



Введение
Свет является экологическим фактором, влияющим на физиологические функции млекопитающих. Кодирование информации о световом режиме осуществляется с помощью эпифизарного гормона – мелатонина. Влияя на функциональную активность гипоталамо-гипофизарно-надпочечниковой и половой систем, мелатонин участвует в регуляции циркадианных и сезонных ритмов. Однако в большинстве случаев изучается ограниченное количество физиологических параметров, хотя в организме функционирование всех систем связано между собой. В настоящем исследовании проверялась гипотеза о том, что торможение с помощью постоянного освещения секреции эпифизом мелатонина приводит к влиянию на антиоксидантную систему (АОС), вызывающему перестройки иммуннореактивности, ускорение полового созревания и уменьшение продолжительности жизни.
Материалы
Эксперименты были проведены на лабораторных крысах линии ЛИО (Ленинградский институт онкологии), полученных из НИИ онкологии им. проф. Н. Н. Петрова. Крыс содержали в стандартных помещениях вивария Петрозаводского государственного университета площадью 25 м2 в клетках размером 44 х 25 х 62 см при температуре 22 ± 2 ºС. Все животные получали стандартный готовый лабораторный корм и воду без ограничений.
Методы
В первой серии экспериментов беременные самки содержались в условиях стандартного (12 ч. свет 750 лк /12 ч. темнота; LD), естественного освещения Республики Карелия (NL), в данном случае учитывались особенности годовой фотопериодичности Северо-Запада России, и постоянного освещения (750 лк; LL) (рис. 1). Потомство от самок первой группы с момента рождения разделили на 2 подгруппы и содержали при стандартном (LD/LD) и постоянном (LD/LL) освещении. Крысы от самок второй и третьей групп после рождения оставались в тех же световых условиях, при которых находились их матери – соответственно естественном (NL/NL) и постоянном освещении (LL/LL). В возрасте 3 месяцев у животных из каждой подгруппы отбирали образцы крови и тканей для последующего анализа. Во второй серии исследований крыс в 25-дневном возрасте рандомизировано разделили на 4 группы. Первая группа крыс находилась в условиях стандартного фиксированного режима освещения (LD), вторая – в условиях естественного освещения (NL). Третья группа крыс содержалась при круглосуточном постоянном освещении (LL). Животные четвертой группы находились в условиях световой депривации (DD), при этом освещенность в помещении составляла 0–0,5 лк на 1 м2 площади.
Исследовали состояние антиоксидантной системы (АОС), состав лейкоцитарной формулы, продолжительность жизни и скорость полового созревания. Лейкоцитарную формулу и количество лейкоцитов в крови определяли общепринятым способом с использованием компьютерной системы анализа изображений с цветной цифровой видеокамерой и программным обеспечением “Видеотест”.
Числовые данные обрабатывали с использованием общепринятых методов вариационной статистики, при сравнении групп применяли непараметрические критерии, оценку различий в динамике полового созревания и смертности проводили с использованием критерия хи-квадрат (Коросов, Горбач, 2007).
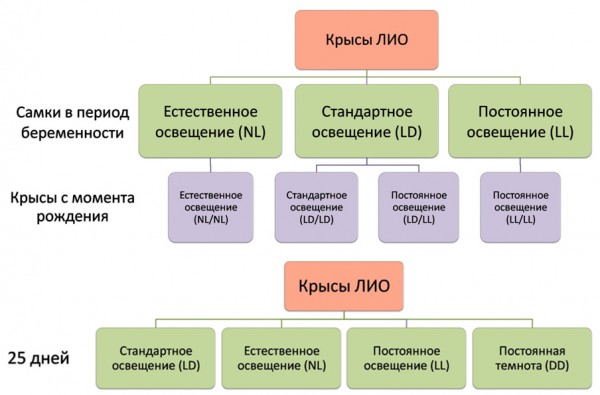
Рис. 1. Схемы экспериментов.
Условные обозначения: NL/NL, LD/LD, LL/LL – беременные самки и их потомство с момента рождения содержались соответственно при естественном, стандартном (12/12) и постоянном освещении, LD/LL – беременные самки содержались при стандартном освещении, а их потомство при постоянном освещении с момента рождения, LD – стандартное освещение, NL – естественное освещение, LL – постоянное освещение, DD – постоянная темнота
Fig. 1. Design of experiments.
NL/NL, LD/LD, LL/LL – pregnant female rats and their pups lives from birth in natural, standard (12/12) and constant light conditions accordingly, LD/LL – pregnant female rats lives under standard light condition and their pups from birth lives in constant light condition, LD – standard light condition, NL – natural light condition, LL – constant light condition, DD – constant darkness
Результаты
Влияние постоянного и естественного освещения зависело от момента начала воздействия измененного светового режима. Установлено, что постоянное освещение матерей в период беременности или потомства с момента рождения приводит к однонаправленным изменениям активности антиоксидантных ферментов у потомства, хотя их выраженность различается (рис. 2). При этом выявлено рассогласование в активности сопряженных антиоксидантных ферментов (АОФ) – супероксиддисмутазы (СОД) и каталазы. При естественном освещении направленность изменений АОФ была противоположна той, которая наблюдалась при постоянном освещении. Это могло быть связано с тем, что исследуемые животные находились в условиях естественного освещения, характерного для осеннего периода (сентябрь – декабрь). Одновременно с этим отмечено существенное снижение в крови уровня сегментоядерных нейтрофилов и увеличение доли лимфоцитов в группе, находившейся при постоянном освещении с момента внутриутробного развития, по сравнению с крысами, содержавшимися при регулярно чередующемся и естественном освещении. Нахождение животных в период внутриутробного развития и с момента рождения при постоянном освещении практически не отразилось на половом созревании самцов (рис. 3). В то же время динамика полового созревания самцов крыс, находившихся в условиях естественного освещения с момента внутриутробного развития, существенно отличалась от таковой у содержавшихся в регулярно чередующейся 12-часовой смене света и темноты (рис. 4). При этом у крыс, находившихся при удлиняющемся фотопериоде, наблюдалось ускорение, а при сокращающемся, наоборот,– замедление полового созревания.
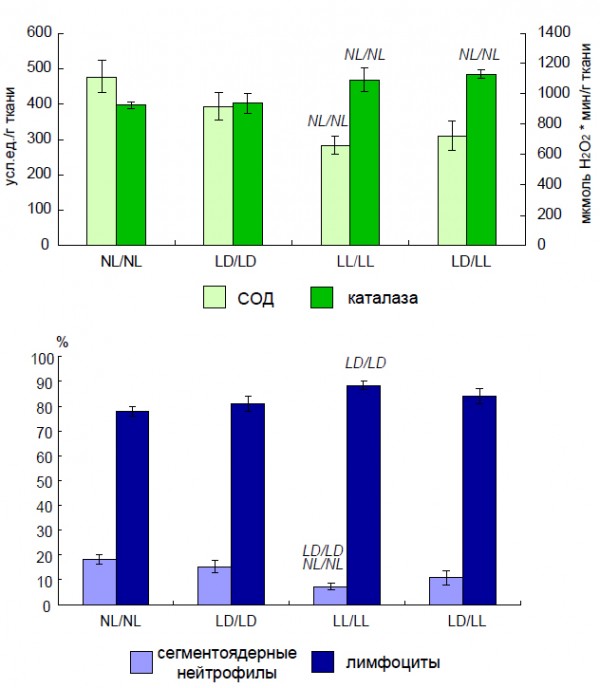
Рис. 2. Влияние режимов освещения на активность АОФ и показатели крови у 3-месячных крыс (M ± m). Условные обозначения: NL/NL, LD/LD, LL/LL – беременные самки и их потомство с момента рождения содержались соответственно при естественном, стандартном и постоянном освещении, LD/LL – беременные самки содержались при стандартном освещении, а их потомство при постоянном освещении с момента рождения; NL/NL, LD/LD – изменения достоверны по сравнению животными, которые содержались соответственно в группе NL/NL и LD/LD (p < 0,05)
Fig. 2. Effect of various light conditions on the antioxidant enzymes activity and blood’s parameters in 3 months old rats (M ± m). NL/NL, LD/LD, LL/LL – pregnant female rats and their pups lives from birth in natural, standard (12/12) and constant light conditions accordingly, LD/LL – pregnant female rats lives under standard light condition and their pups from birth lives in constant light condition; NL/NL, LD/LD – the difference with the relevant parameter in the group NL/NL and LD/LD is significant, p < 0,05
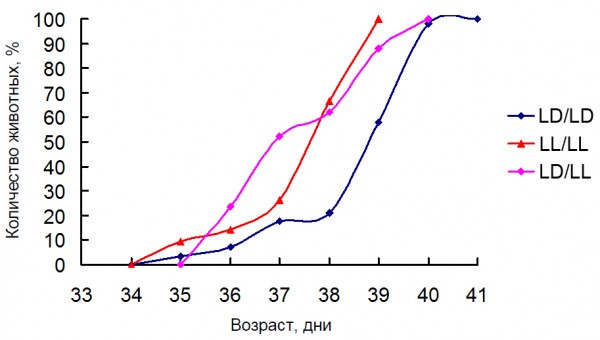
Рис. 3. Влияние постоянного освещения на динамику полового созревания самцов крыс. Условные обозначения: LD/LD, LL/LL – беременные самки и их потомство с момента рождения содержались соответственно при стандартном и постоянном освещении, LD/LL – беременные самки содержались при стандартном освещении, а их потомство при постоянном освещении с момента рождения
Fig. 3. Effect of constant light on the male rat’s pubescence dynamic. LD/LD, LL/LL – pregnant female rats and their pups lives from birth in standard (12/12) and constant light conditions accordingly, LD/LL – pregnant female rats lives under standard light condition and their pups from birth lives in constant light condition
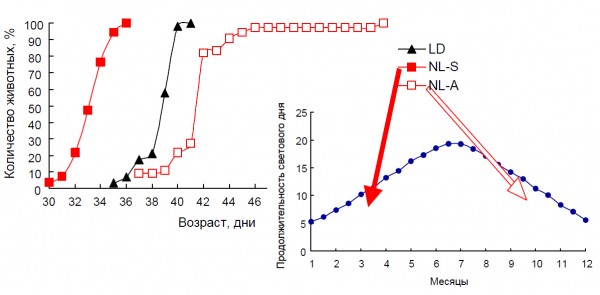
Рис. 4. Влияние естественного освещения на динамику полового созревания самцов крыс и продолжительность светового дня в условиях Карелии. Условные обозначения: LD – стандартное освещение; NL-S – естественное освещение (весна); NL-A – естественное освещение (осень)
Fig. 4. Effect of natural light on the dynamic of male rat’s pubescence and day length inKarelia. LD – standard light condition, NL-S – natural light condition (spring), NL-A – natural light condition (autumn)
Крысы, содержавшиеся при постоянном освещении с 25-дневного возраста, имели максимальное рассогласование в работе АОС (рис. 5) и наиболее низкие показатели продолжительности жизни (табл. 1). Продемонстрированы связанные с этим синхронные изменения ряда других физиолого-биохимических показателей. Так, «старение» иммунитета (соотношение сегментоядерных нейтрофилов и лимфоцитов) происходило наиболее интенсивно у животных, содержавшихся при постоянном освещении – относительное содержание клеток этих типов уже у шестимесячных животных достигало значений, характерных для взрослых и стареющих 12–18 месячных крыс, находившихся в регулярно чередующемся световом режиме (рис. 6). Соотношение гетерофилы/лимфоциты является простым и удобным показателем для оценки функциональной нагрузки на организм и стресса различного происхождения (Tabuchi et al., 2011). Постоянное освещение ускоряло половое созревание самцов крыс, а световая депривация замедляла его (рис. 7). Эффекты естественного освещения выражались в усилении сезонных колебаний большинства изученных показателей и совпадали в летний период по направленности с постоянным освещением.
Таблица 1. Влияние различных режимов освещения на продолжительность жизни самцов крыс
Table 1. Effect of the exposure to various light regimens on survival in male rats
| Световой режим | ||||
| Стандартное освещение (LD) | Естественное освещение (NL) | Постоянное освещение (LL) | Световая депривация (DD) | |
| Количество крыс, шт. | 57 | 50 | 50 | 51 |
| СПЖ, сут. | 644 ± 34,0 | 613 ± 32,9 | 580 ± 35,5 | 652 ± 32,5 |
| МПЖ, сут. | 1045 | 1046 | 1005 | 1017 |
| СПЖ последних 10% крыс, сут. | 999 ± 11,5 | 972 ± 22,7 | 987 ± 13,0 | 987 ± 13,0 |
Примечание. СПЖ – средняя продолжительность жизни, МПЖ – максимальная продолжительность жизни.
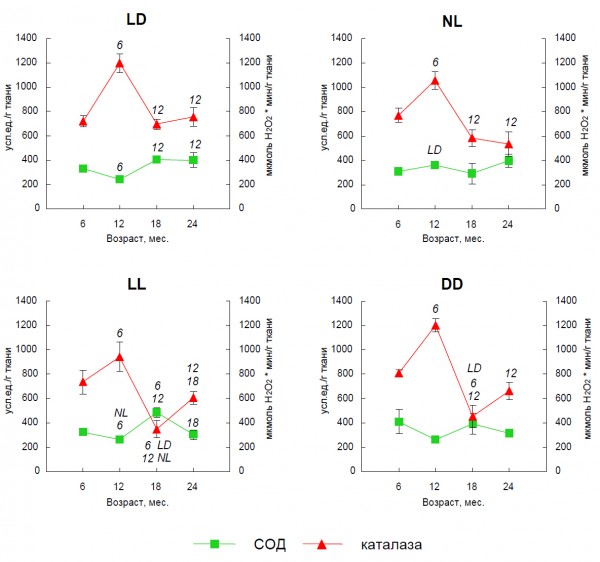
Рис. 5. Влияние световых режимов на активность АОФ в печени крыс разного возраста (M ± m). Условные обозначения: LD – стандартное освещение (12/12), NL – естественное освещение, LL – постоянное освещение, DD – постоянная темнота; LD, NL – изменения достоверны по сравнению с животными, которые содержались при стандартном и естественном освещении, соответственно (p < 0,05); 6, 12, 18 – изменения достоверны по сравнению с 6-, 12- и 18-месячными животными, содержавшимися при одинаковых световых режимах (p < 0,05)
Fig. 5. Effect of various light conditions on age-related dynamics of the antioxidant enzymes activity in rats liver (M ± m). LD – standard light condition (12/12), NL – natural light condition, LL – constant light condition, DD – constant darkness; LD, NL – the difference with the relevant parameter in the group LD and NL is significant, p < 0,05; 6, 12, 18 – the difference with the relevant parameter of 6, 12 and 18 month old rats is significant, p < 0,05
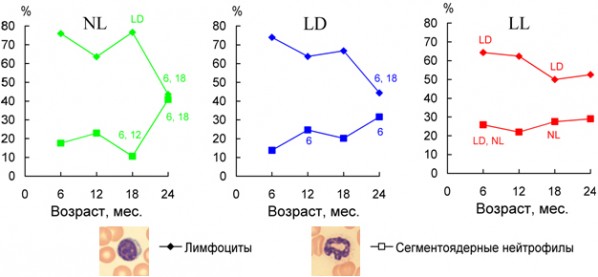
Рис. 6. Изменение количества лимфоцитов и нейтрофилов в периферической крови крыс при различных режимах освещения. Условные обозначения: LD – стандартное освещение (12/12), NL – естественное освещение, LL – постоянное освещение; LD, NL – изменения достоверны по сравнению с животными, которые содержались соответственно при стандартном и естественном освещении, (p < 0,05); 6, 12, 18 – изменения достоверны по сравнению с 6-, 12- и 18-месячными животными, содержавшимися при одинаковых световых режимах (p < 0,05).
Fig. 6. Changes of neutrophils and lymphocytes content in rat peripheral blood under different light condition. LD – standard light condition (12/12), NL – natural light condition, LL – constant light condition;LD, NL – the difference with the relevant parameter in the group LD and NL is significant, p < 0,05; 6, 12, 18 – the difference with the relevant parameter of 6, 12 and 18 month old rats is significant, p < 0,05
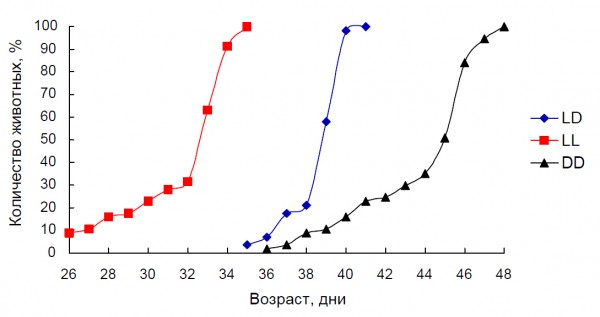
Рис. 7. Динамика полового созревания самцов крыс при различных световых режимах. Условные обозначения: LD – стандартное освещение (12/12), LL – постоянное освещение, DD – постоянная темнота
Fig. 7. Effect of the various light conditions on the dynamic of male rat’s pubescence. LD – standard light condition (12/12), LL – constant light condition, DD – constant darkness
Обсуждение
Большинство выявленных нами эффектов измененного светового режима (постоянное освещение) на такие физиологические функции, как состояние антиоксидантной системы, иммунная функция, скорость полового созревания и старения, достаточно хорошо изучены и связываются исследователями с синтезом пинеальной железой гормона мелатонина (Анисимов, 2003). Прежде всего, обращает на себя внимание различная чувствительность изученных систем к экзогенному подавлению синтеза мелатонина с помощью постоянного освещения и зависимость их реактивности от момента начала экспериментальных воздействий. Так, максимальный эффект на скорость полового созревания самцов выявлен при содержании животных в искусственных световых условиях с 25-дневного возраста, что, очевидно, обусловлено сроками становления функциональной активности эпифиза,– именно к этому периоду происходит становление дефинитивного уровня мелатонин-секретирующей функции (Calvo, 1984). Изменение активности АОФ отмечается и у крыс, подвергавшихся воздействию постоянного освещения в период внутриутробного развития. Такая реакция связана со способностью мелатонина проникать через плацентарный барьер (Arendt, 1998; Simonneaux, Ribelayga, 2003). Возможны как минимум два механизма действия мелатонина на антиоксидантную систему (Melchiorri et al., 1995; Pierreficge et al., 1993; Tan et al., 1994), включающих непосредственное улавливание активных форм кислорода и/или торможение их генерации в клетке и регуляцию активности антиоксидантных ферментов.
Мелатонин, синтезируемый пинеальной железой, играет существенную роль в регуляции полового созревания, репродуктивных циклов, стрессорной реакции и иммунного ответа (Maestroni et al., 1988; Maestroni, Conti, 1993; Guerrero, Reiter, 2002; Nelson, 2004; Esquifino et al., 2004; Maestroni, 2001, Reiter, 1993). В последние годы значительное внимание уделяется взаимодействию между пинеальной железой и иммунной системой (Guerrero, Reiter, 2002; Maestroni, 2001; Srinivasan et al., 2005; Moore et al., 2002). Особенно существенна роль мелатонина как иммуномодулятора при нарушениях иммунологического статуса, вызванных снижением эндогенного уровня гормона при функциональных и фармакологических воздействиях (Srinivasan et al., 2005; Moore et al., 2002). Уровень мелатонина положительно коррелирует как с гуморальным (Akbulut et al., 2001), так и с клеточным иммунитетом (Brennan et al., 2002; Champney et al., 1997). Введение экзогенного мелатонина в периоды, когда наблюдается снижение активности пинеальной железы (Nelson, Drazen, 1999; Haldar et al., 2001), приводит к усилению отдельных показателей иммунитета. Лимфоциты способны синтезировать мелатонин, что является дополнительным свидетельством участия мелатонина в регуляции иммунной системы (Carrillo-Vico et al., 2004). Уровень многих гормонов снижается с возрастом (гормон роста, эстроген, дегидроэпиандростерон и мелатонин), что играет важную роль в процессе старения иммунной системы (Arlt, Hewison, 2004). Следует особо отметить то, что с одной стороны, для синтеза половых стероидных гормонов необходимы активные формы кислорода, а с другой, метаболические превращения этих гормонов связаны с генерацией АФК. Таким образом, организм вынужден тонко регулировать уровень АФК, поскольку его значительное снижение из-за высокой активности АОФ тормозит процесс полового созревания млекопитающих и одновременно приводит к замедлению процесса старения.
Влияние естественного освещения на динамику полового созревания и старения крыс, состояние АОС и соотношение сегментоядерные нейтрофилы/лимфоциты отличалось как от постоянного освещения, так и от световой депривации. Воздействие естественного освещения на скорость полового созревания, как при увеличивающемся фотопериоде весной, так и при сокращающемся осенью выражено даже сильнее, чем при постоянном. Параллельно с этим происходит перестройка активности АОФ и состава лейкоформулы, что опосредованно сказывается на продолжительности жизни. Хорошо известен факт о различной продолжительности жизни осенних и весенних генераций грызунов (Назарова, Евсиков, 2011).
Существенным моментом является то, что в большинстве исследований по влиянию света внимание уделяется какой-либо одной физиологической функции. При этом не учитывается, что организм млекопитающего обладает ограниченным количеством доступных энергетических ресурсов (physiological trade-off), расходуемых в зависимости от окружающих условий только на осуществление отдельных функций, что неминуемо приводит к ограничению их использования на другие (French, 2009). Триггером, осуществляющим переключение потоков ресурсов, является эпифизарный гормон мелатонин.
Заключение
У самцов лабораторных крыс показана взаимосвязь между состоянием антиоксидантной системы, некоторыми параметрами иммунной системы, скоростью полового созревания и продолжительностью жизни (рис. 8). Изменение уровня секреции мелатонина с помощью постоянного и естественного освещения приводит к взаимосвязанным перестройкам в функционировании этих систем. Усиление активности антиоксидантной системы ведет к увеличению продолжительности жизни, но одновременно с этим – замедлению полового созревания животных. Изменение про/антиоксидантного баланса отражается и на составе иммунокомпетентных клеток в крови, для осуществления функции которых также необходимы активные формы кислорода.
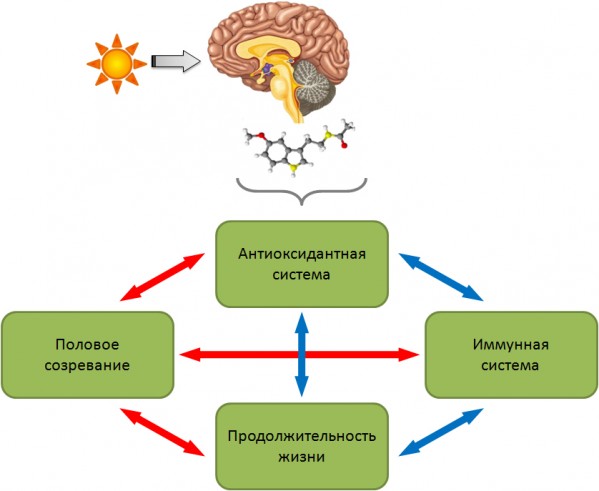
Рис. 8. Схема влияния света на различные физиологические параметры млекопитающих. Условные обозначения: красные стрелки – отрицательные корреляционные связи, синие – положительные
Fig. 8. Influences of light on different physiological parameters of mammals. Red arrow – negative connection, blue arrow – positive connection
Библиография
Анисимов В. Н. Молекулярные и физиологические механизмы старения. СПб.: Наука, 2003. 467 с.
Коросов А. В., Горбач В. В. Компьютерная обработка биологических данных: Метод. пособие. Петрозаводск: Изд-во ПетрГУ. 2007. 76 с.
Назарова Г. Г., Евсиков В. И. Эволюционная экология плодовитости животных: адаптивные возможности потомков предопределяются условиями их пренатального развития (на примере водяной полевки, Arvicola terrestris L.) // Вавиловский журнал генетики и селекции. 2011. Т. 15. № 3. С. 485–492.
Akbulut K. G., Gonul B., Akbulut H. The effects of melatonin on humoral immune responses of young and aged rats // Immunol. Invest. 2001 Vol. 30. № 1. P. 17–20.
Arendt J. Melatonin and the pineal gland: influence on mammalian seasonal and circadian physiology // J. Reprod. Fertility. 1998. Vol. 3. P. 13–22.
Arlt W., Hewison M. Hormones and immune function: implications of aging // Aging Cell. 2004. Vol. 3. P. 209–216.
Brennan C. P., Hendricks G. L., El-Sheikh T. M., Mashaly M. M. Melatonin and the enhancement of immune responses in immature male chickens // Poultry Sci. 2002. Vol. 81. P. 371–375.
Calvo J., Boya J. Postnatal evolution of the rat pineal gland: light microscopy // J. Anat. 1984. Vol. 138. № 1. P. 45–53.
Carrillo-Vico A., Calvo J. R., Abreu P., Lardone P. J., Garcia-Maurino S., Reiter R. J., Guerrero J. M. Evidence of melatonin synthesis by human lymphocytes and its physiological significance: possible role as intracrine, autocrine, and/or paracrine substance // FASEB J. 2004. Vol. 18. P. 537–539.
Champney T. H., Prado J., Youngblood T., Appel K., McMurray D. N. Immune responsiveness of splenocytes after chronic daily melatonin administration in male Syrian hamsters // Immunol. Lett. 1997. Vol. 58. № 2. P. 95–100.
Esquifino A. I., Pandi-Perumal S. R., Cardinali D. P. Circadian organization of the immune response: A role for melatonin // Clin. Appl. Immunol. Rev. 2004. Vol. 4. P. 423–433.
French S. S., Moore M. C., Demas G. E. Ecological immunology: The organism in context // Integrative and Comparative Biology. 2009. Vol. 49. № 3. P. 246–253.
Guerrero J. M., Reiter R. J. Melatonin immune system relationship // Curr. Top. Med. Chem. 2002. Vol. 2. P. 167–179.
Haldar C., Singh R., Guchhait P. Relationship between the annual rhythms in melatonin and immune system status in the tropical palm squirrel, Funambulus pennanti // Chronobiol. Int. 2001. Vol. 18. № 1. P. 61–69.
Maestroni G. J. The immunotherapeutic potential of melatonin // Expert Opin. Investig. Drugs. 2001. Vol. 10. P. 467–476.
Maestroni G. J. M., Conti A. Melatonin and the immune system. // Melatonin and the Pineal Gland – From Basic Science to Clinical Application. / Y. Touitou, J. Arendt, P. Pevet (Eds.). N.Y.: Elsevier, 1993. P. 295–302.
Maestroni G. J. M., Conti A., Pierpaoli W. Role of the pineal gland in immunity: circadian synthesis and release of melatonin modulates the antibody response and antagonizes the immunosuppressive effect of corticosterone // J. Neuroimmunol. 1986. Vol. 13. P. 19–30.
Melchiorri D., Reiter R. J., Attia A. M. et al. Potent protective effect of melatonin on in vivo paraquat-induced oxidative damage in rats // Life Sci. 1995. Vol. 56. P. 83–85.
Moore C. B., Siopes T. D., Steele C. T., Underwood H. Pineal melatonin secretion, but not ocular melatonin secretion, is sufficient to maintain normal immune responses in Japanese quail (Coturnix coturnix japonica) // Gen. Comp. Endocrinol. 2002. Vol. 126. P. 352–358.
Nelson R. J. Seasonal immune function and sickness responses // Trends Immunol. 2004. Vol. 25. P. 187–192.
Nelson R. J., Drazen D. L. Melatonin mediates seasonal adjustments in immune function // Reprod. Nutr. Dev. 1999. Vol. 39. P. 383–398.
Pierrefiche G., Topall G., Cauboin G., Henriet I., Laborit H. Antioxidant activity of melatonin in mice // Res. Commun. Chem. Pathol. Pharmacol. 1993. Vol. 80. № 2. P. 211–223.
Reiter R. J. The melatonin rhythm: Both a clock and a calendar // Experientia. 1993. Vol. 49. P. 654–664.
Simonneaux V., Ribelayga C. Generation of the Melatonin Endocrine Message in Mammals: A Review of the Complex Regulation of Melatonin Synthesis by Norepinephrine, Peptides, and Other Pineal Transmitters // Pharmacol. Rev. 2003. Vol. 55. P. 325–395.
Srinivasan V., Maestroni G. J., Cardinali D. P., Esquino A. I., Perumal S. R., Miller S. C. Melatonin, immune function and aging // Immun. Ageing. 2005. Vol. 29. P. 2–17.
Tabuchi T., Shimazaki J., Satani T., Nakachi T., Watanabe Y., Tabuchi T. The perioperative granulocyte/lymphocyte ratio is a clinically relevant marker of surgical stress in patients with colorectal cancer // Cytokine. 2011. Vol. 53. P. 243–248
Tan D. X., Reiter R. J., Chen L. D. et al. Both physiological and pharmacological levels of melatonin reduce DNA adduct formation induced by the carcinogen safrole // Carcinogenesis. 1994. Vol. 15. P. 215–218.
Благодарности
Работа выполнена при финансовой поддержке грантов Президента РФ НШ-3731.2010.4, НШ-1642.2012.4 и ФЦП ГК № 02.740.11.0700.

 © 2011 - 2026
© 2011 - 2026