



Введение
Саранчовые уже много лет являются популярным объектом экологических, в том числе эколого-морфологических, исследований. В настоящее время существует несколько систем жизненных форм этих насекомых (Бей-Биенко, Мищенко, 1951; Стебаев, 1970, 1987; Черняховский, 1970; Правдин, 1978; Стебаев, Омельченко, 1981), построенных с использованием преимущественно морфологических признаков, которым приписывается то или иное адаптивное значение.
Важным комплексом признаков, традиционно учитываемым при характеристике жизненных форм саранчовых, являются окраска и рисунок тела в целом и отдельных его частей. Речь при этом может идти как о выраженности определенных их элементов, так и об общем цвете тела. При этом функциональное значение окраски и ее элементов у саранчовых в разных случаях может быть разным. В частности, у некоторых ядовитых видов речь, по-видимому, может идти об апосематической (предостерегающей) окраске (см. обзор: Rowell, 1972). Кроме того, расположение темных и светлых пятен, полос и перевязей у саранчовых может играть роль в терморегуляции (Присный, 1988), а ярко окрашенные элементы рисунка на теле и конечностях – в визуальной коммуникации (Стебаев, 1990).
Однако наибольшее значение для этой группы имеет, несомненно, покровительственная окраска, защищающая от хищников (Morse, 1907; Rowell, 1972). В частности, принято считать, что для геофильных саранчовых характерна землистая окраска, а для обитателей толщи растительного покрова – окраска под цвет живых или высохших растений (Бей-Биенко, Мищенко, 1951; Rowell, 1972).
Следует, однако, иметь в виду, что для многих саранчовых характерна внутривидовая изменчивость, так что говорить о какой-либо видоспецифической окраске у таких видов оказывается невозможно. В разных группах саранчовых эта изменчивость характеризуется разными особенностями, как затрагивающими ее проявление, так и обуславливающими ее механизмы. В частности, изменчивы могут быть окраска задних крыльев (например, у некоторых представителей подсемейства Oedipodinae: Mioscirtus wagneri, Angaracris barabensis, Celes variabilis, Bryodema spp., Bryodemella spp. – Бей-Биенко, Мищенко, 1951; Бенедиктов, 1998, 2016) и выраженность рисунка (в частности, у целого ряда представителей трибы Gomphocerini s. l. из подсемейства Gomphocerinae – Воронцовский, 1928). Широко распространена у саранчовых также и изменчивость фоновой окраски тела, причем у представителей разных подсемейств она обусловлена разными механизмами. Так, у представителей рода Oedipoda (подсемейство Oedipodinae) она не только носит ненаследственный характер, но и может изменяться в соответствии с окружающим фоном (Levita, 1970; Moreteau, 1975; Yerushalmi, Pener, 2002). У относящейся к этому же подсемейству перелетной саранчи Locusta migratoria окраска тела также ненаследственна и зависит от принадлежности к одиночной или стадной фазе (Uvarov, 1921). Зависимость окраски тела от плотности популяции известна также и у представителей относящегося к подсемейству Catantopinae s. l. рода Schistocerca, как стадных, так и одиночных (Gotham, Song, 2013). В отношении некоторых нестадных саранчовых из других подсемейств, в том числе представителей рода Chorthippus s. l., входящего в трибу Gomphocerini подсемейства Gomphocerinae, также имеются данные о связи окраски тела с выраженностью миграционного поведения (Rubtzov, 1935). В то же время для некоторых представителей этого рода доказана наследственная природа фоновой окраски и рисунка тела (см. обзор: Rowell, 1972). Любопытно также, что для целого ряда представителей этого подсемейства, этой трибы и этого рода описаны гомологические ряды изменчивости окраски (Воронцовский, 1928; Rubtzov, 1935).
Таким образом, представление о существовании покровительственной окраски, «характерной» или «не характерной» для тех или иных жизненных форм саранчовых, может скрывать под собой сложные картины внутрипопуляционной цветовой дифференциации, опирающейся в разных случаях на разные механизмы. Вполне правдоподобным кажется предположение, что адаптивное значение может иметь не только каждая из отдельно взятых цветовых форм, но и их количественное соотношение в популяции (которое в случаях наследственной природы этой изменчивости может поддерживаться дизруптивным отбором). В терминологии, предложенной нами ранее, речь идет о распределении вариантов покровительственной окраски в популяции как о важной адаптивной части ее метафенотипа (Озерский, 2010) и как о части реализующейся на популяционном уровне жизненной формы (Озерский и др., 2011).
При этом открытым остается вопрос о степени пластичности этого компонента жизненной формы. В настоящее время предполагается, что наиболее консервативными с эволюционной точки зрения признаками жизненных форм являются морфологические (Горохов, 1983), однако до сих пор они сравнивались только с «образом жизни», но не с метафенотипическими компонентами жизненной формы. В то же время, исходя из базовых представлений современных селекционистских концепций, трактующих популяцию как элементарную единицу эволюции (Яблоков, 1987), есть все основания предполагать, что именно изменение количественных соотношений между фенотипами является наиболее оперативным адаптивным ответом на изменение условий обитания популяции и, соответственно, наиболее быстро осуществляемым преобразованием популяционной жизненной формы.
Для проверки этого предположения нами уже несколько лет осуществляется сравнительное исследование представленности различных вариантов окраски у полиморфных видов прямокрылых. В настоящее время можно, в частности, считать доказанным, что существование одних и тех же или сходных цветовых форм, в том числе гомологических рядов изменчивости, у разных видов этих насекомых не означает одних и тех же частот встречаемости этих форм даже у близких видов (Озерский, 2012, 2014). В то же время степень постоянства этих частот в разных популяциях одних и тех же видов все еще требует изучения.
Одним из видов, перспективных для проведения подобных исследований, можно считать зеленую травянку (Omocestus viridulus) – вид саранчовых, широко распространенный в Европе (включая Европейскую часть России) и в Сибири (Бей-Биенко, Мищенко, 1951), относящийся к трибе Gomphocerini s. l., для ряда представителей которой доказана наследуемость окраски, и обладающий выраженным полиморфизмом окраски, в том числе наличием хорошо распознаваемой непосредственно в поле «зеленой» формой (f. viridis по терминологии И. А. Рубцова (Rubtzov, 1935), Б. Петерсена и Дж. Трехерна (Petersen, Treherne, 1949)). Данная форма характеризуется зеленой окраской лица, щек, боковых лопастей переднеспинки и плейритов средне- и заднегруди у обоих полов (рис. 1а, б), в то время как у других обычных цветовых форм (объединенных у Петерсена и Трехерна под общими названиями f. rubiginosa для самцов и f. hyalosuperficies для самок) эти части тела окрашены в различные оттенки бурого и серого цветов (рис. 1в, г). Кроме того, у самцов зеленой травянки, относящихся к «зеленой» форме, в отличие от самцов других форм этого вида, зеленый цвет имеют также темя, затылок, диск переднеспинки и бедра задних ног (у самцов, относящихся к другим известным нам формам, эти части тела буровато-серые).
Встречаемость «зеленой» формы в разных частях ареала этого вида ранее изучалась на территории Швеции и Норвегии (Petersen, Treherne, 1949). Подобных данных для территории России до сих пор не публиковалось.
Настоящая работа обобщает данные о частоте встречаемости «зеленой формы» зеленой травянки, полученные автором в 2015 и 2017 гг. в ходе полевых учетов на территории Псковской и Новгородской областей.

Рис. 1. Цветовые формы Omocestus viridulus: a – самец f. viridis (фото © Julius Rückert, CC BY-SA 3.0); б – самка f. viridis (фото © Gilles San Martin, CC BY-SA 3.0); в – самец f. rubiginosa (фото © Gilles San Martin, CC BY-SA 2.0); г – самка f. hyalosuperficies (фото © G.-U. Tolkiehn, CC BY 2.5)
Fig. 1. Color variants of Omocestus viridulus: a – male, f. viridis (© Julius Rückert, CC BY-SA 3.0); б – female, f. viridis (© Gilles San Martin, CC BY-SA 3.0); в – male, f. rubiginosa (© Gilles San Martin, CC BY-SA 2.0); г – female, f. hyalosuperficies (© G.-U. Tolkiehn, CC BY 2.5)
Материалы
Учет проводился летом 2015 и 2017 гг. на территории Новгородской и Псковской областей, в 10 географических точках (рис. 2), на мезофитных и гигрофитных лугах. Информация о точках учета и о количестве учтенных особей приведена в табл. 1.
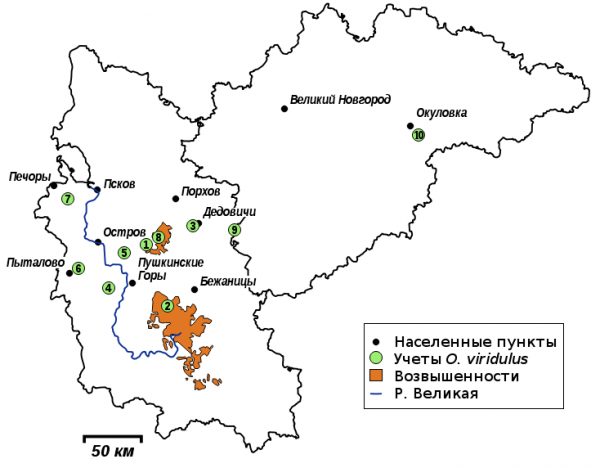
Рис. 2. Точки учета O. viridulus в Псковской и Новгородской областях. Номера точек – как в табл. 1
Fig. 2. Record points of O. viridulus in Pskov and Novgorod provinces. The points are numbered as in Table 1
Таблица 1. Характеристика мест учета зеленых травянок
| № п/п | Географическая точка | Координаты | Дата учета (ч.м.г.) | Учтено особей (♂/♀) |
| 1 | Окр. д. Спиры Островского р-на Псковской обл. | N57°31', E29°03' | 09.07.17 | 51 / 56 |
| 2 | Окр. д. Зубково Бежаницкого р-на Псковской обл. | N56°50', E29°26' | 10.07.17 | 50 / 55 |
| 3 | Окр. д. Хмелевицы Дедовичского р-на Псковской обл. | N57°27', E29°50' | 13.07.17 | 21 / 17 |
| 4 | Окр. с. Велье Пушкиногорского р-на Псковской обл. | N56°58', E28°34' | 18.07.17 | 39 / 48 |
| 5 | Окр. д. Астратово Островского р-на Псковской обл. | N57°16', E28°48' | 27.07.15 | 44 / 50 |
| 04.08.17 | 53 / 53 | |||
| 6 | Окр. д. Захаркино на территории Островского и Пыталовского р-нов Псковской обл. | N57°07', E28°05' | 21.07.17 | 54 / 50 |
| 7 | Окр. с. Изборск Печорского р-на Псковской обл. | N57°42', E28°53' | 23.07.17 | 66 / 50 |
| 8 | Окр. д. Пожни Порховского р-на Псковской обл. | N57°24', E29°18' | 25.07.17 | 50 / 53 |
| 9 | Окр. д. Алексино Дедовичского р-на Псковской обл. у границы с Поддорским районом Новгородской обл. | N57°30', E30°29' | 28.07.17 | 23 / 19 |
| 10 | Окр. д. Яблоновка Окуловского р-на Новгородской обл. | N58°17', E33°25' | 05.07.15 | 130 / 134 |
Методы
Подсчет саранчовых осуществлялся без отлова, на непересекающихся трансектах произвольной длины и формы, с расстоянием между параллельными участками не менее 1.5 м. Учитывались только имаго и личинки последнего возраста. Протоколировались пол и принадлежность или непринадлежность обнаруженных особей к «зеленой» форме.
Полученные данные подвергались статистической обработке с использованием компьютерных программ: PAST 3.16 (Hammer et al., 2001) для вычисления значений критерия Шапиро – Уилка и парного критерия Стьюдента; специально разработанная нами программа для вычисления значения точного критерия Фишера (алгоритм: Гланц, 1999). 95%-ные доверительные пределы рассчитывались с использованием фи-преобразования долей (Ивантер, Коросов, 2010).
Результаты
Общие закономерности. Ни в одной из исследованных нами географических точек, независимо от года сбора материала, «зеленая» форма не встречалась часто. Во всех точках, расположенных на территории Псковской области, такие особи были представлены в малом количестве, а в единственной точке, расположенной в Новгородской области, они отсутствовали полностью. Подавляющее большинство самок, отличных от «зеленой» формы, имело зеленый верх (включая темя, затылок, диск переднеспинки и видимую сверху часть надкрылий) и незеленые лицо, щеки, боковые лопасти переднеспинки и плейриты средне- и заднегруди, в целом соответствуя форме f. hyalosuperficies, как она описана у И. А. Рубцова (Rubtzov, 1935). При этом, однако, эти незеленые части тела у самок были весьма вариабельны по окраске (которая бывала серой, желтовато-бурой, охристой и т. п.) и степени выраженности расчленяющего рисунка (рис. 1в, г). Очень редко встречались также самки, у которых вместо зеленого верх тела был окрашен в сероватый цвет (f. rubiginosa по терминологии И. А. Рубцова): в Новгородской области (Яблоновка, учеты 2015 г.) из 132 самок таких оказалось всего 2, в Псковской же области во время учетов такие самки вообще не были встречены ни разу, и только вне учетов в 2017 г. близ дер. Астратово была обнаружена одна такая особь. В отличие от самок, все самцы зеленой травянки, не относящиеся к «зеленой» форме, во всех исследованных географических точках не имели зеленого цвета сверху и в целом соответствовали форме f. rubiginosa, варьируя при этом по степени выраженности расчленяющего рисунка.
Встречаемость «зеленой» формы среди самцов и самок. Для проверки гипотезы об отсутствии различий между представленностью «зеленой» формы у самцов и у самок был использован парный критерий Стьюдента, при этом каждая пара включала доли «зеленой» формы для самцов и для самок, относящейся к одной и той же географической точке и к одному и тому же году учета. Данные предварительно проверялись при помощи критерия Шапиро – Уилка и были признаны достаточно близкими к нормально распределенным (p > 0.05). Согласно парному критерию Стьюдента, представленность «зеленой» формы у самцов и самок с высокой вероятностью одинакова (p > 0.95).
Представленность «зеленой» формы в разные годы. Имеющиеся в распоряжении автора данные позволяют сравнить только представленность «зеленой» формы в одной географической точке (дер. Астратово) в 2015 и 2017 гг. Основываясь на приведенных выше результатах сравнения представленности этой формы у самцов и самок, мы объединили данные по обоим полам. Согласно точному критерию Фишера, представленность «зеленой» формы оказалась одинаковой с высокой вероятностью (p > 0.99).
Географические различия в представленности «зеленой» формы. В пределах изученной части Псковской области (точки 1–9) «зеленая» форма везде выявлена среди обоих полов и составляет у них от 4.8 (95%-ные доверительные пределы 0.0–18.5) до 11.2 % (4.4–20.7) ото всех учтенных особей. Сходная картина выявляется при отдельном рассмотрении особей мужского и женского пола: соответственно от 4.3 (0.0–16.2) до 12.1 (5.4–21.0) и от 5.3 (0.0–19.4) до 12.5 % (5.5–23.3) ото всех учтенных особей (рис. 3). Среди 130 самцов и 134 самок, учтенных в Новгородской области (точка 10), к «зеленой» форме не относилась ни одна особь.
Сравнение представленности «зеленой» формы в разных исследованных точках (с использованием точного критерия Фишера) показало следующее. С одной стороны, различия между травянками из разных точек Псковской области во всех случаях статистически незначимы (p > 0.05). С другой стороны, во всех исследованных точках, находящихся на территории Псковской области, доля зеленых особей статистически значимо превышает таковую в точке, относящейся к Новгородской области (табл. 2).
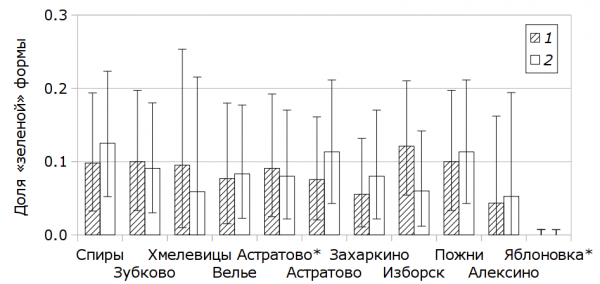
Рис. 3. Представленность «зеленой» формы O. viridulus в разных точках сбора. 1 – самцы, 2 – самки. Данные, помеченные звездочкой, относятся к 2015 г., остальные – к 2017 г. Приведены 95%-ные доверительные пределы
Fig. 3. Rate of the “green” colouration of O. viridulus in different localities. 1 – males, 2 – females. The data marked with an asterisk refer to the year 2015, the other data refer to 2017. 95 % confidence limits are presented
Таблица 2. Попарное сравнение доли «зеленой» формы у зеленых травянок
| Спиры | Зубково | Хмелевицы | Велье | Астратово 2015 | Астратово 2017 | Захаркино | Изборск | Пожни | Алексино | Яблоновка | |
| Спиры | – | ||||||||||
| Зубково | 0.82 | – | |||||||||
| Хмелевицы | 0.76 | 1.00 | – | ||||||||
| Велье | 0.63 | 0.80 | 1.00 | – | |||||||
| Астратово 2015 | 0.64 | 1.00 | 1.00 | 1.00 | – | ||||||
| Астратово 2017 | 0.82 | 1.00 | 1.00 | 0.80 | 1.00 | – | |||||
| Захаркино | 0.34 | 0.61 | 0.73 | 0.79 | 0.79 | 0.61 | – | ||||
| Изборск | 0.83 | 1.00 | 1.00 | 0.81 | 1.00 | 1.00 | 0.62 | – | |||
| Пожни | 1.00 | 0.82 | 0.76 | 0.62 | 0.64 | 0.82 | 0.34 | 0.82 | – | ||
| Алексино | 0.35 | 0.51 | 0.66 | 0.72 | 0.72 | 0.51 | 1.00 | 0.52 | 0.35 | – | |
| Яблоновка | 0.00* | 0.00* | 0.00* | 0.00* | 0.00* | 0.00* | 0.00* | 0.00* | 0.00* | 0.02* | – |
Примечание. Приведены уровни значимости. Звездочкой обозначены результаты сравнений, свидетельствующие о статистически значимых различиях.
Обсуждение
Полученные результаты представляют интерес с разных точек зрения. Во-первых, близкая доля представителей «зеленой» формы у самцов и самок зеленой травянки, при допущении о наследственной природе цветовых форм у этого вида (вполне правдоподобном с учетом того, что к настоящему времени известно о наследовании окраски у Gomphocerini), может быть интерпретирована как свидетельство аутосомной локализации соответствующего гена (или генов). С учетом того, что подавляющее большинство особей, не относящихся к «зеленой» форме, соответствовало форме f. hyalosuperficies в случае самок и форме f. rubiginosa у самцов, можно предположить, что эти два фенотипа соответствуют одному и тому же аллелю, по-разному проявляющемуся в зависимости от пола (при этом вполне вероятно, что аллели, соответствующие у самок двум разным формам, f. hyalosuperficies и f. rubiginosa, у самцов фенотипически проявляются одинаково).
В отношении встречаемости «зеленой» формы у O. viridulus особый интерес представляет сопоставление данных, полученных нами для 10 точек, расположенных на территории Северо-Запада России в 2015 и 2017 гг., с данными, имеющимися для Швеции и Норвегии на момент середины прошлого века (Petersen, Treherne, 1949). Согласно Петерсену и Трехерну, в 5 исследованных ими географических точках (расположенных южнее 62-й параллели) «зеленая» форма являлась преобладающей, составляя от 64.8 до 75.2 % ото всех особей. Еще в одной точке, исследованной этими же авторами на территории Швеции, но расположенной севернее (N 63°10'), доля особей «зеленой» формы оказалась хотя и меньше половины, но все равно весьма высокой – 41.2 %. Сравнение данных Петерсена и Трехерна с нашими (с использованием точного критерия Фишера) показало, что представленность «зеленой» формы O. viridulus в каждой из изученных ими географических точек значимо выше, чем в каждой из изученных нами (p < 0.001).
К сожалению, без проведения дополнительных исследований соотношений между цветовыми формами O. viridulus на территории Скандинавии невозможно судить, чем вызваны выявленные нами различия между зелеными травянками, с одной стороны, Швеции и Норвегии и, с другой стороны, Северо-Запада России. Речь может идти как о географических особенностях соответствующих популяций, так и об изменении соотношения между цветовыми формами, произошедшем за более чем полувековой срок. В то же время выявленные нами различия между представителями этого вида из Псковской и Новгородской областей безусловно носят именно географический характер. При этом особо следует заметить, что ни расстояния между точками учета, ни реки, ни перепады высот не оказали сколь-либо заметного влияния на соотношение между цветовыми формами у зеленых травянок в разных точках Псковской области. Так, расстояние между крайней западной («Изборск») и крайней восточной («Алексино») точками учета в Псковской области составляет около 150 км, точки «Изборск», «Захаркино» и «Велье» отделены от прочих р. Великой, а точки «Зубково» и «Пожни», в отличие от прочих, располагаются на возвышенностях: первая – на Бежаницкой, вторая – на Судомской. При этом представленность «зеленой» формы во всех этих точках оказалась примерно такой же, как и в остальных на территории Псковской области. Таким образом, либо на исследованной нами части Псковской области обитает одна большая популяция зеленой травянки, поддерживающая единство своего генофонда несмотря на существование потенциальных географических барьеров, либо же какие-то внешние факторы (например, селективное выедание особей с определенной окраской хищниками) поддерживают близкое соотношение между цветовыми формами у нескольких соседствующих популяций. В то же время зеленые травянки из точки «Яблоновка», расположенной на востоке Новгородской области, характеризуются полным отсутствием у них «зеленой» формы, статистически значимо отличаясь в этом отношении от травянок Псковской области. Можно полагать, что в данном случае речь идет о другой популяции этого вида, западная граница ареала которой находится на территории Новгородской области и проходит западнее Валдайской возвышенности или непосредственно по ней. Для определения этой границы, а также для проверки гипотезы о популяционном единстве зеленых травянок средней части Псковской области необходимы дальнейшие исследования (учеты саранчовых в западной и центральной частях Новгородской области и на сопредельных территориях, молекулярно-генетические исследования и т. п.).
С точки зрения возможного адаптивного значения соотношений между цветовыми формами у O. viridulus заслуживает внимания малая доля или полное отсутствие у этого вида на всей исследованной нами территории как «зеленой» формы (f. viridis), так и (у самок) формы, полностью лишенной зеленых элементов окраски (f. rubiginosa). Следует заметить, что данная закономерность относится именно к зеленой травянке и не распространяется на ряд симпатрических видов из той же трибы Gomphocerini s. l. Так, на территории в радиусе не более 15 км от точки «Астратово» преобладают полностью или по большей части окрашенные в зеленый цвет особи короткокрылого конька Chorthippus parallelus (соответствующие формам f. viridis, f. hyalolateralis и f. porphyrica в терминологии Рубцова (Rubtzov, 1935)), в то время как почти лишенные зеленых элементов окраски особи этого вида (соответствующие форме f. rubiginosa в смысле Рубцова) чрезвычайно редки (Озерский, 2014). Обитающие там же луговые коньки Chorthippus dorsatus представлены преимущественно формами f. hyalosuperficies и f. rubiginosa в соотношении, близком к 1:1 (Озерский, 2014). При этом зеленая травянка и оба вида коньков не только симпатричны в зоогеографическом смысле, но и нередко населяют на указанной территории одни и те же стации. Точно так же, совершенно по-разному, представлены цветовые формы у зеленой травянки и у северного конька Chorthippus montanus в точке «Яблоновка» Новгородской области, учитывавшихся на одном и том же лугу: первый вид представлен там формами f. rubiginosa (все самцы и единичные самки) и f. hyalosuperficies (подавляющее большинство самок), второй – формами f. viridis, f. hyalolateralis и f. porphyrica (Озерский, 2014). Такие различия между видами могут быть объяснены как случайностью (последствиями генетического дрейфа, независимо протекавшего у разных видов), так и различиями в их специализации к разным микростациям (например, выбором разных ярусов травостоя и участков с разным проективным покрытием). В данном случае также целесообразно проведение дополнительных (прежде всего этологических) исследований.
Заключение
Обобщая, можно сделать следующие выводы:
1. На всей изученной нами территории «зеленая» форма у O. viridulus встречается редко (точки, расположенные в Псковской области) или не встречается вовсе (точка в Новгородской области).
2. В пределах изученной нами территории Псковской области частота встречаемости «зеленой» формы O. viridulus примерно одинакова (несмотря на большую удаленность некоторых точек друг от друга, различия в рельефе и существование потенциального географического барьера в виде р. Великой), однако статистически значимо отлична от таковой в исследованной нами точке Новгородской области.
3. На всей изученной нами территории доля особей O. viridulus «зеленой» формы существенно (статистически значимо) меньше, чем наблюдавшаяся в середине XX века в Швеции и Норвегии.
4. «Зеленая» форма на территории Псковской области встречается с примерно одной и той же частотой у самцов и у самок O. viridulus, в то время как встречаемость остальных форм у самцов и у самок весьма различна.
Библиография
Бей-Биенко Г. Я., Мищенко Л. Л. Саранчовые фауны СССР и сопредельных стран. Ч. 1 . М.; Л.: Изд-во АН СССР, 1951. 378 с.
Бенедиктов А. А. К систематике палеарктических представителей саранчовых трибы Bryodemini (Orthoptera, Acrididae) // Зоологический журнал. 1998. Т. 77. № 7. С. 788–799.
Бенедиктов А. А. Изменчивость узора на крыльях саранчовых трибы Bryodemini Bey-Bienko (Orthoptera: Acrididae) // Труды Зоологического института РАН. 2016. Т. 320. № 4. C. 467–472.
Воронцовский П. А. К вопросу о гомологических рядах цветовой изменчивости у саранчовых // Известия Оренбургской станции защиты растений. 1928. Вып. 1. С. 27–39.
Гланц С. Медико-биологическая статистика . М.: Практика, 1999. 459 с.
Горохов А. В. Жизненные формы сверчковых (Orthoptera, Grylloidea) Дальнего Востока СССР // Научные доклады высшей школы. Биол. науки. 1983. № 1. С. 49–56.
Ивантер Э. В., Коросов А. В. Элементарная биометрия: Учеб. пособие . Петрозаводск: Изд-во ПетрГУ, 2010. 104 с.
Озерский П. В. Метафенотип популяции как структурно-функциональное отражение ее экологической ниши // Функциональная морфология, экология и жизненные циклы животных. Вып. 10. СПб.: Изд-во РГПУ им. А. И. Герцена, 2010. С. 15–29.
Озерский П. В. О перспективах использования изменчивости окраски в классификации жизненных форм прямокрылых насекомых // Функциональная морфология, экология и жизненные циклы животных. Вып. 12. СПб.: Тесса, 2012. С. 11–17.
Озерский П. В. К вопросу об изменчивости окраски у прямокрылых (Insecta, Orthoptera) разных жизненных форм // Функциональная морфология, экология и жизненные циклы животных. 2014. Т. 14. № 2. С. 42–52.
Озерский П. В., Боброва Т. В., Кузнецова Л. С. К вопросу о функциональном значении угла наклона лба у саранчовых разных жизненных форм: постановка проблемы и проверка одной из гипотез // Известия Российского государственного педагогического университета имени А. И. Герцена. 2011. № 41. С. 124–131.
Правдин Ф. Н. Экологическая география насекомых Средней Азии. Ортоптероиды . М.: Наука, 1978. 272 с.
Присный А. В. Функциональная дифференциация элементов окраски у саранчовых // Ландшафтная экология насекомых. Новосибирск: Наука, 1988. С. 34–47.
Стебаев И. В. Жизненные формы и половой диморфизм саранчовых Тувы и Юго-Западного Алтая // Зоологический журнал. 1970. Т. 49. № 3. С. 325–338.
Стебаев И. В. Морфоадаптогенез саранчовых и система их жизненных форм // Журнал общей биологии. 1987. Т. 48. № 3. С. 626–639.
Стебаев И. В. Закономерности сочетания сигнальной и покровительственной окраски саранчовых // Зоологический журнал. 1990. Т. 69. № 6. С. 58–69.
Стебаев И. В., Омельченко Л. В. Общие особенности морфоадаптационных типов, или жизненных форм, саранчовых Южной Сибири и сопредельных территорий // Вопросы экологии. Поведение и экология насекомых, связанных с агробиогеоценозами. Новосибирск: Изд-во НГУ, 1981. С. 13–39.
Черняховский М. Е. Морфо-функциональные особенности жизненных форм саранчовых // Ученые записки МГПИ им. В. И. Ленина. 1970. № 394. С 47–63.
Яблоков А. В. Популяционная биология . М.: Высшая школа, 1987. 303 с.
Gotham S., Song H. Non-swarming grasshoppers exhibit density-dependent phenotypic plasticity reminiscent of swarming locusts // J. Insect Physiol. 2013. Vol. 59. P. 1151–1159.
Hammer Ø., Harper D. A. T., Ryan P. D. Past: paleontological statistics software. Package for education and data analysis // Palaeontologia Electronica. 2001. Vol. 4. Iss. 1. Art. 4. 9 p. URL: http://palaeo-electronica.org/2001_1/past/past.pdf (дата обращения: 03.10.2017).
Levita B. Étude de l'homochromie chez Oedipoda coerulescens L. (Acridien, Orthoptère) // Bulletin Biologique de la France et de la Belgique. 1970. Vol. 104. № 2. P. 149–213.
Moreteau B. Fonction chromatrope de la pars intercerebralis chez l’acridien Oedipoda caerulescens // J. Insect Physiol. 1975. Vol. 21. № 7. P. 1407–1413.
Morse A. P. Further researches on North American Acrididae. Carnegie Inst. Washington publication № 68. Washington: Wilkens-Sheiry, 1907. 54 p.
Petersen B., Treherne J. E. On the distribution of colour forms in Scandinavian Omocestus viridulus L. // Oikos. 1949. Vol. 1. № 2. P. 175–183.
Rowell C. H. F. The variable coloration of the acridoid grasshoppers // Advances in Insect Physiology. 1972. Vol. 8. P. 145–198.
Rubtzov I. A. Phase variation in non-swarming grasshoppers // Bull. Ent. Res. 1935. Vol. 26. № 4. P. 499–520.
Uvarov B. P. A revision of the genus Locusta L. (=Pachytilus Fieb.), with a new theory as to the periodicity and migrations of locusts // Bull. Ent. Res. 1921. Vol. 12. № 2. P. 135–163.
Yerushalmi Y., Pener M. P. Age-dependent response of adults of a homochrome grasshopper, Oedipoda miniata, to the dark-colour-inducing neurohormone (DCIN) of locusts // Physiological Entomology. 2002. Vol. 27. № 2. P. 165–169.

 © 2011 - 2026
© 2011 - 2026