



Введение
Высшая водная растительность оказывает существенное влияние на видовую структуру и обилие зоопланктона (Семенченко, Разлуцкий, 2009; Семенченко и др., 2013; Курбатова и др., 2018). Зоопланктоценозы зарослей макрофитов характеризуются высоким видовым богатством и количественным развитием (Крылов, 2005; Лобуничева, 2008). Видовая структура сообществ зоопланктона в прибрежной зоне водоемов определяется, главным образом, интенсивностью развития макрофитов (Kuczynska-Kippen, 2006), а также их морфологическим строением (Семенченко, Разлуцкий, 2009; Jeong et al., 2014). Погруженные водные растения с большей биомассой образуют высокую гетерогенность местообитаний, что вызывает увеличение разнообразия и плотности зоопланктона (Spoljar et al., 2012; Choi et al., 2014). В то же время некоторые макрофиты (кубышка желтая, пузырчатка) оказывают угнетающее действие на зоопланктон (Зимбалевская и др., 1987; Курбатова и др., 2012; Зайцева и др., 2014). Кроме того, формирующиеся в разнотипных по морфологическому строению зарослях макрофитов специфические физико-химические условия и трофические взаимоотношения обуславливают различия видовой структуры сообществ зоопланктона (Курбатова и др., 2018).
К настоящему времени зоопланктон зарослей высших водных растений равнинных рек изучен крайне слабо. В литературе имеются сведения, что зоопланктон зарослей прибрежно-водных макрофитов рек количественно более развит по сравнению с незаросшей медиалью (Крылов, 2005). Кроме того, наибольшее количественное развитие зоопланктона зарослей макрофитов в пределах зоны подпора речных вод отмечено для устьевой области малой реки-притока равнинного водохранилища (Столбунова, 2011). В зонах слияния вод незарегулированных рек структура зоопланктона зависит от скорости течения и степени зарастания высшей водной растительностью (Болотов и др., 2012). В целом высшая водная растительность оказывает существенное влияние на пространственное размещение сообществ зоопланктона на различающихся по морфометрическим характеристикам участках равнинных рек (Гаврилко и др., 2018).
Особого внимания заслуживает исследование зоопланктоценозов участков слияния рек и озер, где формируются переходные зоны, в которых снижается скорость течения реки, что благоприятно влияет на развитие высшей водной растительности в рипали. Участки слияния разнотипных водных объектов определяют структурно-функциональную организацию сообществ гидробионтов объектов-реципиентов (Болотов и др., 2012). В отечественной литературе практически отсутствуют сведения о видовой структуре сообществ зоопланктона разнотипных макрофитов в зоне контакта речных и озерных вод. Одной из таких зон является участок впадения р. Сережа в озеро Великое, прибрежье которого занято разнотипными растительными ассоциациями.
Река Сережа – правый приток второго порядка р. Ока. Длина реки составляет 196 км, площадь бассейна – 2730 км2. Река характеризуется извилистым, слаборазветвленным песчаным с каменистыми перекатами руслом. Пойма реки покрыта лесом, кустарниками, местами заболочена. В среднем течении формирует систему карстовых озер (Великое, Глубокое, Паровое и др.) (Природа…, 1974).
Сведения о зоопланктоне р. Сережа представлены в ряде работ (Шурганова и др., 2012; Ильин и др., 2015, 2016; Ильин, 2016). Однако изучение зоопланктона зарослей высшей водной растительности реки было проведено впервые.
Цель настоящей работы – характеристика видовой структуры и пространственного распределения сообществ зоопланктона медиали и зарослей высшей водной растительности на акватории р. Сережа в месте впадения в озеро Великое.
Материалы
Исследования зоопланктона были проведены в среднем течении р. Сережа (Арзамасский р-он, Нижегородская область) в месте впадения в озеро Великое (N 55.658543, E 43.596692) в середине июля 2016 г. Отбор проб проводился на участке длиной 1.3 км до впадения в озеро, а также в оз. Великое, на участке, после впадения р. Сережа (рис. 1).
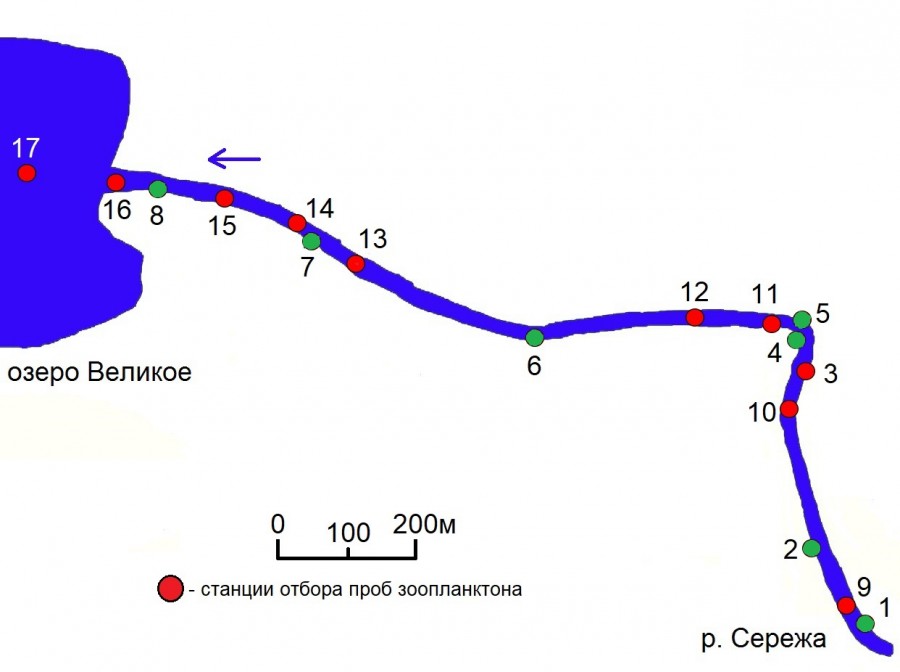
Рис. 1. Схема расположения станций отбора проб зоопланктона р. Сережа в зоне впадения в озеро Великое. Зеленым цветом отмечены станции в зарослях макрофитов, красным – в открытой воде. 1 – биотоп кубышки желтой; 2 – биотоп хвоща приречного; 3 – открытое прибрежье; 4 – биотоп элодеи канадской; 5 – биотоп телореза обыкновенного; 6 – биотоп стрелолиста обыкновенного; 7 – биотоп рдеста блестящего; 8 – биотоп рдеста плавающего; 9–12 – медиаль речного участка; 13–16 – медиаль зоны подпора, 17 – озеро Великое
Fig. 1. Sceme of location of zooplankton sampling stations on the river Serezha in the zone of its influx into lake Velikoye. The stations in the macrophyte thickets are marked in green, in open water - in red. 1 – biotop of Nuphar lutea; 2 – biotop of Equisetum fluviatile; 3 – open coastal area; 4 – biotop of Elodea canadensis; 5 – biotop of Stratiotes aloides; 6 – biotop of Sagittaria sagittifolia; 7 – biotop of Potamogeton lucens; 8 – biotop of Potamogeton natans; 9–12 – medial of the river area; 13–16 – medial of the backwater zone, 17 – lake Velikoye
Были выбраны заросли макрофитов разного морфологического строения: кубышка желтая Nuphar lutea (L.) Sm., хвощ приречный Equisetum fluviatile L., Sm., элодея канадская Elodea canadensis Michx., телорез обыкновенный Stratiotes aloides L., стрелолист обыкновенный Sagittaria sagittifolia L., рдест блестящий Potamogeton lucens L. Все эти биотопы расположены в рипали реки, медиальная часть которой характеризовалась значительной скоростью течения (0.3–0.35 м/с). В прибрежье реки в зоне подпора озера располагался биотоп рдеста плавающего Potamogeton natans (L.).
Методы
Пробы зоопланктона в зарослях высших водных растений отбирали мерным ведром, путем процеживания 50 литров воды через планктонную сеть (нейлоновое сито с ячеей 70 мкм) и фиксировали 4 % раствором формалина. В медиальной зоне реки и в озере пробы отбирали сетью Джеди, путем процеживания столба воды от дна до поверхности. В каждом зарослевом биотопе было взято по 3 пробы зоопланктона. Обработка материала осуществлялась общепринятыми методами (Методические рекомендации…, 1982). Идентификация видов зоопланктона осуществлялась с использованием определителей (Кутикова, 1970; Определитель…, 2010).
Параллельно с отбором проб проводили измерения глубины, прозрачности, температуры, рН, электропроводности воды, концентрации растворенного кислорода, скорости течения, процента проективного покрытия растениями биотопа.
Оценку сходства видового состава зоопланктона проводили на основе коэффициента Сёренсена (Шитиков и др., 2003). Доминирующие виды зоопланктона выявляли согласно индексу доминирования Палия – Ковнацки (Шитиков и др., 2003). Пробы зоопланктона классифицировали с помощью кластерного анализа на основе сходства видовой структуры, в качестве меры сходства использовали косинус угла между векторами проб в многомерном пространстве численностей видов (Шурганова и др., 2003; Шурганова, 2007). Кластерный анализ проводили на основе расчетов расстояния между группами проб (кластерами) методом средней связи. Для выбора оптимального числа кластеров при кластеризации проб зоопланктона проводили анализ силуэтов и анализ коэффициентов корреляции Мантеля (Legendre & Legendre, 2012; Якимов и др., 2016). Ординацию данных проводили с применением анализа избыточности (RDA) (Legendre & Legendre, 2012; Шитиков, Розенберг, 2013). Все расчеты проводились в среде R (R Core Team, 2015).
Исследуемый участок реки характеризовался нейтральной и слабощелочной реакцией среды (рН 7.23–8.28). Электропроводность изменялась в пределах 165.7–219.0 мкСм/см. Во всех биотопах наблюдались высокие концентрации растворенного кислорода (9.54–12.58 мг/л), за исключением зарослей телореза и хвоща (7.51–7.85 мг/л). Для всех растительных ассоциаций было характерно доминирование одного вида растения, который занимал более 60 % проективного покрытия в биотопе.
Результаты
В зоопланктоне исследованного участка р. Сережа было идентифицировано 120 видов, из них 65 принадлежало коловраткам (Rotifera), 39 – ветвистоусым ракообразным (Cladocera), 16 – веслоногим ракообразным (Copepoda). Среди идентифицированных видов была найдена коловратка Kellicottia bostoniensis (Rousselet, 1908) – вид-вселенец североамериканского происхождения. Коловратка была отмечена только на медиальных станциях, а также открытого прибрежья и не встречалась в зарослевых биотопах.
На основе коэффициента сходства видового состава Сёренсена была построена кластерная дендрограмма (рис. 2). Станции отбора проб зоопланктона сгруппировались в два четко обособленных кластера. В первый кластер вошли станции из прибрежных биотопов (1–8), при этом наиболее далеко от зарослевых проб разместилась станция № 3, расположенная в открытом прибрежье. Второй кластер сформировали станции медиальной зоны реки (9–16) и озера (17).
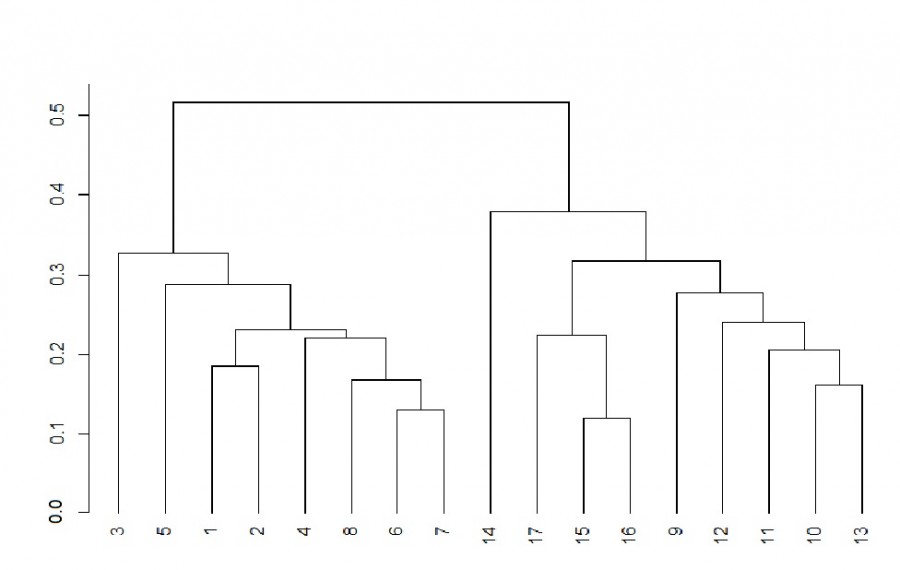
Рис. 2. Дендрограмма иерархической кластеризации проб зоопланктона на основе коэффициента Сёренсена. По оси отложено расстояние объединения. Описание станций отбора проб как на рис. 1
Fig. 2. Dendrogram of hierarchical clustering of zooplankton samples based on the Sörensen coefficient. Axis represents the joining distance. Description of sampling stations as in Fig. 1
Результаты анализа дендрограммы (см. рис. 2) свидетельствуют о различиях видового состава зоопланктона медиальной и прибрежной зон реки. Наибольшее количество видов зоопланктона среди всех прибрежных биотопов было характерно для зарослей телореза (80 видов) (табл. 1), а наименьшее – для биотопа открытого прибрежья (45 видов). При этом в медиальной зоне реки количество видов составило 42, что было сравнительно ниже, чем в зарослевых биотопах. Значительная разветвленность и большая плотность телореза способствовали формированию высокого видового богатства зоопланктона.
Таблица 1. Показатели видовой структуры и количественного развития сообществ зоопланктона
| Станция | Биотоп | Доминанты | D | S | N, тыс. экз./м3 | N, % Rot:Cla:Cop | B, г/м3 | B, % Rot:Cla:Cop |
| 1 | Кубышка желтая | Conochiloides coenobasis
Keratella cochlearis Hexarthra mira |
33.3 16.6 15.8 | 61 | 755.7± 183.9 | 84:5:11 | 1.31±0.23 | 54:9:37 |
| 2 | Хвощ приречный | Copepodit Cyclopoida
Ceriodaphnia pulchella Thermocyclops oithonoides Conochiloides coenobasis |
17.7 13.6 13.4 10.3 | 65 | 579.7± 299.7 | 23:34:43 | 7.31±5.31 | 2:44:54 |
| 3 | Открытое прибрежье | Nauplii Copepoda
Conochiloides coenobasis Hexarthra mira |
20.6 17.2 13.5 | 45 | 133.9± 38.9 | 63:10:27 | 0.23±0.02 | 38:20:42 |
| 4 | Элодея канадская | Ceriodaphnia pulchella
Ceriodaphnia quadrangula Thermocyclops crassus Diaphanosoma brachyurum |
26.3 17.1 11.1 10.1 | 61 | 872.4± 89.8 | 1:64:35 | 22.82± 5.96 | 1:79:20 |
| 5 | Телорез обыкновенный | Conochilus unicornis
Copepodit Cyclopoida Nauplii Copepoda |
29.5 25.1 16.3 | 80 | 343.2± 21.3 | 36:20:44 | 2.31±0.22 | 3:62:35 |
| 6 | Стрелолист обыкновенный | Diaphanosoma brachyurum
Ceriodaphnia pulchella Copepodit Cyclopoida |
26.2 15.1 12.6 | 65 | 682.2± 327.8 | 5:58:37 | 13.55± 2.03 | 1:78:21 |
| 7 | Рдест блестящий | Nauplii Copepoda
Copepodit Cyclopoida Conochiloides coenobasis |
22.4 20.3 12.8 | 62 | 111.7± 25.3 | 34:22:44 | 1.49±0.67 | 4:84:12 |
| 8 | Рдест плавающий | Nauplii Copepoda
Polyarthra euryptera |
23.7 13.5 | 73 | 534.2± 92.3 | 53:13:34 | 2.02±0.31 | 29:40:31 |
| 9–12 | Медиаль, свободное течение реки | Nauplii Copepoda
Conochiloides coenobasis Copepodit Cyclopoida Keratella cochlearis |
19.4 17.0 16.2 10.3 | 42 | 179.1± 114.4 | 56:7:37 | 0.74±0.41 | 43:12:45 |
| 13–16 | Медиаль, зона подпора | Nauplii Copepoda
Bosmina longirostris Brachionus angularis Copepodit Cyclopoida |
41.8 14.2 11.8 11.5 | 41 | 728.2± 157.8 | 32:15:53 | 2.18±0.83 | 31:18:51 |
| 17 | Озеро, после впадения реки | Asplanchna priodonta
Copepodit Cyclopoida |
20.8 14.9 | 29 | 278.8 | 58:12:30 | 3.55 | 36:25:39 |
Примечание. D – индекс доминирования, S – общее число видов, N – общая численность, B – общая биомасса, Rot:Cla:Cop – соотношение коловраток, ветвистоусых и веслоногих ракообразных.
По результатам анализа видовой структуры сообществ зоопланктона была построена дендрограмма иерархической кластеризации (рис. 3). На основе анализа ширины силуэта и коэффициента корреляции Мантеля было принято разбиение дендрограммы на пять кластеров. Станции № 9–12, расположенные в медиальной зоне реки с выраженной скоростью течения, характеризовались высоким сходством видовой структуры, что позволило им сгруппироваться в один кластер. В этом сообществе зоопланктона доминировали науплиальные и копеподитные стадии (СI–CV) веслоногих ракообразных, коловратки Conochiloides coenobasis Skorikov, 1914 и Keratella cochlearis (Gosse, 1851) (табл. 1). Средняя численность зоопланктона составила 179.1 ± 114.4 тыс. экз./м3, а биомасса – 0.74±0.41 г/м3.
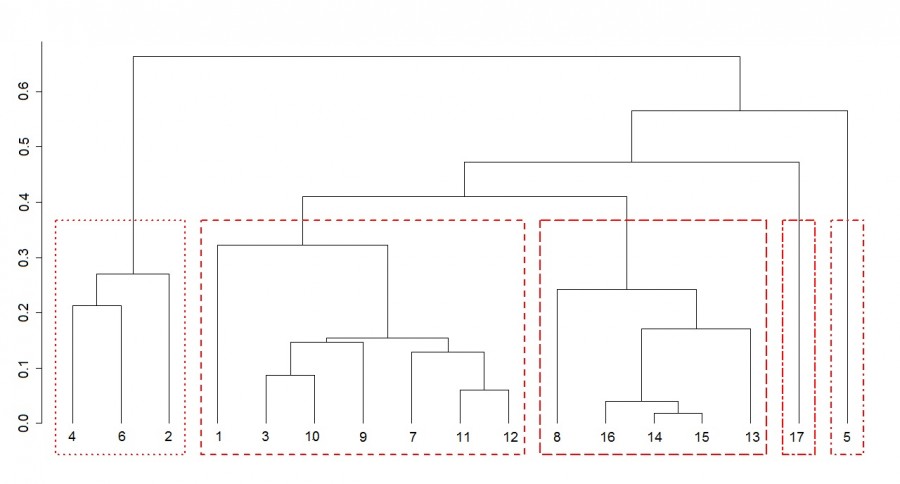
Рис. 3. Дендрограмма иерархической кластеризации проб зоопланктона на основе видовой структуры. По оси отложено расстояние объединения. Одинаковым типом линии выделены соответствующие кластеры. Описание станций отбора проб как на рис. 1
Fig. 3. Dendrogram of hierarchical clustering of zooplankton samples based on species structure. Axis shows thejJoining distance. The corresponding clusters are selected with the same line type. Description of sampling stations as in Fig. 1
Станции № 13–16, расположенные в зоне подпора озера, обладали сходной видовой структурой зоопланктона (см. рис. 3), что свидетельствует о формировании переходного зоопланктоценоза на этом участке. Здесь доминантами выступали младшие возрастные стадии веслоногих ракообразных, а также ветвистоусый рачок Bosmina longirostris (O. F. Müller, 1785) и коловратка Brachionus angularis Gosse, 1851 (см. табл. 1). Численность и биомасса зоопланктона переходного ценоза были выше, чем на речном участке (728.2 ± 157.8 тыс. экз./м3 и 2.18 ± 0.83 г/м3). Обособленно на дендрограмме расположилась станция № 17 озера Великое (см. рис. 3). Видовая структура зоопланктона этого участка существенно отличалась от речных сообществ. На этой станции доминировали коловратка Asplanchna priodonta и копеподитные стадии (CI–CV) веслоногих рачков. Численность зоопланктона составила 278.8 тыс. экз./м3, биомасса – 3.55 г/м3.
Видовая структура сообществ зоопланктона прибрежных биотопов (станции № 1–8) значительно различалась между собой. Наиболее обособленно на дендрограмме расположены биотопы погруженных макрофитов с большим проективным покрытием (элодея, телорез) и биотопы воздушно-водных растений (хвощ и стрелолист) (см. рис. 3). Зоопланктоценозы зарослей растений с плавающими листьями более тяготели к зонам медиали. Так, заросли кубышки желтой (ст. 1) были расположены в прибрежье свободного течения реки. В этом зоопланктоценозе доминировали коловратки, характерные для зон открытой воды, – Conochiloides coenobasis, Keratella cochlearis и Hexarthra mira (Hudson, 1871). Однако общая численность и биомасса зоопланктона были значительно выше, чем в медиальной зоне (см. табл. 1). Заросли рдеста плавающего (ст. 8) были расположены в прибрежье зоны контакта речных и озерных вод. Здесь преобладали науплиальные стадии веслоногих рачков и коловратка Polyarthra euryptera (Wierzejski, 1891), предпочитающая озерные и прудовые условия. Биотоп зарослей рдеста блестящего (ст. 7) попал в один кластер с пробами из медиальной зоны реки (см. рис. 3). Низкое проективное покрытие растений наряду со значительной скоростью течения способствовало формированию сходной видовой структуры зоопланктоценозов этих зарослей и зоны медиали. В данный кластер также вошел биотоп открытого прибрежья (ст. 3).
Зоопланктоценоз биотопа телореза (ст. 5) значительно отличался от других зарослевых сообществ присутствием в числе доминантов колониальной коловратки Conochilus unicornis Rousselet, 1892. При этом численность и биомасса зоопланктона здесь были сравнительно невысокими (см. табл. 1).
В сообществе зоопланктона зарослей элодеи (ст. 4) доминировали ветвистоусые ракообразные Ceriodaphnia pulchella Sars, 1862, Ceriodaphnia quadrangula (O. F. Müller, 1785), Diaphanosoma brachyurum (Liévin, 1848), а также веслоногий рачок Thermocyclops crassus (Fischer, 1853). Для этого сообщества были отмечены максимальные значения численности (872.4 ± 89.8 тыс. экз./м3) и биомассы (22.82 ± 0.96 г/м3) зоопланктона среди всех биотопов реки. Для зоопланктоценоза зарослей стрелолиста (ст. 6) также было характерно преобладание ветвистоусых рачков Diaphanosoma brachyurum и Ceriodaphnia pulchella. Это позволило ему попасть в один кластер с зоопланктоценозом зарослей элодеи (см. рис. 3). Здесь были отмечены также высокие значения численности и биомассы зоопланктона (682.2 ± 327.8 тыс. экз./м3 и 13.55 ± 2.03 г/м3 соответственно).
Состав доминантов зоопланктоценоза зарослей хвоща (ст. 2) отличался от зарослей элодеи и стрелолиста присутствием рачка Thermocyclops oithonoides (Sars, 1863) и коловратки Conochiloides coenobasis. Однако доминирование Ceriodaphnia pulchella и копеподитных стадий (CI–CV) веслоногих рачков обусловило сходство видовой структуры зоопланктона этих зарослей макрофитов и формирование единого кластера (см. рис. 3). Количественное развитие зоопланктона зарослей хвоща было ниже, чем в зарослях элодеи и стрелолиста (см. табл. 1).
Соотношение численностей таксономических групп зоопланктона исследованных зарослей макрофитов существенно различалось. Для растений с плавающими листьями (кубышка желтая, рдест плавающий) было характерно преобладание коловраток в общей численности зоопланктона (см. табл. 1). В зарослях элодеи и стрелолиста доминировали ветвистоусые ракообразные. Для остальных биотопов было свойственно преобладание веслоногих ракообразных.
В отличие от численности зоопланктона, большую часть биомассы большинства зарослевых сообществ составляли ветвистоусые ракообразные. Исключение – биотоп кубышки, где наблюдалось преобладание коловраток, а также биотоп хвоща, где на первый план выходили веслоногие ракообразные (см. табл. 1).
Для оценки зависимости видовой структуры сообществ зоопланктона от факторов окружающей среды была построена модель на основе анализа избыточности (RDA) с учетом следующих абиотических и биотических показателей.
Статистический анализ вспомогательных моделей, которые были построены для каждого отдельного фактора, показал, что значимым влиянием p < 0.05 обладали все факторы, кроме концентрации растворенного кислорода (табл. 2). Полная модель RDA объясняла 40.94 % (р < 0.001) от общей дисперсии видовой структуры сообществ зоопланктона. Статистически значимыми в модели были первые две оси (табл. 3).
Таблица 2. Статистический анализ значимости объяснения изменчивости видовой структуры для каждого отдельного фактора
| Фактор | Скорректированная доля объясняемой дисперсии, % | Значение критерия Фишера, F | p |
| Проективное покрытие растений | 21.0 | 5.26 | 0.001 |
| Температура | 14.1 | 3.63 | 0.002 |
| Скорость течения | 13.1 | 3.41 | 0.007 |
| Электропроводность | 13.5 | 3.49 | 0.012 |
| рН | 12.2 | 3.22 | 0.007 |
| Прозрачность | 10.7 | 2.93 | 0.014 |
| Глубина | 6.7 | 2.15 | 0.046 |
| Концентрация растворенного кислорода | 1.3 | 1.21 | 0.276 |
Таблица 3. Статистический анализ данных модели на основе анализа избыточности
| Модель, ось | Скорректированная доля объясняемой дисперсии, % | Значение критерия Фишера, F | p |
| Полная модель | 40.94 | 2.58 | 0.001 |
| Ось I | 19.18 | 8.47 | 0.002 |
| Ось II | 10.75 | 4.75 | 0.003 |
| Ось III | 3.94 | 1.74 | 0.138 |
| Ось IV | 3.12 | 1.38 | 0.224 |
Расположение станций на ординационной диаграмме, построенной по результатам анализа избыточности, согласуется с результатами кластерного анализа и свидетельствует о различающейся видовой структуре сообществ зоопланктона зарослей макрофитов (рис. 4). Станции, соответствующие медиальным и зарослевым сообществам, располагаются вдоль первой (горизонтальной) оси ординации, значимо объясняющей 19.18 % общей дисперсии численности зоопланктона (см. табл. 3). Положение станций на диаграмме вдоль второй (вертикальной) оси, объясняющей 10.75 % дисперсии (см. табл. 3), связано с влиянием абиотических факторов. Станции медиали речного участка реки (9–12) расположены вдоль коррелирующих между собой векторов скорости течения и прозрачности (см. рис. 4). Напротив, станции, находящиеся в медиали переходного участка реки (13–16), а также ниже впадения реки (17), разместились вдоль вектора рН. В сторону увеличения рН расположилась и станция № 8 зарослей рдеста плавающего, находящаяся в прибрежье переходного участка реки. Большинство других зарослевых станций разместились вдоль вектора проективного покрытия растений (см. рис. 4).
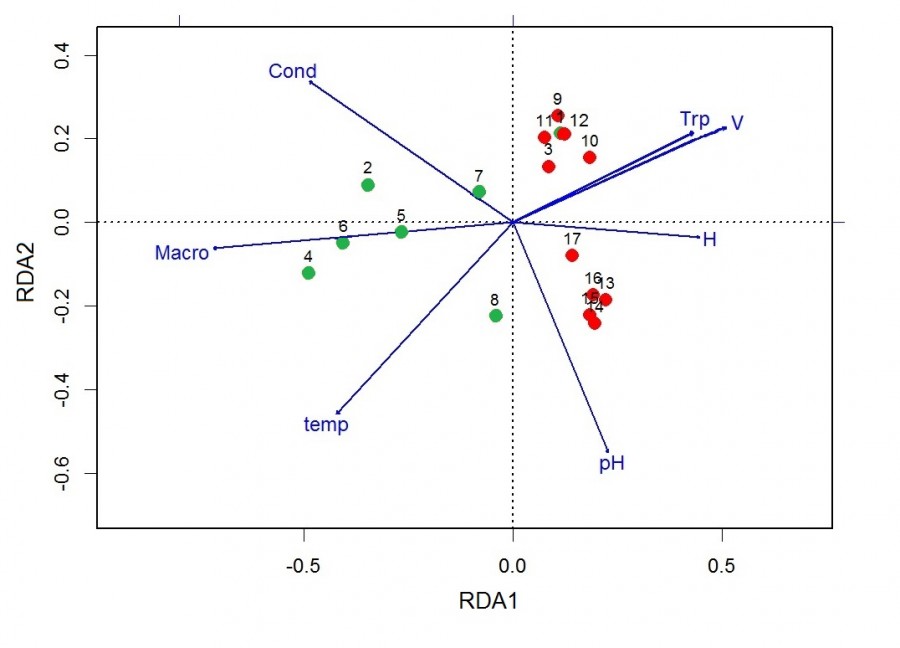
Рис. 4. Ординационная диаграмма, построенная по результатам анализа избыточности (RDA) для проб зоопланктона, отобранных на акватории р. Сережа: 1–17 – номера станций, H – глубина, temp – температура, Cond – электропроводность, Trp – прозрачность, pH – водородный показатель, V – скорость течения, Macro – проективное покрытие растений. Описание станций как на рис. 1
Fig. 4. Ordination dendrogramm plotted on the basis of redundancy analysis for zooplankton sampling, selected in the waters of the river Serezha. 1–17 – number of station, H – depth, temp – temperature, Cond – conductivity, Trp – transparency, pH – acidity, V – flow rate, Macro – plant cover. Description of stations as in Fig.1.
Обсуждение
Анализ полученных результатов позволяет выделить в медиали р. Сережа участки, занятые различающимися по видовой структуре и показателям количественного развития сообществами зоопланктона. Зоопланктоценоз речного участка с выраженной скоростью течения характеризовался невысоким количественным развитием зоопланктона и численным преобладанием коловраток. Для сообщества зоопланктона переходного участка реки с замедленным течением было характерно возрастание количественного развития зоопланктона по сравнению с речным участком. Здесь по численности и биомассе зоопланктона преобладали веслоногие ракообразные. Выделение переходного зоопланктоценоза р. Сережа в месте впадения в озеро Великое хорошо согласуется с результатами предыдущих исследований (Ильин и др., 2015; Ильин, 2016).
Видовая структура сообществ зоопланктона зарослей высшей водной растительности существенно различалась между разными биотопами и зоной медиали. Для зарослевых сообществ было характерно наибольшее видовое богатство зоопланктона в реке. В наиболее плотно сомкнутых зарослях макрофитов (элодеи канадской и стрелолиста обыкновенного) с высоким проективным покрытием (85–90 %) наблюдалось наибольшее количественное развитие зоопланктона. Большой объем погруженной части макрофита формирует высокую гетерогенность водной среды, которая является ключевым фактором массового развития зоопланктона в зарослях. Ряд авторов также отмечают, что наибольшего развития в погруженных макрофитах достигают именно ветвистоусые ракообразные (Stefanidis, Parastergiadou, 2010; Bolduc et al., 2016; Курбатова и др., 2017), которые обеспечивают высокие значения биомассы зоопланктона.
Заросли растений с плавающими листьями (кубышка желтая, рдест плавающий) создают низкую гетерогенность водной среды, что приводит к близости расположения этих станций на дендрограмме (см. рис. 3) со станциями медиали реки. Для этих зоопланктоценозов было характерно преобладание коловраток в общей численности зоопланктона. В работе О. В. Мухортовой с соавторами (Mukchortova et al., 2015) также отмечается сходство зоопланктона зарослей кубышки желтой с зоной открытой воды.
В густых зарослях элодеи и стрелолиста по численности зоопланктона доминировали ветвистоусые ракообразные. Основу биомассы большинства зарослевых сообществ зоопланктона формировали ветвистоусые ракообразные.
Основным фактором, определяющим изменчивость видовой структуры сообществ зоопланктона, являлось проективное покрытие растений. Близость расположения зарослевых станций к вектору проективного покрытия на дендрограмме (см. рис. 3) определялась густотой зарослей. Среди абиотических факторов особого внимания заслуживают скорость течения и рН воды. Для речного участка р. Сережа характерны значительные скорости течения (0.3 м/с) и нейтральная реакция среды (7.49–7.55). С продвижением к озеру в переходном участке реки скорость течения падала до 0.1 м/с, а рН воды возрастала до 8.15. Это закономерно отражалось на видовой структуре сообществ зоопланктона и расположении станций на ординационной дендрограмме (см. рис. 4).
Таким образом, заросли высшей водной растительности являются ведущим фактором, обуславливающим видовую структуру сообществ зоопланктона. Они образуют зоны рефугиумов в реке, формирующих благоприятные условия для массового развития зоопланктона и позволяющих обогащать планктонную фауну реки. Если в медиали реки формирование планктона определяется преимущественно скоростью течения, то в рипали главную роль играет высшая водная растительность (архитектоника макрофита и густота зарослей).
Заключение
На исследованной акватории зоны слияния реки Сережи и озера Великого выделены сообщества зоопланктона с различающейся видовой структурой, расположенные в медиали реки, – сообщество речного участка и сообщество переходного участка.
Установлено, что видовая структура сообществ зоопланктона разнотипных зарослей макрофитов существенно отличалась между зарослевыми биотопами и зоной медиали.
Выявлено, что наибольшее видовое богатство зоопланктона в реке сосредоточено в прибрежных зарослях высшей водной растительности. Наибольшее количественное развитие сообществ зоопланктона характерно для наиболее густых зарослей макрофитов (элодеи канадской и стрелолиста обыкновенного).
Анализ влияния факторов среды на видовую структуру зоопланктоценозов показал, что наибольшую часть общей дисперсии объяснял такой параметр, как проективное покрытие растений.
Библиография
Болотов С. Э., Цветков А. И., Крылов А. В. Зоопланктон зон слияния незарегулированных рек // Биология внутренних вод. 2012. № 2. С. 29–36.
Гаврилко Д. Е., Кудрин И. А., Ручкин Д. С., Шурганова Г. В. Влияние высших водных растений на структуру сообществ зоопланктона малой реки (на примере реки Вьюница г. Нижнего Новгорода) // Материалы III международной конференции «Актуальные проблемы планктонологии». Калининград: АтлантНИРО, 2018. С. 47–50.
Зайцева В. Л., Филиппов Д. А., Лобуничева Е. В., Михайлова А. А. Влияние Urticularia intermedia на структуру сообществ водных беспозвоночных болотных водоемов // Известия Самарского научного центра РАН. 2014. Т. 16. № 5. С. 276–281.
Зимбалевская Л. Н., Плигин Ю. В., Хороших Л. А., Долинский В. Л., Сидоренко В. М., Левина О. В., Георгиевская Л. М., Ревенко И. Г., Гошовская Г. А., Козина С. Я., Емельянов Л. В., Левитская Н. А. Структура и сукцессии литоральных биоценозов днепровских водохранилищ . Киев: Наук. думка, 1987. 204 с.
Ильин М. Ю., Шурганова Г. В., Куклина Т. В., Кудрин И. А. Пространственное размещение сообществ зоопланктона в зоне контакта речных и озерных вод (на примере реки Сережи и озера Великого Нижегородского Поволжья) // Современные проблемы науки и образования. 2015. № 6. URL: www.science–education.ru/130–23340.
Ильин М. Ю. Состав и структура сообществ зоопланктона водных объектов особо охраняемых природных территорий (на примере Нижегородской области) : автореф. дис. ... канд. биол. наук. Н. Новгород, 2016. 27 с.
Ильин М. Ю., Кудрин И. А., Золотарева (Куклина) Т. В., Шурганова Г. В. Биоиндикация водных объектов особо охраняемых природных территорий Нижегородской области на основе анализа видовой структуры зоопланктона // Вода: химия и экология. 2016. № 3. С. 25–33.
Крылов А. В. Зоопланктон равнинных малых рек . М.: Наука, 2005. 263 с.
Курбатова С. А.. Лаптева Н. А., Ершов И. Ю., Борисовская Е. В. Фоновые характеристики среды и динамика планктонных сообществ в экосистемах с гидрофитами // Поволжский экологический журнал. 2012. № 1. С. 42–52.
Курбатова С. А., Ершов И. Ю., Борисовская Е. В. Влияние плотности зарослей гидрофитов на зоопланктон // Биология внутренних вод. 2017. № 1. С. 84–92
Курбатова С. А., Мыльникова З. М., Ершов И. Ю., Быкова С. Н., Виноградова О. Г. Влияние водных растений разных экологических групп на распределение и обилие зоопланктона // Сибирский экологический журнал. № 1. 2018. С. 56–66.
Кутикова Л. А. Коловратки фауны СССР . Л.: Наука, 1970. 742 с.
Лобуничева Е. В. Зарослевый зоопланктон некоторых малых озер Вологодской области // Водные экосистемы: трофические уровни и проблемы поддержания биоразнообразия: Материалы Всерос. конф. с междунар. участием «Водные и наземные экосистемы: проблемы и перспективы исследований» (Вологда, Россия, 24–28 ноября 2008 г.). Вологда, 2008. С. 188–192.
Методические рекомендации по сбору и обработке материалов при гидробиологических исследованиях на пресноводных водоемах // Зоопланктон и его продукция. Л.: Гос. НИИ озер. и реч. рыб. хоз-ва, 1982. 33 с.
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 1. Зоопланктон / Под ред. В. Р. Алексеева, С. Я. Цалолихина. М.: Товарищество научных изданий КМК, 2010. 495 с.
Природа Горьковской области / Под. ред. Н. В. Кузнецова. Горький: Волго-Вятское кн. изд-во, 1974. 416 с.
Семенченко В. П., Разлуцкий В. И. Факторы, определяющие суточное распределение и перемещения зоопланктона в литоральной зоне пресноводных озер // Журнал Сибирского федерального университетата. Сер. Биология. 2009. № 2. С. 191–225.
Семенченко В. П., Разлуцкий В. И., Бусева Ж. П., Палаш А. Л. Зоопланктон литоральной зоны озер разного типа . Минск: Беларус. навука, 2013. 181 с.
Столбунова В. Н. Зоопланктон зарослей макрофитов в устьевой области малой реки // Биология внутренних вод. 2011. № 2. С. 35–42.
Шитиков В. К., Розенберг Г. С., Зинченко Т. Д. Количественная гидроэкология: методы системной идентификации . Тольятти: ИЭВБ РАН, 2003. 463 с.
Шитиков В. К., Розенберг Г. С. Рандомизация и бутстреп: статистический анализ в биологии и экологии с использованием R . Тольятти: Кассандра, 2013. 314 с.
Шурганова Г. В., Черепенников В. В., Артельный Е. В. Динамика пространственного распределения основных зоопланктоценозов Чебоксарского водохранилища // Поволжский экологический журнал. 2003. № 3. С. 297–304.
Шурганова Г. В. Динамика видовой структуры зоопланктоценозов в процессе их формирования и развития (на примере водохранилищ средней Волги: Горьковского и Чебоксарского) : Автореф. дис. … д-ра биол. наук. Н. Новгород, 2007. 48 с.
Шурганова Г. В., Черепенников В. В., Тарбеев М. Л., Маслова Г. О. Видовая структура зоопланктона реки Сережа Нижегородской области // Вестник Нижегородского университета им. Н. И. Лобачевского. 2012. № 3 (1). С. 111–117.
Якимов В. Н., Шурганова Г. В., Черепенников В. В., Кудрин И. А., Ильин М. Ю. Методы сравнительной оценки результатов кластерного анализа структуры гидробиоценозов (на примере зоопланктона реки Линда Нижегородской области) // Биология внутренних вод. 2016. № 2. С. 94–103.
Bolduc P., Bertolo А., Pinel-Alloul В. Does submerged aquatic vegetation shape zooplankton community structure and functional diversity? A test with a shallow fluvial lake system // Hydrobiologia. 2016. № 778. Р. 151–165.
Choi J. Y., Jeong K. S., La G. H., Joo G. J. Effect of removal of free-floating macrophytes on zooplankton habitat in shallow wetland // Knowledge and Management of Aquatic Ecosystems. 2014. Vol. 414 (11).
Jeong K. S., Choi J. Y., Jeong Kw. S. Influence of aquatic macrophytes on the interactions among aquatic organisms in shallow wetlands (Upo Wetland, South Korea) // J. Ecol. Environ. 2014. Vol. 37 (4). P. 185–194.
Kuczynska-Kippen N. Zooplankton structure in architecturally differentiated macrophyte habitats of shallow lakes in the Wielkopolska Region, Poland // International Journal of Oceanography and Hydrobiology. 2006. Vol 35. № 2. P. 179–191.
Legendre P., Legendre L. Numerical ecology. Oxford: Elsevier, 2012. 990 p.
Mukchortova O., Bykova S., Tarasova N., Unkovskaya E., Bolotov S. Plankton Community in the Pelagic and Littoral Zones of the Overgrown Lake Beloe (Volzhsko-Kamskiy Biosphere Natural State Reserve, Republic of Tatarstan, Russian Federation) // Journal of Siberian Federal University. Biology 1. 2015 (8). P. 66–84.
R Core Team. R: A language and environment for statistical computing / R Core Team. 2015. URL: http://www.R–project.org.
Spoljar M., Drazina T., Sargac J., Kralj Borojevic K., Zutinic P. Submerged macrophytes as a habitat for zooplankton development in two reservoirs of a flow-through system (Papuk Nature Park, Croatia) // Ann. Limnol. Int. J. Lim. 2012. Vol. 48. P. 161–175.
Stefanidis K., Parastergiadou E. Influence of hydrophyte abundance on the spatial distribution of zooplankton in selected lakes in Greece // Hydrobiology. 2010. № 656. P. 55–65.
Благодарности
Работа выполнена при финансовой поддержке РФФИ (грант 18-04-00673 А).

 © 2011 - 2026
© 2011 - 2026