающих гетерогенный характер объединяемых выборок и искажающих формируемые выводы. Одним из таких факторов могут быть половые и возрастные различия показателей асимметрии (Гелашвили и др., 2007).
Анализ проверки гипотезы наличия полового диморфизма показал отсутствие различий во всех трех анализируемых группах: Бангкок (U = 8759, Z = –0.20, p = 0.83), Пномпень (U = 2368, Z = 1.54, p = 0.12), Вьентьян (U = 2456, Z = 1.46, p = 0.14). Также не были выявлены возрастные различия в уровне ФА: Бангкок (U = 10089, Z = –0.07, p = 0.94), Пномпень (U = 1643, Z = –0.63, p = 0.52), Вьентьян (U = 2996, Z = –0.31, p = 0.75). Учитывая, что в анализируемых группах не выявлены ни половые, ни возрастные различия нарушения стабильности развития особей, дальнейший анализ групп проводился по объединенной выборке как самцов, и самок, как половозрелых, так и ювенильных особей.
Внутрипопуляционный анализ ФА
Для проверки гипотезы различий уровня флуктуирующей асимметрии провели сравнение выборок из центральной части города и «краевой» зоны. Используя объединенную выборку (без учета типа субстрата), мы не выявили значимых различий в нарушении стабильности развития (табл. 5).
Таблица 5. Результаты сравнения биотопов
| Биотоп | Бангкок | ||||
| e | n | s | w | c | |
| e | 0.22 | 0.96 | 0.51 | 0.59 | |
| n | 1358 | 0.32 | 0.06 | 0.36 | |
| s | 890 | 780 | 0.60 | 0.78 | |
| w | 1455 | 1241 | 835 | 0.16 | |
| c | 3804 | 3672 | 2214 | 3485 | |
| Пномпень | |||||
| e | 0.25 | 0.24 | 0.47 | 0.91 | |
| n | 200 | 0.90 | 0.78 | 0.20 | |
| s | 219 | 215 | 0.64 | 0.09 | |
| w | 263 | 228 | 242 | 0.28 | |
| c | 1009 | 667 | 688 | 839 | |
| Вьентьян | |||||
| e | 0.02 | 0.11 | 0.20 | 0.09 | |
| n | 169 | 0.72 | 0.38 | 0.42 | |
| s | 119 | 167 | 0.71 | 0.82 | |
| w | 129 | 149 | 103 | 0.90 | |
| c | 851 | 1029 | 693 | 705 | |
Примечание. Над диагональю – уровень значимости, под диагональю – U-статистика, жирным шрифтом выделены статистически значимые значения, c – центр, e – восточная окраина, n – северная окраина, s – южная окраина, w – западная окраина.
Во Вьентьяне в краевой зоне анализ сравнения выборок показал, что существуют статистически значимые различия между северной и восточной точками сбора материала.
При проверке гипотезы о различиях интегрального индекса с учетом типа субстрата во всех трех анализируемых совокупностях выявлены существенные различия (табл. 6). Причем разница зафиксирована как в одной точке сбора материала, так и при сравнении локалитетов между собой. В Бангкоке уровень флуктуирующей асимметрии в центральной зоне выше у особей, обитающих на стенах домов, по сравнению с особями, живущими на стволах деревьев.
Таблица 6. Результаты сравнения уровня флуктуирующей асимметрии с учетом субстрата
| Бангкок | ||||
| Субстрат | c-t | c-w | o-t | o-w |
| c-t | 0.03 | 0.44 | 0.06 | |
| c-w | 2004 | 0.15 | 0.78 | |
| o-t | 3228 | 2501 | 0.22 | |
| o-w | 3630 | 3517 | 4440 | |
| Пномпень | ||||
| c-t | <0.01 | <0.01 | 0.02 | |
| c-w | 544 | 0.92 | 0.66 | |
| o-t | 304 | 550 | 0.66 | |
| o-w | 914 | 1328 | 794 | |
| Вьентьян | ||||
| c-t | <0.01 | 0.01 | 0.01 | |
| c-w | 503 | 0.02 | <0.01 | |
| o-t | 777 | 719 | 0.69 | |
| o-w | 927 | 754 | 1089 | |
Примечание. Над диагональю – уровень значимости, под диагональю – U-статистика, жирным шрифтом выделены статистические значимые значения. c-t – центр – деревья, c-w – центр – стены, o-t – окраина – деревья, o-t – окраина – стены.
В Пномпене в центре города значение коэффициента ФА выше у гекконов, собранных на стенах домов. Анализ также показал, что выборка центр – деревья отличается более низким нарушением стабильности развития по сравнению с выборками окраина – деревья и окраина – стены. Наиболее ярко различия в уровне стабильности развития H. platyurus выражены во Вьентьяне.
Сравнение по типам субстрата показало высокозначимые различия в уровне нарушения стабильности развития в центральной и краевой зонах. Причем как на окраине, так и в центре города интегральный индекс ФА выше у ящериц, собранных со стен строений.
С увеличением доли доминирования H. frenatus снижается общий вклад H. platyurus в сообщество (наблюдается отрицательная корреляция на высоком уровне между этими двумя видами, r = –0.895). По нашим данным, в изученных локалитетах отношение к присутствию других видов в сообществе (Gehyra mutilata, Hemidactylus brookii, Dixonius siamensis) доминирующие в городских условиях H. frenatus и H. platyurus проявляют по-разному. У H. frenatus наблюдается толерантное отношение к присутствию других видов (r = 0.363), а вот H. platyurus показывает диаметрально противоположную картину – наблюдается отрицательная корреляция к наличию других видов в сообществе (r = –0.643).
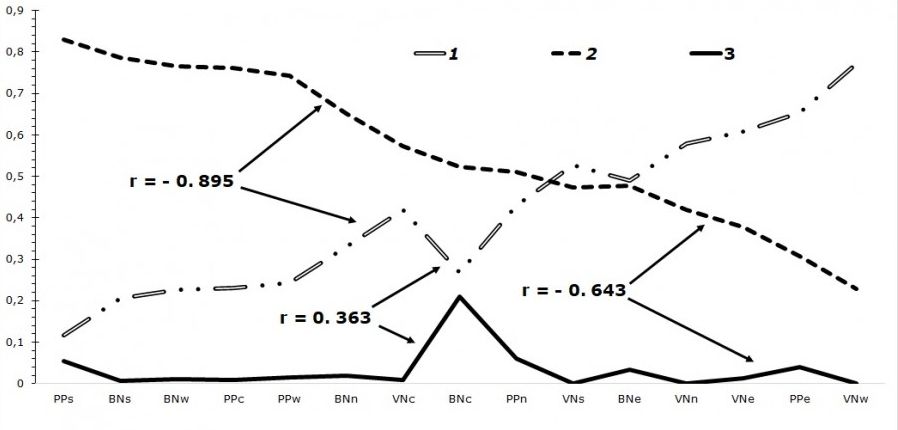
Рис. 3. Зависимость встречаемости синантропных гекконов, совместно обитающих на территории городов. D – индекс доминирования; BN – Бангкок; PP – Пномпень; VN – Вьентьян; c – центр города; n – северная окраина; s – южная окраина; w – западная окраина; e – восточная окраина, 1 – H. frenatus, 2 – H. platyurus, 3 – другие виды
Fig. 3. Dependence of the occurrence of synanthropic geckos living in cities together. Where, D is the dominance index; BN – Bangkok; PP – Phnom Penh; VN – Vientiane; c – center; n – northern outskirts; s – southern outskirts; w – western outskirts; e – eastern outskirts, 1 – H. frenatus, 2 – H. platyurus, 3 – other species
Межпопуляционные различия
При межпопуляционном сравнение выборок выявлены высоко значимые различия в индексе ФА между парами сравнения: Бангкок – Пномпень (U = 18999, Z = –3.59, p = < 0.01), Бангкок – Вьентьян (U = 22412, Z = –4.26, p = < 0.01). Разница в уровне ФА у особей между Пномпенем – Вьентьяном (U = 11512, Z = –0.49, p = 0.62) статистически не значима. Сравнение выборок в зависимости от типа биотопа выявило различия между центрами городов в Бангкоке и Вьентьяне. Различия в нарушении стабильности развития на окраине отмечены: Бангкок – Пномпень и Бангкок – Вьентьян. Разница в коэффициенте флуктуирующей асимметрии для Пномпеня – Вьентьяна не наблюдается ни в центре города, ни в краевой зоне (табл. 7).
Таблица 7. Результаты сравнения межпопуляционного распределения биотопов
| Биотоп | Центр | Биотоп | Окраина | ||||
| BN-c | PP-c | VN-c | BN-o | PP-o | VN-o | ||
| BN-c | 0.17 | 0.02 | BN-o | 0.01 | 0.02 | ||
| PP-c | 5216 | 0.23 | PP-o | 7393 | 0.64 | ||
| VN-c | 5599 | 3528 | VN-o | 7967 | 4193 | ||
Примечание. Над диагональю – уровень значимости, под диагональю – U-статистика, жирным шрифтом выделены статистически значимые значения.
Обсуждение
В аналогичных исследованиях по изучению ФА у животных влияние половых различий на уровень флуктуирующей асимметрии не выявлено, даже на фоне существенных половых различий по анализируемым признакам (Staley, Green, 1971; Zakharov, 1981; Leary et al., 1983; Pankakoski, 1985).
Ранее анализ встречаемости девиаций фолидоза у H. platyurus на окраине и в центре городов выявил, что во Вьентьяне различия между центральной и краевой зоной не наблюдаются, в Пномпене число аномалий щиткования увеличивается в центре города, в Бангкоке наблюдается обратная картина – увеличение числа аномальных вариаций на окраине (Константинов, Булдова, 2016).
В большинстве работ анализ ФА связан с индикацией неблагоприятных воздействий антропогенного характера на живой организм. В значительно меньшей степени внимание исследователей в настоящее время уделяется изучению воздействия естественных биотических факторов. В ряде публикаций по изучению ФА доказано их влияние на уровень стабильности развития. В частности, исследования, проведенные на обыкновенной бурозубке Sorex araneus, динамика численности которой подвержена четкому четырехлетнему циклу и максимально достигает 70-кратного размера (Шефтель, 1983), показали увеличение уровня флуктуирующей асимметрии у особей, родившихся в год максимальной численности, и уменьшение асимметричности у их потомков при спаде численности (Захаров и др., 1984).
Ранее было показано (Константинов, Гаврилов, 2012; Чулисов, Константинов, 2013), что в ряде городов Лаоса и Камбоджи в зависимости от степени урбанизации происходит изменение структуры сообществ, а доминирующее положение в зависимости от численности принадлежит H. platyurus и H. frenatus, причем H. platyurus является антагонистом в сообществе не только к H. frenatus, но и другим видам, вероятно, конкурируя за ресурсы среды.
Некоторые исследования на территории Флориды и на островах тропической части Тихого океана показывают, что у особей H. frenatus наблюдается как агонистическое поведение во время конфликтов между собой, так и агрессия к другим видам в сообществе, например, с Hemidactylus garnotii и Lepidodactylus lugubris (Bolger, Case, 1992; Dame, Petren, 2006).
По нашим наблюдениям, H. platyurus в городских условиях тяготеет к обитанию на деревьях, в сборах на деревьях его численность в 2.6 раза выше, чем H. frenatus. В связи с этим в районах города с меньшей озелененностью и большей освещенностью стен строений повышается вероятность как внутривидовых, так и межвидовых взаимодействий H. platyurus, что будет выражаться в обострении конкуренции.
Заключение
В ходе исследования был проведен статистический анализ ФА. Анализ направленности асимметрии признаков показал, что статистически значимые различия между величинами признака на левой и правой сторонах тела наблюдаются по нескольким признакам: количество непарных подпальцевых пластинок на 5-м пальце передней лапы (Ads5), количество подпальцевых пластинок на 1-м пальце задней лапы (Lm6), количество парных подпальцевых пластинок на 2-м пальце задней лапы (Ds7), количество щитков следующего ряда, соприкасающихся с верхнегубными (P. sl.), количество нижнегубных щитков (Il). В связи с этим данные признаки были исключены из анализа уровня ФА.
Для оценки стабильности развития H. platyurus рекомендованы следующие признаки: количество парных подпальцевых пластинок на 3-м и 5-м пальцах задней лапы (Ds8, Ds10), количество непарных подпальцевых пластинок на 3-м пальце задней лапы (ADs8), количество подпальцевых пластинок на 2, 3, 4 и 5-м пальцах передней лапы (Lm2, Lm3, Lm4, Lm5), количество подпальцевых пластинок на 2, 3 и 4-м пальцах задней лапы (Lm7, Lm8, Lm9), количество верхнегубных щитков (Sl.), количество чешуй следующего ряда, соприкасающихся с нижнегубными (G. il.), количество нижнечелюстных щитков (Im.), количество следующего ряда чешуек за нижнечелюстными (G. im.).
Половой диморфизм во всех трех анализируемых группах не выявлен (p = 0.12–0.83). Также не были выявлены возрастные различия в уровне ФА (p = 0.52–0.94), что согласуется с теорией флуктуирующей асимметрии как общебиологического явления.
Значимых различий в дифференциации биотопов по уровню ФА между центральной и краевой зонами для выборок без учета типа субстрата не выявлено ни в одной изучаемой совокупности. Проводя сравнения точек сбора на окраине, во Вьентьяне обнаружены статистически значимые различия между восточной и северной окраиной (p = 0.02).
Различия интегрального индекса в зависимости от типа субстрата выявлены во всех трех городах, причем как при сравнении выборок внутри городов, так и между собой (p < 0.05). Отмечено, что коэффициент ФА асимметрии выше на стенах (p < 0.36–0.84), чем на стволах деревьев (0.10–0.58). Обитание на стенах городских сооружений, как эволюционно молодой экологической нише, сказывается на стабильности развития вида, который в природных условиях и в городах предпочитает стволы деревьев.
При межпопуляционном сравнение выборок из Бангкок – Пномпеня, Бангкок – Вьентьян выявлены высоко значимые различия в индексе ФА (p < 0.05). При сравнении Пномпеня – Вьентьяна различий не выявлено. Бангкок – крупный транспортный и промышленно развитый мегаполис с населением более 5 млн человек, тогда как Пномпень и Вьентьян существенно уступают по этим показателям. Транспорт и промышленные предприятия являются основным источником физико-химического загрязнения городской среды, прямо или косвенно оказывающим влияние на стабильность развития живых организмов, что отмечается в целом ряде работ (Захаров, 1987; Логинов и др., 2003; Васильев и др., 2006).
Библиография
Васильев А. Г., Марин Ю. Ф., Васильева И. А. Феногенетический мониторинг березы повислой (Betula pendula): оценка качества среды в Висимском заповеднике и в зоне влияния техногенных поллютантов от предприятий цветной металлургии // Экологические исследования в Висимском биосферном заповеднике: Материалы науч. конф., посвящ. 35-летию Висимского заповедника (Екатеринбург, 2–3.10.2006 г.) / Отв. ред. Ю. Ф. Марин. Екатеринбург, 2006. С. 85–93.
Гелашвили Д. Б., Нижегородцев А. А., Епланова Г. В., Табачишин В. Г. Флуктуирующая асимметрия билатеральных признаков разноцветной ящурки Eremias arguta как популяционная характеристика // Известия Самарского научного центра Российской академии наук. 2007. Т. 9. № 4. С. 941–949.
Гелашвили Д. Б., Якимов В. Н., Логинов В. В., Епланова Г. В. Cтатистический анализ флуктуирующей асимметрии билатеральных признаков разноцветной ящурки Eremias arguta // Актуальные проблемы герпетологии и токсикологии. Вып. 7. Тольятти, 2004. C. 45–59.
Захаров В. М. Асимметрия животных (популяционно-феногенетический подход) . М.: Наука, 1987. 213 с.
Захаров В. М., Шефтель Б. И., Александров Д. Ю. Нарушение стабильности развития на фазе пика численности в популяции млекопитающих // Доклады Академии наук СССР. 1984. Т. 275. № 3. С. 761–764.
Константинов Е. Л., Гаврилов А. Б. Экологические особенности распределения некоторых видов синантропных гекконов в городах Юго-Восточной Азии (Лаос, Камбоджа) // Вопросы герпетологии: Материалы Пятого съезда герпетологического общества им. А. М. Никольского, Минск, 25–28 сентября. Минск: Право и экономика, 2012. С. 113–117.
Константинов Е. Л., Булдова О. Ю. Изменчивость фолидоза нижней челюсти Hemidactylus platyurus (SCHNEIDER, 1792) на территории Лаоса и Камбоджи // Вестник СПбГУ. 2016. Вып. 3. С. 57–62.
Логинов В. В., Гелашвили Д. Б., Чупрунов Е. В., Силкин А. А. Структурно-информационные показатели стабильности развития амфибий на антропогенно трансформированных территориях // Актуальные проблемы герпетологии и токсинологии: Сб. науч тр. Вып. 6. Тольятти, 2003. С. 85–90.
Чулисов А. С., Константинов Е. Л. Распределение синантропных гекконов в городской среде крупных городов на территории Лаоса и Камбоджи // Современная герпетология: проблемы и пути их решения: Статьи по материалам докл. Первой междунар. молодеж. конф. герпетологов России и сопредельных стран (Санкт-Петербург, Россия, 25–27 ноября 2013 г.). СПб., 2013. С. 159–162.
Шефтель Б. И. Многолетняя динамика численности землероек-бурозубок в Среднеенисейской тайге // Биологические проблемы Севера. Ч. 2. Животный мир: Тез. X Всесоюз. симпоз. Магадан, 1983. С. 57–58.
Bolger D. T., Case T. J. Intra-and interspecific interference behaviour among sexual and asexual geckos // Animal Behaviour. 1992. № 44. P. 21–30.
Dame E. A., Petren K. Behavioural mechanisms of invasion and displacement in Pacific island geckos Hemidactylus // Animal Behaviour. 2006. № 71. P. 1165–1173.
Leary R. F., Allendorf F. W., Knudsen K. L. Developmental stability and enzyme heterozygosity in rainbow trout // Nature. 1983. Vol. 301. № 5895. P. 71–72.
Mather K. Genetical control of stability in development // Heredity. 1953. Vol. 7. Pt. 3. P. 297–336.
Pankakoski E. Epigenetic asymmetry as an ecological indicator in muskrats // J. Mammal. 1985. Vol. 66. № 1. P. 52–57.
Staley R. N., Green L. J. Bilateral asymmetry in tooth cusp occurrence in human monozygotic twins, dizygotic twins, and nontwins // J. Dent. Res. 1971. Vol. 50. № 1. P. 83–89.
Waddington C. H. Organisers and genes. Cambrige: Univ. press, 1940. 160 p.
Zakharov V. M. Fluctuating asymmetry as an indez of developmental homeostasis // Genetikа (Belgrade). 1981. Vol. 13. P. 241–256.

 © 2011 - 2026
© 2011 - 2026