



Введение
Орнитологическая изученность Приморского края по отношению к большинству регионов бывшего СССР сравнительно высока (15-е место из 89) (Исаков, 1982), хотя в основном здесь выполняли фаунистические исследования и работы по отдельным видам птиц. Количественные характеристики орнитокомплексов и их пространственной неоднородности до настоящего времени явно недостаточны, хотя работы начаты почти полвека назад (Назаренко, 1968).
За последние годы накоплены и опубликованы хотя и разнородные материалы, но позволяющие характеризовать население птиц значительной части растительных формаций и их антропогенных сукцессий на территории Приморья (Назаренко, 1968, 1971, 1984; Кулешова, 1976; Кушнарев, 1982, 1984; Симонов, 1985, 1987; Елсуков, 1987). Целью исследования, результаты которого изложены в предлагаемой вниманию читателей статье, является обобщение накопленных данных по летнему населению птиц Приморья и выявление пространственной структуры изменчивости орнитокомплексов. Актуальность работы состоит в необходимости качественной характеристики птиц для оценки воздействия естественных и антропогенных факторов, определяющих территориальную неоднородность орнитокомплексов Приморья.
Материалы
Учеты птиц охватывают территорию Приморского края неравномерно (рис. 1) и выполнены преимущественно с середины мая до середины июля в период с 1961 по 1971 г. и с 1976 по 1982 г. разными методами. Последнее – наиболее существенная преграда при обобщающих работах, т. к. методические особенности учетов и индивидуальные склонности исследователей нередко уменьшают сопоставимость данных. В дальнейшем использованы данные упомянутых во Введении исследователей, наиболее полно отражающие количественные характеристики населения и оригинальные материалы автора, собранные в 1976–1982 гг. в окрестностях Владивостока, в Уссурийском заповеднике и Михайловском районе края.
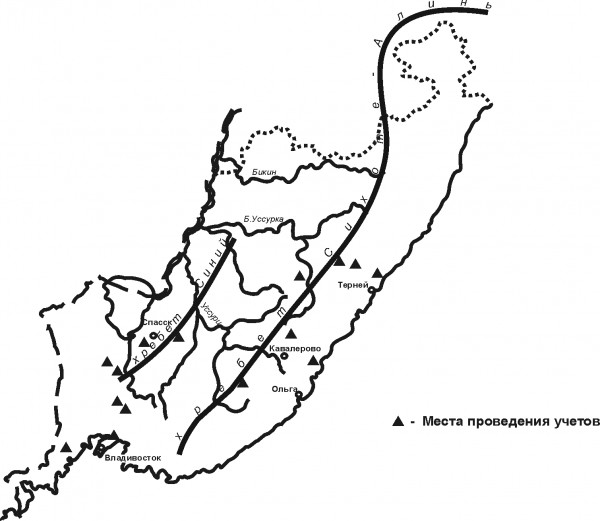
Рис. 1. Места сбора материалов
Fig. 1. Places of material collection
Методы
Учеты птиц автором данной статьи проведены по методике Ю. С. Равкина (Равкин, 1967). Методы учета по остальным материалам приведены в соответствующих публикациях. Общая протяженность учетных маршрутов С. В. Елсукова (Елсуков, 1975), Л. В. Кулешовой (Кулешова, 1976), Е. Л. Кушнарева (Кушнарев, 1982, 1984) и автора составляет более 587 км; в публикациях А. А. Назаренко (Назаренко, 1984) объем учетных работ не указан.
Для выявления пространственной дифференциации населения птиц использовали модифицированную меру сходства Сёренсена (Василевич, 1969). Эта мера сходства двух наборов А и В может быть определена как
К0 (А,В) = 2 ∑min (А, В) / (А + В) · 100 %,
где ∑min (А, В) – сумма минимальных значений компонента из двух сравниваемых. А и В – сумма компонентов в 1-м и 2-м местообитаниях (рис. 2).
При этом необходимо учитывать, что при анализе разнородного материала использование численности может привести к искажению сходства. Более подходящей в данном случае следует считать долю вида в населении. При анализе населения по доле формула несколько меняется:
К0 (А ,В) = ∑min (А, В).
Расчеты сходства проведены не только по населению, но и по фауне птиц. В дальнейшем структурные особенности населения взяты за основу типологических обобщений при составлении классификации населения птиц Приморья, при этом использована «Карта растительности бассейна Амура» (1969). Доминантами мы считали виды, составляющие не менее 5 % общей численности птиц. Названия птиц даны по Л. С. Степаняну (Степанян, 1975, 1978).
Результаты
Наиболее обособленно по сходству выступает население птиц поселков, луговых сообществ (табл. 1) и каменноберезняков, которые либо вообще не имеют сходства с другими лесными местообитаниями, либо это сходство чрезвычайно мало (Ко(А,В) ≤ 20–30 %). Самобытность птичьего населения поселков отмечали неоднократно. Здесь доминируют полевой воробей и сизый голубь (50 и 83 %). Суммарная численность орнитофауны – 715–1761 особь/км2. Поселки экологически разнородны даже для наиболее адаптированных к ним птиц: в зависимости от характера застройки, величины приусадебных участков, наличия условий для гнездования преобладают те или иные виды, вбирается в население большее или меньшее количество птиц, характерных для сопредельных природных биотопов. Поэтому даже в пределах этой небольшой группы по структурным особенностям можно выделить орнитоценозы, типичные для населенных пунктов сельской и городской застроек, различающихся участием в населении синантропных видов (полевого воробья, сизого голубя, в меньшей степени рыжепоясничной и деревенской ласточек, серого скворца и обыкновенной сороки).
Таблица 1. Население птиц (особей/км2) луговых местообитаний и поселков среди них
Table 1. Bird population (individuals/km2) in meadow habitats and adjacent settlments
| Вид | Местообитание, №* | ||
| 48 | 49 | 51 | |
| Воробей полевой | 0 | 0 | 355 |
| Ворона черная | 1 | 2 | 0 |
| Дубонос малый черноголовый | 0 | 0 | 3 |
| Дубровник | 66 | 115 | 0 |
| Жаворонок полевой | 43 | 2 | 0 |
| Зеленушка китайская | 0 | 0 | 10 |
| Иволга китайская | 0 | 0 | 7 |
| Камышевка дроздовидная | 0 | 0 | 3 |
| Камышевка пестроголовая | 7 | 18 | 0 |
| Конек степной | 15 | 3 | 0 |
| Кукушка обыкновенная | 3 | 1 | 0 |
| Ласточка деревенская | 6 | 0 | 90 |
| Ласточка рыжепоясничная | 0 | 0 | 30 |
| Лунь пегий | 1 | 2 | 0 |
| Овсянка ошейниковая | 58 | 133 | 22 |
| Перепел японский | 4 | 0 | 0 |
| Скворец малый | 0 | 0 | 3 |
| Скворец серый | 5 | 4 | 100 |
| Снегирь длиннохвостый | 1 | 2 | 0 |
| Сорока | 1 | 5 | 27 |
| Сорокопут сибирский | 0 | 0 | 20 |
| Трясогузка белая | 0 | 0 | 3 |
| Трясогузка желтая | 3 | 0 | 0 |
| Удод | 0 | 0 | 33 |
| Фазан | 2 | 1 | 7 |
| Цапля серая | <0.5 | 1 | 0 |
| Чекан черноголовый | 1 | 0 | 0 |
| Чибис | 1 | 2 | 0 |
| Всего | 218 | 291 | 713 |
Примечание. * – полный список местообитаний представлен в подписи к рис. 2.
В лугах начинают преобладать птицы открытых пространств, а также редколесий и кустарниковых зарослей. В первую очередь это овсянки (ошейниковая и дубровник), на долю которых приходится 57–85 % всего населения. Суммарное обилие всех видов 218–290 особей/км2. В зависимости от экологических условий соотношение видов претерпевает значительные изменения. В более увлажненных, вейниково-осоковых лугах в группу доминантов входит пестроголовая камышевка (6 % населения). В суходольных разнотравно-злаковых лугах доля ее вдвое меньше, но одновременно значительно больше полевого жаворонка (0.6 и 20 %) и степного конька (0.9 и 7 % населения). Участие остальных видов, как правило, незначительно.
Из лесных растительных формаций каменноберезняки с кедровым стлаником в нижнем ярусе наиболее обособлены по населению птиц. Они полностью приурочены к высотам более 1100–1200 м н. у. м. и представляют собой переходный пояс растительности от местообитаний с абсолютным господством древесных пород в верхнем ярусе к безлесным пространствам горных тундр. Тундры на территории Приморского края выражены сравнительно слабо, и на больших высотах каменноберезняки нередко занимают последний высотный пояс растительности. Доминируют пеночки – бурая и таловка (28 и 8 %), а также синехвостка, пятнистый конек и чиж (22, 14 и 5 %). Суммарное обилие птиц сравнительно высоко – 540 особей/км2. Преобладают птицы редколесий и кустарниковых зарослей, многоярусники – дрозды и кронники (44, 22 и 19 %), что хорошо согласуется с ярусной структурой растительности каменноберезняков.
Форпостами обширных лесных массивов на западе и юге Приморья являются биогеоизоляты, «островные» леса – участки лесной растительности, сохранившиеся среди трансформированных сельскохозяйственной деятельностью ландшафтов (табл. 2). По соотношению видов в населении птиц изоляты можно разделить на две группы. К первой относятся местообитания, занимающие значительные площади (десятки или сотни гектаров), приуроченные к нераспаханным вершинам сопок и имеющие, как правило, хорошо сформированный растительный покров с древостоем среднего или более старшего возраста. Вторая группа представлена небольшими колками, подчас с сильно изрезанной границей. Эти особенности фитоценозов обусловливают специфику орнитоценозов. В крупных лесах сравнительно невысока доля птиц редколесий и кустарниковых зарослей – 7–24 % населения (в мелких изолятах – 57–80 %) и значительно участие лесных видов, например, среднеярусников-наземников – 18–28 % населения (в мелких «островных» лесах – от 0 до 9 %). Поэтому и доминирующие виды существенно различны. В первой группе это малый черноголовый дубонос, восточная синица, желтогорлая овсянка. Во второй – седоголовая и ошейниковая овсянки, сибирский сорокопут, большая горлица, местами с пестроголовой камышевкой, японским перепелом и дубровником – типичными представителями луговых ценозов. Имеется немалое число общих доминантов.
Таблица 2. Население птиц (особей/км2) «островных» лесов
Table 2. Bird population (individuals/km2) of "island" forests
| Вид | Местообитание, № | ||
| 52 | 53 | 54 | |
| Белоглазка | 0 | 2 | 6 |
| Воробей полевой | 40 | 0 | 0 |
| Ворона черная | 7 | 1 | 0 |
| Горлица большая | 11 | 3 | 61 |
| Дрозд сизый | 0 | 0 | 109 |
| Дубонос малый черноголовый | 0 | 0 | 152 |
| Дятел белоспинный | 0 | 0 | 4 |
| Зеленушка китайская | 7 | 1 | 0 |
| Иволга китайская | 1 | 5 | 0 |
| Камышевка пестроголовая | 27 | 0 | 0 |
| Камышевка толстоклювая | 58 | 0 | 0 |
| Короткохвостка | 0 | 0 | 76 |
| Кукушка обыкновенная | 0 | 1 | 0 |
| Ласточка рыжепоясничная | 7 | 0 | 0 |
| Личинкоед | 0 | 0 | 6 |
| Мухоловка желтоспинная | 0 | 0 | 95 |
| Мухоловка синяя | 0 | 0 | 13 |
| Овсянка желтогорлая | 0 | 0 | 196 |
| Овсянка ошейниковая | 49 | 16 | 0 |
| Овсянка седоголовая | 13 | 8 | 0 |
| Перепел японский | 27 | 0 | 0 |
| Синица восточная | 0 | 4 | 234 |
| Синица длиннохвостая | 0 | 0 | 38 |
| Скворец серый | 0 | 1 | 44 |
| Снегирь длиннохвостый | 11 | 4 | 0 |
| Соловей синий | 0 | 0 | 64 |
| Сорока | 5 | 2 | 0 |
| Сорока голубая | 0 | 0 | 19 |
| Сорокопут сибирский | 0 | 20 | 0 |
| Стриж иглохвостый | 0 | 1 | 0 |
| Удод | 2 | 1 | 0 |
| Фазан | 7 | 2 | 0 |
| Цапля серая | 1 | 0 | 0 |
| Всего | 273 | 72 | 1117 |
Суммарное обилие птиц колеблется в пределах 70–120 особей/км2. По данным Е. Л. Кушнарева (1982), в мелких «островных» лесах площадью до 1 га оно составляет 2000–4000 особей/км2. Для «островных» полезащитных лесов Литвы и Средней Европы значения выше (1922–6140 и 2000–10000 особей/км2) (Курлавичюс, 1986).
По соотношению видов в населении птиц все хвойные и хвойно-широколиственные леса можно разделить на четыре крупных блока: 1 – коренные (условно коренные) леса и 2 – хвойно-широколиственные леса восточного и западного макросклонов Сихотэ-Алиня, испытывающие значительное антропогенное воздействие; 3 и 4 – их аналоги на западном макросклоне хребта (табл. 3 и 4).
Таблица 3. Население птиц (особей/км2) хвойно-широколиственных лесов
Table 3. Bird population (individuals/km2) of coniferous-broadleaf forests
| Вид | Местообитания | |
| 46 | 47 | |
| Белоглазка | 0 | 2 |
| Ворона большеклювая | 13 | 3 |
| Гаички: буроголовая и черноголовая | 33 | 22 |
| Горлица большая | 4 | 4 |
| Дрозд бледный | 4 | 8 |
| Дрозд пестрый | 0 | 2 |
| Дрозд сизый | 2 | 1 |
| Дубонос большой черноголовый | 47 | 24 |
| Дубонос | 13 | 2 |
| Дятел белоспинный | 18 | 7 |
| Дятел малый острокрылый | 0 | 4 |
| Желна | 0 | 5 |
| Короткохвостка | 74 | 54 |
| Кукушка глухая | 2 | 8 |
| Кукушка ширококрылая | 0 | 3 |
| Личинкоед | 2 | 5 |
| Московка | 24 | 13 |
| Мухоловка желтоспинная | 0 | 2 |
| Мухоловка синяя | 11 | 31 |
| Мухоловка таежная | 0 | 4 |
| Мухоловка ширококлювая | 33 | 71 |
| Овсянка желтогорлая | 27 | 2 |
| Таежная овсянка | 47 | 58 |
| Пеночка бледноногая | 4 | 30 |
| Пеночка зеленая | 0 | 2 |
| Пеночка корольковая | 50 | 47 |
| Пеночка светлоголовая | 31 | 40 |
| Пищуха | 0 | 8 |
| Поползень | 28 | 35 |
| Рябчик | 0 | 8 |
| Соловей синий | 78 | 59 |
| Широкорот | 0 | 1 |
| Всего | 545 | 564 |
Таблица 4. Население птиц (особей/км2) широколиственных лесов
Table 4. Bird population (individuals/km2) of broad-leaved forests
| Вид | Местообитания | ||||||
| 31 | 43 | 44 | 45 | 50 | 55 | 56 | |
| Белоглазка | 0 | 22 | 10 | 21 | 0 | 0 | 0 |
| Ворона большеклювая | 0 | 6 | 9 | 7 | 0 | 5 | 4 |
| Ворона черная | 0 | 0 | 0 | 0 | 2 | 0 | 0 |
| Гаички: буроголовая и черноголовая | 50 | 50 | 36 | 33 | 0 | 117 | 0 |
| Горлица большая | 0 | 0 | 4 | 5 | 5 | 4 | 0 |
| Дрозд бледный | 0 | 0 | 0 | 1 | 0 | 5 | 0 |
| Дрозд пестрый | 0 | 0 | 0 | 5 | 0 | 0 | 0 |
| Дрозд сизый | 10 | 3 | 24 | 16 | 3 | 20 | 0 |
| Дубонос большой черноголовый | 0 | 11 | 0 | 26 | 0 | 0 | 0 |
| Дубонос малый черноголовый | 10 | 0 | 0 | 0 | 15 | 10 | 0 |
| Дубонос | 0 | 0 | 6 | 9 | 0 | 13 | 10 |
| Дятел белоспинный | 20 | 25 | 8 | 19 | 0 | 0 | 0 |
| Дятел большой пестрый | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Дятел малый острокрылый | 0 | 0 | 0 | 9 | 0 | 0 | 0 |
| Дятел малый пестрый | 0 | 0 | 0 | 6 | 0 | 0 | 0 |
| Дятел седой | 0 | 0 | 0 | 0 | 0 | 7 | 0 |
| Желна | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Зеленушка китайская | 0 | 0 | 26 | 6 | 0 | 8 | 0 |
| Зуек малый | 0 | 0 | 6 | 0 | 0 | 0 | 0 |
| Иволга китайская | 0 | 0 | 6 | 0 | 5 | 3 | 0 |
| Камышевка дроздовидная | 0 | 0 | 0 | 0 | 0 | 13 | 0 |
| Камышевка пестроголовая | 0 | 0 | 10 | 0 | 0 | 20 | 5 |
| Камышевка толстоклювая | 0 | 0 | 59 | 0 | 5 | 0 | 0 |
| Конек пятнистый | 0 | 0 | 0 | 0 | 0 | 25 | 74 |
| Короткохвостка | 0 | 0 | 0 | 122 | 0 | 0 | 0 |
| Кукушка глухая | 10 | 3 | 5 | 10 | 0 | 3 | 0 |
| Кукушка индийская | 0 | 0 | 1 | 0 | 0 | 6 | 6 |
| Кукушка обыкновенная | 0 | 0 | 1 | 1 | 1 | 3 | 4 |
| Кукушка ширококрылая | 0 | 0 | 0 | 2 | 0 | 0 | 0 |
| Личинкоед | 10 | 17 | 7 | 14 | 0 | 25 | 20 |
| Московка | 0 | 0 | 0 | 4 | 0 | 0 | 0 |
| Мухоловка желтоспинная | 70 | 47 | 136 | 63 | 0 | 57 | 20 |
| Мухоловка синяя | 10 | 39 | 5 | 55 | 0 | 0 | 0 |
| Мухоловка ширококлювая | 10 | 141 | 45 | 150 | 0 | 31 | 0 |
| Неясыть длиннохвостая | 0 | 0 | 8 | 0 | 0 | 0 | 0 |
| Овсянка желтогорлая | 30 | 83 | 0 | 82 | 40 | 70 | 0 |
| Овсянка ошейниковая | 0 | 0 | 58 | 0 | 0 | 0 | 0 |
| Овсянка седоголовая | 110 | 0 | 195 | 42 | 15 | 325 | 122 |
| Овсянка таежная | 0 | 0 | 3 | 45 | 0 | 0 | 0 |
| Пеночка бледноногая | 20 | 0 | 19 | 22 | 0 | 0 | 0 |
| Пеночка зеленая | 0 | 0 | 0 | 2 | 0 | 0 | 0 |
| Пеночка корольковая | 0 | 0 | 0 | 15 | 0 | 0 | 0 |
| Пеночка светлоголовая | 30 | 22 | 8 | 102 | 0 | 144 | 60 |
| Пищуха | 0 | 0 | 0 | 6 | 0 | 0 | 0 |
| Погоныш большой | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Поползень | 20 | 56 | 22 | 44 | 0 | 75 | 9 |
| Синица восточная | 150 | 125 | 32 | 31 | 10 | 17 | 27 |
| Синица длиннохвостая | 0 | 33 | 6 | 3 | 0 | 50 | 0 |
| Скворец серый | 0 | 8 | 2 | 2 | 19 | 83 | 0 |
| Снегирь длиннохвостый | 0 | 0 | 22 | 3 | 0 | 33 | 0 |
| Сойка | 0 | 8 | 0 | 1 | 0 | 0 | 0 |
| Соловей синий | 100 | 28 | 28 | 34 | 0 | 9 | 0 |
| Сорока | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| Сорока голубая | 30 | 0 | 9 | 0 | 0 | 4 | 0 |
| Сорокопут сибирский | 10 | 0 | 6 | 0 | 8 | 0 | 3 |
| Трясогузка белая | 0 | 0 | 6 | 0 | 0 | 17 | 0 |
| Трясогузка горная | 0 | 0 | 0 | 2 | 0 | 13 | 0 |
| Трясогузка древесная | 0 | 0 | 0 | 0 | 0 | 0 | 11 |
| Удод | 0 | 0 | 0 | 0 | 12 | 0 | 0 |
| Чекан черноголовый | 0 | 0 | 25 | 0 | 0 | 0 | 0 |
| Широкорот | 0 | 0 | 3 | 6 | 0 | 0 | 0 |
| Всего | 700 | 727 | 858 | 1027 | 141 | 1208 | 375 |
Различия в соотношении видов птиц в населении восточного и западного макросклонов обусловлены, в первую очередь, географическим положением местообитаний и степенью освоенности прилегающих территорий. Так, хребет Синий окружен с одной стороны Приханкайской равниной, а с другой – широкой, хорошо освоенной долиной реки Арсеньевка. В нижнем поясе растительности обычно преобладают широколиственные и мелколиственно-широколиственные, нередко вторичные леса. В населении птиц это влечет за собой уменьшение доли таежных видов и увеличение неморальных. Так, доля корольковой пеночки и московки составляет в оптимальных биотопах по 4 % (в лесах восточного макросклона – 12 и 25 % соответственно). С другой стороны, даже в коренных лесах западного макросклона участие светлоголовой пеночки составляет 18–20 % против 1–4 % на восточном макросклоне. Для хвойно-широколиственных лесов хребта Синий характерна и желтоспинная мухоловка (до 6 % населения), тогда как на восточном макросклоне в аналогичных типах местообитаний она отсутствует.
В группу мелколиственно-широколиственных лесов западного макросклона входят местообитания, характерные для хребта Синий, которые отнесены к одной группе с дубняками и долинными полидоминантными лесами юго-западной периферии Сихотэ-Алиня. Аналогичная картина свойственна и вторичным мелколиственным лесам Восточного Сихотэ-Алиня. Основное фитоценотическое отличие этих биотопов – резко ослабленные позиции широколиственных пород, в частности дуба, и нередко присутствие в древостое кедра как представителя той растительной формации, на месте которой формировались восточные леса. Этими же особенностями обусловлены различия населения птиц западного и восточного макросклонов. Принципиально не различаясь по ярусной структуре населения, они, как и хвойно-широколиственные леса, отличаются участием в населении птиц элементов неморального и бореального фаунистических комплексов. Например, участие желтоспинной мухоловки и большого черноголового дубоноса (неморальный фаунистический комплекс) в лесах Западного Сихотэ-Алиня составляет до 17 и 2 % соответственно. На восточном макросклоне первый вид в учетах встречен единично, а второй – отсутствует. С другой стороны, рябчик (бореальный фаунистический комплекс), отсутствующий в лесах хребта Синий, характерен для Восточного Сихотэ-Алиня (до 5 %). Суммарное обилие птиц мелколиственных и мелколиственно-широколиственных лесов разных районов Приморья примерно одного уровня.
В коренных слабо нарушенных хвойных и хвойно-широколиственных лесах прослежено непрерывное изменение структуры населения птиц в соответствии с высотно-поясными отличиями растительности. Такая смена еще раз подтверждает представление об известной автономии отдельных компонентов орнитоценозов (Кулешова, 1976) и о постепенности изменений населения при континуальном переходе одной растительной формации в другую. Это четко проявляется, если учеты проведены в значительном числе промежуточных типов местообитаний, тогда как при рассмотрении крайних сообществ преобладает дискретность, которая, по-видимому, прежде всего должна проявляться в смене доминирующих видов птиц. Исходя из этого все варианты населения проанализированы по сходству фаунистического состава доминантов. Для хвойных лесов это позволило в значительной степени снять «шумы», вызванные антропогенной деятельностью, и обособить несколько групп местообитаний, в первую очередь – пихтово-еловые леса. Из доминантов в них наиболее характерны желтоголовый королек, зеленая пеночка, таежная мухоловка и поползень. Поползень преобладает почти во всех хвойно-широколиственных лесах Приморья. В зеленомошных ельниках к ним добавляются крапивник и синехвостка. Последняя наиболее многочисленна в каменноберезняках. В травяных ельниках обычными становятся корольковая пеночка и московка.
В самостоятельную группу выделены и кедрово-еловые леса, к которым, видимо, следует отнести и северные кедровники. Основное отличие их от типичных кедровников – это доминирование, среди прочих характерных для кедрачей видов, таежной мухоловки, местами с желтоголовым корольком, которые наиболее типичны для ельников.
Также обособляется в самостоятельную группу население типичных кедровников и чернопихтово-широколиственных лесов. Последние характеризуются значительным участием среди доминантов неморальных фаунистических элементов, например, светлоголовой пеночки, большого черноголового дубоноса и др.
Анализ сходства населения птиц исследованных местообитаний позволил объединить их в некоторые структурно-однородные блоки и провести типизацию орнитоценозов для построения зоогеографической карты населения птиц Приморья (рис. 2, табл. 5, 6). Картографическая основа ограничила возможность детально охарактеризовать пространственную структуру населения, хотя связь населения птиц с высотной поясностью растительности прослежена достаточно четко. Отсутствие материалов для зоны сельскохозяйственного освоения и некоторых типов местообитаний в ряде районов края не позволило также в полном объеме отразить основные черты пространственной дифференциации сообществ птиц.
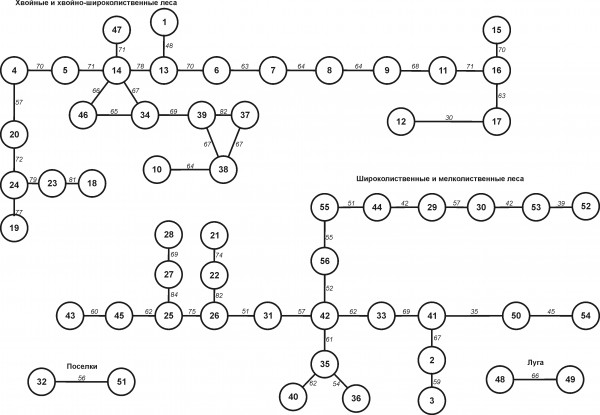
Рис. 2. Пространственная дифференциация населения птиц по сходству фаунистического состава: 1–56 – местообитания (см. табл. 5). Цифры между обозначениями населения птиц обследованных местообитаний – коэффициенты сходства Сёренсена
Fig. 2. Spatial differentiation of the bird population according to the similarity of faunal composition: 1 - 56 - habitats (see tabl. 5). The numbers between the symbols of the bird population in surveyed habitats – factors of Sorensen similarity
Таблица 5. Местообитания птиц
Table 5. Bird habitats
| 1 | Приморские дубняки | [Coastal oak forests] |
| 2 | Мелколиственные осиново-березовые леса | [Small-leaved aspen-birch forests] |
| 3 | Мелколиственные осиново-березовые долинные леса | [Small-leaved aspen-birch valley forests] |
| 4 | Широколиственные долинные леса | [Broad-leaved valley forests] |
| 5 | Кедрово-широколиственные леса на низких надпойменных террасах | [Cedar-broadleaved forests on low terraces above the floodplain] |
| 6 | Кедрово-широколиственные леса на высоких надпойменных террасах | [Cedar-broadleaved forests on high terraces above the floodplain] |
| 7 | Кедрово-широколиственные леса на склонах гор южных экспозиций | [Cedar-broadleaved forests on the slopes of the southern exposure mountains] |
| 8 | Кедрово-широколиственные леса на склонах гор северных экспозиций | [Cedar-broadleaved forests on the slopes of the northern exposure mountains] |
| 9 | Кедрово-еловые леса | [Cedar - spruce forests] |
| 10 | Кедрово-еловые леса с примесью широколиственных пород в верховьях рек | [Cedar - spruce forests with an admixture of broad-leaved species in headwaters of the river] |
| 11 | Пихтово-еловые леса | [Fir-spruce forests] |
| 12 | Каменноберезняки | [Erman's birch forests] |
| 13 | Чернопихтово-широколиственные леса | [Blackfir-broadleaved forests] |
| 14 | Хвойно-широколиственные леса | [Coniferous- broadleaved forests] |
| 15 | Кедрово-еловые леса в верховьях реки Уссури | [Cedar - spruce forests in headwaters of the Ussuri River] |
| 16 | Кедрово-еловые леса в верховьях реки Большой Уссурки | [Cedar - spruce forests in headwaters of the Big Ussurka River] |
| 17 | Пихтово-еловые зеленомошные леса горы Облачной | [Fir-spruce green-mossy forests of the Mountain Cloud] |
| 18 | Елово-кедрово-широколиственные коренные леса | [Spruce-cedar-broadleaved native forests] |
| 19 | Елово-кедрово-широколиственные леса, пройденные рубками | [Spruce-cedar-broadleaved forests covered by logging] |
| 20 | Кедрово-елово-широколиственные долинные леса, пройденные рубками | [Spruce-cedar-broadleaved valley forests covered by logging] |
| 21 | Мелколиственные леса на гари (50–60 лет) | [Small-leaved forests in place of burnt (50-60 years)] |
| 22 | Мелколиственно-липово-дубовые долинные леса, пройденные рубками | [Small-leaved-lime-oak-forest covered by logging] |
| 23 | Кедрово-широколиственные коренные леса | [Cedar-broadleaved native forests] |
| 24 | Кедрово-широколиственные леса, пройденные рубками | [Cedar-broadleaved forests covered by logging] |
| 25 | Мелколиственно-липово-дубовые долинные леса, пройденные рубками | [Small-leaved-lime-oak-forest valley covered by logging] |
| 26 | Березово-осиновые леса с примесью липы (50–60 лет) | [Birch-aspen forests with an admixture of lime (50-60 years)] |
| 27 | Липово-ясенево-дубовые леса | [Lime-ash-oak forests] |
| 28 | Мелколиственно-широколиственные полуостровные леса | [Small-leaved-broad-leaved peninsular forests] |
| 29 | Мелколиственно-широколиственные леса с выпасом скота | [Small-leaved-broad-leaved forests with grazing] |
| 30 | Мелколиственно-широколиственные леса без выпаса скота | [Small-leaved-broad-leaved forests without grazing] |
| 31 | Ясенево-дубовые леса | [Ash-oak forests] |
| 32 | Поселок Хрустальный | [The Khrustalny village] |
| 33 | Мелколиственные леса | [Small-leaved forests] |
| 34 | Кедрово-широколиственные леса | [Cedar-broadleaved forests] |
| 35 | Кедрово-широколиственные редины | [Cedar-broadleaved sparse forests] |
| 36 | Гари | [Burnt forests] |
| 37 | Пихтово-еловые леса | [Fir-spruce forests] |
| 38 | Пихтово-еловые долинные леса | [Fir-spruce valley forests] |
| 39 | Пихтово-кедровые леса | [Fir- cedar forests] |
| 40 | Ольхово-березовые долинные леса | [Alder-birch valley forests] |
| 41 | Дубняки, мелколиственные леса | [Oak forests, small-leaved forests] |
| 42 | Широколиственные полидоминантные леса | [Broadleaved polydominant forests] |
| 43 | Дубняки | [Oak forests] |
| 44 | Перелески | [Coppices] |
| 45 | Широколиственные полидоминантные долинные леса | [Broadleaved polydominant valley forests] |
| 46 | Чернопихтово-широколиственные леса | [Blackfir-broadleaved forests] |
| 47 | Хвойно-широколиственные леса | [Coniferous- broadleaved forests] |
| 48 | Разнотравно-злаковые луга | [Motley-grass meadow] |
| 49 | Вейниково-осоковые луга | [Vein-sedge meadows] |
| 50 | Дубняки | [Oak forests] |
| 51 | Поселок Перелётный | [The Pereletny village] |
| 52 | Дубовые островные леса, занимающие площадь менее 10 га | [Oak outlier forests occupying an area of less than 10 hectares] |
| 53 | Дубовые островные леса | [Oak outlier forests] |
| 54 | Дубово-липово-мелколиственные островные леса | [Oak-lime-small-leaved outlier forest] |
| 55 | Мелколиственно-широколиственные долинные леса | [Small-leaved-broad-leaved valley forests] |
| 56 | Приморские дубняки | [Coastal oak forests] |
Таблица 6. Классификация летнего населения птиц Приморья
Table 6. Classification of the summer bird population in Primorye
| Местообитания | Доминирующие виды |
| 1 | 2 |
| КАМЕННОБЕРЕЗНЯКИ: | Пеночки – бурая и таловка, синехвостка, пятнистый конек, чиж |
| ВЫСОКОГОРНЫЕ ЕЛЬНИКИ: | Желтоголовый королек, зеленая пеночка, таежная мухоловка, поползень |
| 1) зеленомошные | Синехвостка, крапивник |
| 2) травяные: | |
| а – на склонах | Корольковая пеночка, московка |
| б – в долинах | Бледноногая пеночка, соловьи – свистун и синий |
| ХВОЙНО-ШИРОКОЛИСТВЕННЫЕ ЛЕСА (кроме западного макросклона): | Поползень, корольковая пеночка, синий соловей, ширококлювая мухоловка |
| 1) южные кедровники и чернопихтарники | Гаички – черноголовая и буроголовая, короткохвостка, светлоголовая пеночка: местами с таежной овсянкой и большим черноголовым дубоносом |
| 2) типичные кедровники | Московка, местами с таежной овсянкой и короткохвосткой |
| 3) северные кедровники и кедрово-еловые леса | Таежная мухоловка, местами с желтоголовым корольком |
| ХВОЙНО-ШИРОКОЛИСТВЕННЫЕ ЛЕСА западного макросклона: | Таежная и желтогорлая овсянки, светлоголовая пеночка, синий соловей, желтоспинная мухоловка |
| 1) коренные, слабо нарушенные | Поползень, ширококлювая мухоловка |
| 2) вторичные мелколиственные и мелколиственно-широколиственные леса | Белоспинный дятел, личинкоед; местами с восточной синицей |
| ХВОЙНО-ШИРОКОЛИСТВЕННЫЕ ЛЕСА ПРИДОЛИННОГО КОМПЛЕКСА: | Синий соловей, бледноногая пеночка, таежная овсянка, ширококлювая мухоловка, поползень |
| 1) коренные, слабо нарушенные | Корольковая пеночка, короткохвостка; местами с московкой |
| 2) вторичные мелколиственные и мелколиственно-широколиственные леса | Светлоголовая пеночка; местами с синей мухоловкой |
| Производные КЕДРОВО-ШИРОКОЛИСТВЕННЫХ ЛЕСОВ: | Желтогорлая овсянка, светлоголовая пеночка |
| 1) редины | Поползень, московка, таежная овсянка, седоголовая овсянка, толстоклювая пеночка |
| 2) слабо возобновившиеся гари | Толстоклювая пеночка, красноухая овсянка, пятнистый конек |
| 3) мелколиственные леса | Синий соловей, синяя мухоловка; местами с бледным дроздом и гаичками – черноголовой и буроголовой |
| Полидоминантные ШИРОКОЛИСТВЕННЫЕ ЛЕСА: | Светлоголовая пеночка, поползень |
| 1) коренные ненарушенные | Синий соловей, бледноногая пеночка, таежная овсянка, ширококлювая мухоловка; местами с чижом |
| 2) нарушенные и вторичные мелколиственные леса | Желтогорлая и седоголовая овсянки; местами с пятнистым коньком и гаичками – черноголовой и буроголовой |
| МЕЛКОЛИСТВЕННО-ШИРОКОЛИСТВЕННО-ДУБОВЫЕ ЛЕСА: | Седоголовая овсянка, восточная синица |
| 1) сплошные массивы | |
| а – прибрежной полосы восточного макросклона | Светлоголовая пеночка, гаички – черноголовая и буроголовая; местами с личинкоедом, пятнистым коньком |
| б – южного Приморья | Синий соловей, гаички – черноголовая и буроголовая, желтоспинная мухоловка |
| 2) изолированные лесные массивы среди нелесной растительности | |
| а – молодые | Ошейниковая овсянка, длиннохвостый снегирь; местами с толстоклювой камышевкой, соловьем-красношейкой |
| б – спелые | Малый черноголовый дубонос, китайская иволга, желтоспинная мухоловка, сизый дрозд, большая горлица, желтогорлая овсянка |
| ЛУГА: | Ошейниковая овсянка, дубровник |
| 1) разнотравно-злаковые (суходольные) | Полевой жаворонок, степной конек |
| 2) вейниково-осоковые (увлажненные) | Пестроголовая камышевка |
| НАСЕЛЕННЫЕ ПУНКТЫ | Полевой воробей |
| 1) городской застройки | Сизый голубь, местами с сорокой |
| 2) сельской застройки | Серый скворец, деревенская и рыжепоясничная ласточки, сибирская горихвостка |
Заключение
В результате исследований построенная иерархическая классификация орнитокомплексов включает 10 типов населения, 21 подтип и 6 классов. Различия типов населения связаны с застроенностью, облесенностью и составом лесообразующих пород, подтипов: в лесах – с особенностями наземного яруса растительности, экспозицией склонов, коренным или вторичным характером лесов, площадью и окружением их; в лугах – с увлажнением, а в селитебных местообитаниях – с характером застройки (городской и сельской). Отличия классов населения коррелируют в лесах с рельефом, возрастом насаждений и их местоположением.
Библиография
Василевич В. И. Статистические методы в геоботанике . Л.: Наука, 1969. 232 с.
Елсуков С. В. Летнее население птиц в кедровниках Сихотэ-Алинского заповедника // Сихотэ-Алинский биосферный район: фоновое состояние природных компонентов. Владивосток: ДВО АН СССР, 1987. С. 109–115.
Исаков Ю. А. Состояние изученности авифауны СССР // Птицы СССР. История изучения. Гагары, поганки, трубконосые. М.: Наука, 1982. С. 208–227.
Карта растительности бассейна Амура . Масштаб 1:2500000 / Под ред. В. Б. Сочава. М.: ГУГК, 1969.
Кулешова Л. В. Закономерности обособления типов населения птиц в лесах Среднего Сихотэ-Алиня // Орнитология. М.: МГУ, 1976. Вып. 12. С. 26–54.
Курлавичюс П. Биотопическое распределение птиц в агронасаждениях . Вильнюс: Моксклас, 1986. 106 с.
Кушнарев Е. Л. Влияние хозяйственной деятельности на распределение и численность птиц в очагах клещевого энцефалита Западного Сихотэ-Алиня // Влияние хозяйственной деятельности на структуру природных очагов клещевого энцефалита в Приморском крае. Владивосток: ДВНЦ АН СССР, 1982. С. 80–89.
Кушнарев Е. Л. Антропогенные сукцессии орнитосообществ и территориальные связи местообитаний Западного Сихотэ-Алиня // Фаунистика и биология птиц юга Дальнего Востока. Владивосток: ДВНЦ АН СССР, 1984. С. 71–78.
Назаренко А. А. Птицы чернопихтово-широколиственных лесов и южных кедровников // Биогеоценотические исследования в лесах Приморья. Л.: Наука, 1968. С. 134–149.
Назаренко А. А. Птицы вторичных широколиственных лесов южного Приморья и некоторые аспекты формирования природных сообществ // Орнитологические исследования на юге Дальнего Востока. Владивосток: ДВНЦ АН СССР, 1971. С. 79–97.
Назаренко А. А. Птичье население смешанных и темнохвойных лесов Южного Приморья, 1962–1971 гг. // Фаунистика и биология птиц юга Дальнего Востока. Владивосток: ДВНЦ АН СССР, 1984. С. 60–70.
Равкин Ю. С. К методике учета птиц лесных ландшафтов // Природа очагов клещевого энцефалита на Алтае. Новосибирск: Наука, 1967. С. 66–75.
Симонов С. Б. Закономерности пространственной дифференциации населения птиц Среднего Сихотэ-Алиня : Автореф. дис. … канд. геогр. наук. М., 1985. 22 с.
Симонов С. Б. Пространственное распределение населения птиц антропогенных районов Сихотэ-Алинского биосферного района // Сихотэ-Алинский биосферный район: фоновое состояние природных компонентов. Владивосток: ДВНЦ АН СССР, 1987. С. 116–127.
Степанян Л. С. Состав и распределение птиц фауны СССР. Неворобьиные . М.: Наука, 1975. 372 с.
Степанян Л. С. Состав и распределение птиц фауны СССР. Воробьиные . М.: Наука, 1978. 392 с.

 © 2011 - 2026
© 2011 - 2026