



Введение
Ученые в разные времена делили животных на группы по отношению к температуре.
Холоднокровные – теплокровные, т. е. животные, которые «на ощупь» холодные или теплые. Первым, отметившим это обстоятельство, был, конечно, Аристотель (IV век до н. э.; но на русском языке результаты его биологических исследований были опубликованы только в первой половине ХХ в. – Аристотель, 1937). А уже в середине XIX в. замечательный немецкий анатом, физиолог и биолог Карл Бергман опубликовал работу, в которой отметил способность одних животных поддерживать постоянную высокую температуру тела, которая выше температур окружающей среды, а другие животные на это не способны (Bergmann, 1848). В этой работе он впервые применил термины «теплокровные» и «холоднокровные». Эти термины также использовал австрийский медик и анатом Франц Сетбер (Soetbeer, 1898). Затем, уже в начале XX в., финский медик и физиолог Роберт Тигерстедт, изучая теплообразование у разных животных, также применял термины «теплокровные» и «холоднокровные» (Tigerstedt, 1910). Такой же терминологией пользовался и замечательный, но забытый советский биолог Александр Владимирович Рюмин в своем подробном исследовании термобиологии позвоночных животных разных групп (Рюмин, 1939, 1940). В дальнейшем эти термины использовались в научной литературе очень широко (Glossary..., 2003 и др.).
Пойкилотермные – гомойотермные, т. е. животные с переменной или относительно постоянной температурой тела. Впервые эту пару терминов использовал Карл Бергман (Bergmann, 1848), а за ним и другие исследователи (Soetbeer, 1898; Tigerstedt, 1910; Bligh, Johnson, 1973 и др.). Эти термины и поныне широко используются (Glossary..., 2003 и др.).
Эктотермные – эндотермные, т. е. животные, которые получают тепло извне или производят его сами внутри организма за счет различных экзотермических реакций (Cowles, 1962). Термины применяются и сейчас (Glossary..., 2003 и др.).
Брадиметаболические – тахиметаболические, т. е. животные с низким или высоким уровнем метаболизма (Слоним, 1984).
Все эти термины по сути описывают разделение животных на две большие группы по отношению к температуре. Частично они могут быть применены соответственно к каждой из этих групп: с одной стороны – холоднокровные – пойкилотермные – эктотермные – брадиметаболические, с другой стороны – теплокровные – гомойотермные – эндотермные – тахиметаболические. Но каждая пара терминов описывает эти две группы лишь частично. Внутри каждой из этих двух больших групп (холоднокровные… и теплокровные…) характеристики указанных выше пар дополняют друг друга, создавая более полное представление о группах в целом.
Первые опубликованные сообщения о температуре тела холоднокровных (пойкилотермных…) животных относятся к концу XVII в., когда датский врач и натуралист Олигер Якобеус (Хольгер Якоби) писал, что у лягушек и ящериц температуры тела и окружающей воды практически не различаются (Jacobaeus, 1686). Затем, во второй четверти XVIII в., результаты своих исследований на эту же тему опубликовали французский естествоиспытатель, натуралист, энтомолог, физик и математик, впервые предложивший универсальный термометр и общую температурную шкалу, Рене Реомюр (Réaumur, 1736), изучавший насекомых, и шотландский врач Георг Мартин, изучавший возможности измерения температуры, в частности, на лягушках (Martine, 1740) и др.
В свете последних исследований становится очевидным, что в попытках систематизировать знания по этой проблеме исследователи до сих пор не учитывают важнейшие свойства животных, без осмысления которых описание и изучение этой сложнейшей проблемы становится неполным. Каждая из указанных выше пар понятий имеет в основе то или иное свойство: холодное или теплое тело, изменчивая или постоянная температура тела, поступление теплоты из внешних или внутренних источников, низкие или высокие уровни метаболизма, т. е. разные интенсивности термогенеза. Однако анализ эволюции термоэнергетики у позвоночных животных показал, что у них есть более глубинные различия – свойства «хамилотермии» и «псилотермии», описания которых приводятся ниже (впервые эти термины были применены нами – Черлин, 2021). А все пары терминов, указанные выше, являются в той или иной степени сопутствующими свойствами или их следствиями.
Аналитический обзор
Термоэнергетика
По современным представлениям, температура является одним из важнейших факторов, влияющих практически на все процессы в организме животных. При этом температура неразрывно связана с еще одним очень важным фактором – метаболизмом (Vernon, 1897; Tigerstedt, 1910 и др.). Процессы энергетического обмена являются ключевыми в понимании того, в каком направлении, как и почему животные с тем или иным анатомическим и физиологическим комплексом признаков формируют адаптивные взаимоотношения с внешней средой. При этом все обменные процессы самым непосредственным, жестким и прямым образом связаны с температурой. По сути, нам придется учитывать, рассматривать и изучать единый функциональный блок – «температура – метаболизм». Именно он и может быть назван термоэнергетикой. Может быть, более благозвучно было бы название «термометаболизм», но по крайней мере в англоязычной литературе оно уже преоккупировано – им обозначаются физиологические, метаболические процессы, связанные с термогенезом (например, «thermometabolism» в работе Legendre, Davesne, 2020 и др.). Поэтому наиболее приемлемый, на наш взгляд, вариант обозначения безусловно целостного физиологического комплекса, связывающего температурные и энергетические процессы и реакции, – это «термоэнергетика» (thermoenergetics).
Таким образом, можно обоснованно говорить о том, что реально существует определенный комплекс научных проблем, который в целом касается отношений биологии позвоночных животных с температурным фактором. Огромный массив научных данных, касающийся всех уровней организации живых систем (от биофизики и биохимии до физиологии и экологии), говорит об объективности этого комплекса (Van ‘T Hoff, 1884; Vernon, 1897; Arrhenius, 1889; Tigerstedt, 1910; Рюмин, 1939, 1940; Вильсон, 1952; Слоним, 1971; Шмидт-Нильсен, 1982; Рубин, 1987; Березов, Коровкин, 1998; Биохимия, 2004; Черлин, 2014 и др.).
Псилотермия и хамилотермия
Именно на уровне развития рептилий впервые четко проявилось важнейшее свойство, которое характерно только для высших позвоночных животных, – псилотермия.
Псилотермия (psilothermia) (греч. ψηλός – высокий, θέρμη – тепло). 1. Терморегуляционная реакция, направляющая животных на периодическое или постоянное повышение температуры тела (чаще всего более 28–30° и до примерно 45° в зависимости от видов). 2. Тип термобиологического статуса, при котором животные постоянно или часть времени в сутках поднимают и удерживают температуру тела (обычно выше 28–30°). Температура тела оказывается хотя бы на несколько градусов выше основного фона температур окружающей среды. Причем животные могут поднимать и удерживать температуру тела на высоком уровне разными способами: а) в основном за счет перераспределения в тепловом балансе тела идущих извне тепловых потоков благодаря поведенческим реакциям (эктотермия, экзотермия), или же б) в основном за счет изменения интенсивности термометаболизма (т. е. внутреннего сократительного или несократительного термогенеза) и теплоотдачи (эндотермия) (Черлин, 2014).
Важно здесь то, что и эктотермные рептилии, и эндотермные млекопитающие и птицы поднимают температуру своего тела выше 30° (то есть и те, и другие – псилотермы), только реализуют это разными способами. При этом сам феномен псилотермии явился естественным следствием и развитием принципа стабилизации высокой температуры тела в эволюции позвоночных животных (Рюмин, 1940; Черлин, 1990).
Хамилотермия (hamilothermia) (греч. χαμηλός – низкий, θέρμη – тепло) – тип термобиологического статуса, при котором у животных нет терморегуляционной псилотермической реакции, т. е. которые имеют температуру тела, мало отличающуюся от температур внешней среды, чаще всего ниже 28–30°, и у них не проявляются реакции, направленные на ее регулярное повышение (Черлин, 2024). Правда, при изучении хамилотермных животных есть по крайней мере одно важное методическое осложнение. Когда ученые изучают температуры тела, при которых рыбы, амфибии и рептилии активны, или их предпочитаемые температуры тела, то методики этих исследований для каждой из этих групп сильно отличаются друг от друга. Нет и единого понимания того, являются ли понятия «температура активности», «предпочитаемая температура» и другие термобиологические характеристики у этих групп аналогичными. Создается впечатление, что, скажем, предпочитаемые температуры тела для рыб, амфибий и рептилий, хоть могут быть названы одинаково, но на самом деле представляют собой совершенно разные характеристики, существенно отличающиеся по биологическому смыслу, по внутренним механизмам проявления и по методам определения. Проблема требует специального подробного изучения.
Хамилотермия рыб. Стремление повышать температуру тела и выбирать достаточно высокий диапазон температур тела у некоторых (далеко не у всех) групп рыб выражен, но достаточно слабо (Голованов, 2013 и др.).
Хамилотермия амфибий. Большинство амфибий по отношению к температуре мало отличаются от рыб.
Тем не менее по имеющимся данным можно сделать кое-какие выводы. Так, при изучении активности мраморной лягушки, Nannophrys marmorata в Шри-Ланке в разные сезоны (рис. 1) очевидно, что во влажный и прохладный сезон лягушки проводят бо́льшую часть времени в убежищах, где температура их тела остается в диапазоне 22–23° (рис. 2). В сухой и теплый сезон в середине дня, когда температура воздуха поднимается порой до 40°, лягушки, даже имея возможность, не стремились нагреваться и температура их тела и в это время не поднималась выше практически тех же 24–25° (Senanayake et al., 2019).
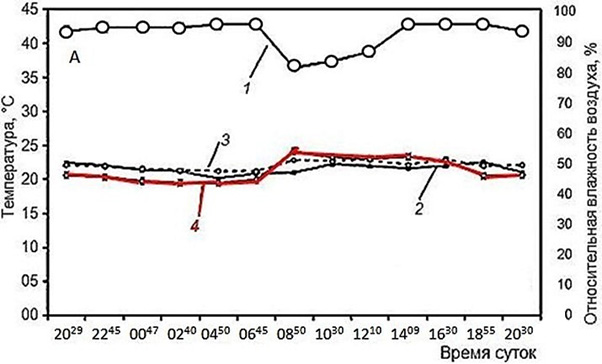
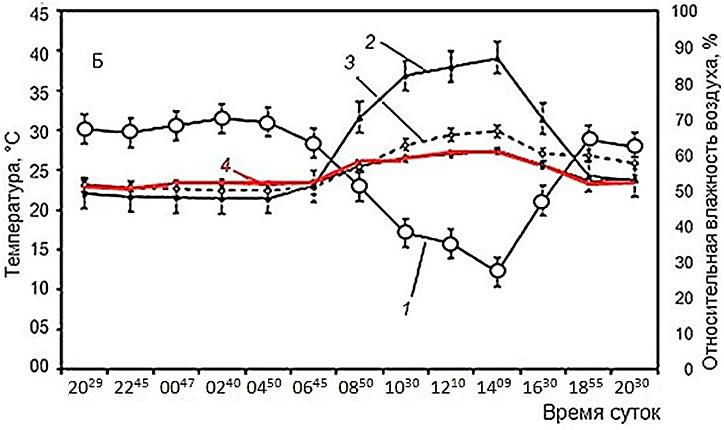
Рис. 1. Иллюстрация хамилотермического статуса. Активность мраморной лягушки в связи с условиями внешней среды в Шри-Ланка. А – влажный сезон, Б – сухой сезон (Senanayake et al., 2019). 1 – относительная влажность воздуха, 2 – температура воздуха, 3 – температура убежища, 4 – температура тела (красная линия)
Fig. 1. Illustration of the hamilothermal status. The activity of the marbled streamlined frog due to environmental conditions in Sri Lanka (Senanayake et al., 2019). A – wet season, Б – dry season. 1 – relative humidity, 2 – air temperature, 3 – shelter temperature, 4 – body temperature (red line)

Рис. 2. Мраморная лягушка в щели среди камней в Шри-Ланке (Senanayake et al., 2019)
Fig. 2. The marbled streamlined frog in a crevice among the stones in Sri Lanka (Senanayake et al., 2019)
Сильно повышать температуру тела для подавляющего большинства амфибий вообще и лягушек в частности смертельно опасно, поскольку при этом стремительно возрастают потери воды из организма. Эффективно сопротивляться этому могут только некоторые пустынные жабы, у которых кожа приобретает достаточно большую плотность и может становиться сухой, что препятствует потере воды их организмом. Периодически температура их тела может подниматься даже до 39° (Pearson, Brandford, 1976).
Псилотермия рептилий характерна тем, что данная терморегуляционная реакция заставляет этих животных постоянно или часть времени в сутках поднимать и удерживать температуру тела в пределах 28–45° (чаще всего в диапазонах ~28–35, 35–39, 36–42, 38–45° в зависимости от вида животного). Температура тела оказывается хотя бы на несколько градусов выше основного фона температур окружающей среды. Поднимать и удерживать температуру тела на высоком уровне псилотермы могут в основном эктотермным путем (в отдельных, редких случаях используются элементы эндогенного пути).
Проиллюстрировать псилотермию рептилий можно на примере среднеазиатской эфы, Echis multisquamatus, ушастой круглоголовки, Phrynocephalus mystaceus и обыкновенной гадюки, Vipera berus.
На рис. 3, 4 и 5 показано, что в любые сезоны года при активности рептилий, в ситуациях, когда условия внешней среды позволяют, эти животные благодаря внешним источникам тепла и поведенческим регуляторным реакциям поднимают и удерживают температуру своего тела в более или менее определенном диапазоне.
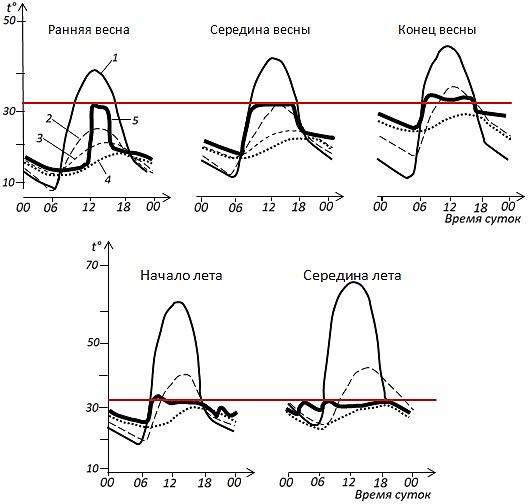
Рис. 3. Иллюстрация псилотермического статуса. Активность среднеазиатской эфы в связи с условиями внешней среды (по: Черлин, Целлариус, 1981). 1 – температура поверхности почвы, 2 – температура воздуха на высоте 2 см, 3 – температура почвы на глубине 20 см, 4 – температура почвы на глубине 30 см, 5 – температура тела. Красная линия показывает уровень температуры примерно 32°
Fig. 3. Illustration of psilothermal status. The activity of the Saw-scaled viper due to environmental conditions (by Cherlin, Tsellarius, 1981). 1 – soil surface temperature, 2 – air temperature at a height of 2 cm, 3 – soil temperature at a depth of 20 cm, 4 – soil temperature at a depth of 30 cm, 5 – body temperature. The red line shows a temperature level of about 32°
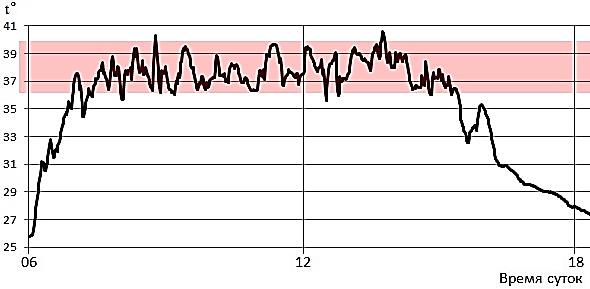
Рис. 4. Иллюстрация псилотермического статуса. Суточный ход температуры тела ушастой круглоголовки на бархане Сарыкум (Дагестан, РФ) 31 мая 2024 г., полученный с помощью внедренного парентерально миниатюрного логгера температуры, регистрировавшего и запоминавшего температуру тела раз в минуту. t° – температура. Розовой полосой отмечен преимущественный диапазон температуры тела (~36–40°), который ящерица направленно удерживает у себя в дневное время, ведя очень подвижный образ жизни в мозаичных по температуре условиях внешней среды (при температуре поверхности песка на разных участках ~ от 30 до 60°)
Fig. 4. Illustration of psilothermal status. The daily course of body temperature in the toad-headed agama, Phrynocephalus mystaceus, on the Sarykum dune (Dagestan, Russia) on May 31, 2024, obtained using a parenterally embedded miniature temperature logger that recorded and stored body temperature once a minute. t° is the temperature. The pink stripe marks the predominant range of body temperature (~36–40 °), which the lizard directionally retains during the daytime, leading a very mobile lifestyle in mosaic-like ambient conditions (with sand surface temperatures in different areas ranging from 30 to 60°)
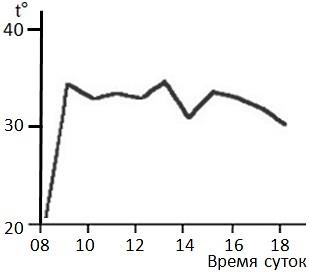
Рис. 5. Суточный ход температуры тела обыкновенной гадюки в мае в Карелии, полученный с помощью телеметрии (по: Коросов, 2008), иллюстрирует ее терморегуляционное поведение и псилотермический статус
Fig. 5. The daily course of body temperature of the common adder in May in Karelia obtained by using of telemetry (according to Korosov, 2008) illustrates its thermoregulatory behavior and psilothermic status
Экологический смысл преимущества подъема температуры тела, т. е. наличия псилотермической реакции, связан, в частности, с тем, что при высокой температуре подавляющее большинство химических реакций, а значит, и физиологических процессов, и процессов в нервной системе происходят быстрее. Особенно это важно для огромных по размерам и массе динозавров, гигантозавров (бронтозавров, диплодоков, брахиозавров и др.) длиной до 30–35 м и весом до 50–80 т, огромных спинозавров, тираннозавров и т. п. При низких температурах тела (даже около 25°) их движения, реакции на внешние раздражители (даже на опасности), требующие порой быстрых, стремительных, оперативных ответов, были бы медленными, плавными. Такие животные не способны были бы ни добывать большое количество необходимых для них кормов, ни их переваривать и усваивать, ни оперативно избегать хищников и других внешних опасностей. В реальной природной обстановке они были бы не способны выжить. Так что иметь высокую температуру тела, и к тому же желательно независимо от климатических условий внешней среды, для таких животных – не абстрактное, отвлеченное, желательное условие, а насущная необходимость. На это же обстоятельство указывал и Валерий Михайлович Гаврилов (Гаврилов, 2012).
Но первым на это важное обстоятельство обратил внимание Александр Владимирович Рюмин (Рюмин, 1940), затем Иван Дмитриевич Стрельников (Стрельников, 1948), а уже потом мы (Черлин, 1990 и др.).
Изучая термобиологию пресмыкающихся и представив вариант организации у них механизма регуляции температуры тела в нервной системе, мы, помимо прочих, описали тип терморегуляционной реакции рептилий, названный нами «реакцией максимизации температуры тела». «Проявление этого механизма выражается в стремлении к повышению, "максимизации" температуры вплоть до верхней границы. В полевых и лабораторных исследованиях этот регулятор проявляется в том, что рептилии, имея низкую температуру тела, в подавляющем большинстве случаев пытаются различными, преимущественно поведенческими приемами ее повысить до допустимой верхней границы. Такая ситуация регулярно наблюдается у всех видов рептилий. Она четко проиллюстрирована, в частности, описанием стратегии терморегуляции у обыкновенных гадюк в Карелии (Коросов, 2008), выражающейся в стремлении к повышению температуры тела до определенного, достаточно высокого уровня. Можно предположить, что, исходя из структуры, описанной моделью А. В. Коросова, штатное значение физиологической константы фиксирует высокий, практически недостижимый для животного уровень температуры тела, к которому она должна стремиться. При увеличении величины отклонения температуры тела от штатного значения константы, поведенческие реакции обратной связи повышают температуру тела (в идеале – до штатного значения)» (Черлин, 2014, с. 148–149).
Таким образом, еще на два года раньше нас Андрей Викторович Коросов, предложив модель терморегуляции обыкновенной гадюки (Коросов, 2008), показал, что рептилии (по крайней мере – этот вид змей) стремятся к повышению температуры тела до уровня примерно 28–30° или даже выше. «Максимальная добровольная температура (Тм), физиологическая константа терморегуляции, играет роль целевого параметра, к которому система стремится, но достичь почти никогда не может… Обычно же достигается менее высокая температура – 28–30 °С. Но это и есть нужная "оптимальная" (с физиологической точки зрения) температура для гадюки (То), при которой должным образом идут процес,сы обмена, роста, пищеварения и пр… Здесь Тм является псевдоцелью, стремление к которой обеспечивает истинную цель, физиологически оптимальную температуру тела» (Коросов, 2008, с. 65). Здесь также идет речь о некоем механизме, заставляющем змей повышать температуру тела (стремление к псевдоцели – высокой, недостижимой температуре тела).
Таким образом, сейчас уже существуют осторожные подходы к тому, чтобы понять, каким образом может быть организовано и поддержано состояние псилотермии у позвоночных животных. Но это лишь предположения. Проблема требует дальнейших серьезных исследований, поскольку она имеет очень важное значение для понимания сути и функциональной направленности развития отношения животных с тепловым фактором, теплокровности птиц и млекопитающих, механизмов организации и, при необходимости, путей воздействия на терморегуляционную сферу у человека и т. п. И здесь важно понять три основные вещи:
1) являются ли описанные выше аспекты именно теми, которые определяют механизмы проявления и закрепления стратегического направления развития термобиологического статуса – псилотермии;
2) базируется ли поддержание псилотермии для разных уровней организации на одних и тех же материальных основаниях, на одних и тех же биохимических и организационных механизмах, а если на разных – то на каких;
3) почему именно уровень температуры тела 28–30° является настолько важным, что становится неким «водоразделом» в отношениях позвоночных животных с температурой, разделяющим хамилотермов и псилотермов, анамний и амниот, каково физиологическое значение этого температурного уровня.
К сожалению, удовлетворительных, исчерпывающих, научно обоснованных ответов на эти вопросы мы в литературе пока не находим. Их еще предстоит дать будущим поколениям ученых. А пока мы можем только констатировать эмпирически выявленные факты, что уже не мало. В частности, становится понятно, что серьезное внимание в исследованиях по биологии позвоночных животных следует в значительной степени уделять не только механизмам термогенеза и способам поступления тепла в организм животных, но и механизмам, вызывающим физиологическую потребность в повышении и поддержании высокой температуры тела. Способы поступления тепла в организм – лишь способы реализации этой стратегически важной физиологической потребности.
Одной из биологических основ псилотермии является, видимо, определенный уровень клеточного митохондриального дыхания, который примерно в два раза повысился по сравнению с исходным, амфибийным, хамилотермическим состоянием (Черлин, 2017). Он улучшил энергетическую обеспеченность активности, заложил основу для будущего комплекса поведенческих терморегуляторных реакций, а также для биохимических механизмов тахиметаболизма.
Другой биологической основой псилотермии является механизм регуляции температуры тела, который стал обеспечивать ее направленность на повышение по крайней мере до 28–30° или выше. Что так повлияло на стабилизацию установочного регулирующего параметра на этом уровне – вопрос пока открытый. Но он тоже требует решения.
Псилотермия теплокровных животных очевидна: температура их тела поддерживается благодаря прежде всего несократительному и сократительному (в частности – дрожательному) термогенезу (эндотермия) на почти постоянном уровне, который практически всегда и у всех теплокровных животных выше 30°.
Псилотермы могут поднимать и удерживать температуру тела на высоком уровне разными способами: 1) в основном за счет поступающего извне тепла и его перераспределения в теле благодаря поведенческим и физиологическим реакциям (относительная аналогия в общепринятых терминах – холоднокровность, эктотермия, пойкилотермия, брадиметаболизм), или же 2) в основном за счет внутреннего сократительного или несократительного термогенеза (относительная аналогия в общепринятых терминах – теплокровность, эндотермия, гомойотермия, тахиметаболизм). Псилотермическими являются все позвоночные животные, которые достигли уровня развития рептилий: среди диапсид – первые же базовые архозавроморфы, развитые архозавры, включая птицетазовых и ящеротазовых динозавров, а также птерозавры, морские ящеры и настоящие птицы; среди синапсид – базовые синапсиды (пеликозавры, териодонты), развитые млекопитающие (Черлин, 2024).
Здесь, наверно, стоит проанализировать определения двух других, уже употребляемых и привычных, терминов, с которыми новые термины не следует путать.
Холоднокровное животное – «тепловое состояние животного, при котором температура ядра тела остается близкой к температуре окружающей среды при воздействии низкой температуры окружающей среды…» (Glossary…, 2003, p. 79).
Теплокровное животное – «тепловое состояние животного, которое поддерживает температуру ядра своего тела значительно выше, чем температура окружающей среды при воздействии низкой температуры окружающей среды…» (Glossary…, 2003, p. 103).
Но в определениях холоднокровности имеется одно спорное обстоятельство. Температура тела холоднокровных животных действительно очень сильно зависит от температур окружающей среды. Однако это проявляется лишь в определенном варианте экспериментальных условий, когда у животных нет возможности температуру тела регулировать, т. е. менять. И тут проблема упирается в понятие «терморегуляция».
Традиционно в Советском Союзе, и до сих пор в Российской Федерации многие ученые (особенно физиологи) делятся на две группы по отношению к понятию «терморегуляция». В результате своих представлений о терморегуляции одни из них отказывают рептилиям в наличии терморегуляции, другие – нет.
Отрицающие терморегуляцию у рептилий ученые вкладывают в это понятие следующий смысл: «Терморегуляция – совокупность физиологических процессов, обеспечивающих постоянство температуры тела у теплокровных животных (птиц и млекопитающих) и человека. Осуществляется путем изменения интенсивности теплопродукции (при окислительных процессах в организме) и путем изменения теплоотдачи через кожу (испарение пота и др.)» (Большой…, 2000). Другими словами, терморегуляция в таком понимании заведомо связывается исключительно с биохимическими и физиологическими эффекторными системами и подходит только эндотермным, теплокровным животным. Но это произвольная трактовка, не отражающая всей сложности самого явления терморегуляции в природе у разных групп животных.
Другие ученые вкладывают в этот термин более широкий смысл. Например: «Терморегуляция… физиологическая функция, обеспечивающая поддержание оптимальной для данного вида температуры глубоких областей тела в условиях меняющейся температуры окружающей среды. Способность к терморегуляции в значительной мере определяет границы расселения и выживания животных в различных климатических условиях и является одним из важных механизмов их гомеостаза» (Биологический…, 1986, с. 627). В таком варианте терморегуляция вполне может быть приписана и рептилиям.
Четкое определение терморегуляции было дано Комиссией по термальной физиологии при Международном обществе физиологических наук (Commission for Thermal Physiology of the International Union of Physiological Sciences): «Регулирование температуры[1]: поддержание температуры или температур тела в ограниченном диапазоне в условиях переменных внутренних и/или внешних тепловых нагрузок. Регулирование температуры тела осуществляется в некоторой степени вегетативными или поведенческими средствами» (Glossary…, 2003, p. 97). В круг действия этого определения в полной мере входят и реакции рептилий, направленные на контроль за температурой своего тела в основном поведенческими способами.
Замечательный русский / советский ученый Иван Дмитриевич Стрельников уделял большое внимание изучению терморегуляции насекомых и рептилий. Еще в начале тридцатых годов прошлого века он не признавал у них возможностей регуляции температуры тела: «Пойкилотермные животные не обладают способами регулирования температуры тела и поддержания ее в узких пределах нескольких градусов, как у гомойотермных» (Стрельников, 1934, с. 361). И хотя это была довлеющая на то время концепция среди биологов, тем не менее его собственные исследования часто ставили это положение под сомнение: «Среди разнообразных и непрерывно меняющихся экологических условий Stenodes caspius и другие обитатели пустыни лавируют таким образом, что сохраняют некоторое постоянство наиболее для благоприятных условий. Прохладно в воздухе – жуки вылезают из песка, питаются, спариваются; жарко или холодно – зарываются в песок, где находят для себя ту же температуру, что была для них в воздухе» (Стрельников, 1934, с. 354). Причем, обратите внимание: две последних цитаты взяты из одной статьи!
Но Иван Дмитриевич Стрельников был действительно замечательный, думающий ученый, и его полевые исследования и эксперименты в конечном итоге заставили его изменить свою точку зрения. «…Рептилии в период активной жизни днем имеют температуру тела в среднем около 35–36º (30–40º), т. е. такую температуру тела, как млекопитающие. Рептилии активно поддерживают температуру своего тела движением или, чаще и больше всего, греясь лучами солнца; зарываясь в песок, прячась в тень, они избегают перегревания» (Стрельников, 1944, с. 256).
Кроме чисто поведенческих реакций И. Д. Стрельников показал и другие приемы терморегуляции у рептилий и даже у амфибий. «Одним из главных приспособлений пустынных животных является их окраска. Преобладающей окраской жителей пустыни являются светлые окраски, отражающие значительную часть падающих лучей. Во время моего пребывания в Репетеке, мне удалось отметить, что ушастая круглоголовка Phr. mystaceus рано утром при более низких температурах имеет темную окраску. По мере поднятия температуры, в особенности около полудня, Phr. mystaceus становятся очень светлыми, и имеют соломенно-желтоватые светлые тона в окраске. Темный пигмент, рассеянный по телу у ящериц в холодном воздухе, концентрируется в пятнах, рассеянных по светло-желтоватому фону» (Стрельников, 1934, с. 357–358). Эти закономерности он проиллюстрировал своими опытами и на ряде видов земноводных – жабах и лягушках (Стрельников, 1944). Таким образом, безусловное наличие терморегуляции у амфибий и в особенности у рептилий было практически доказано.
Однако значительная часть физиологов отрицала возможность терморегуляции у пойкилотермных (эктотермных) животных (Бахметьев, 1899; Стрельников, 1934; Коштоянц, 1950 и др.). Основной их довод сводился к следующему: существует вполне научно доказанное утверждение, что температура тела пойкилотермных животных (в частности амфибий и рептилий) напрямую зависит от температуры окружающей среды. Если такое животное поместить в экспериментальные условия с температурой воздуха, например, 25°, то температура его тела также будет 25° (± примерно 1°); при температуре воздуха 30° температура тела животного также будет примерно 30° и т. п. Исходя из этого авторы делают заключение о том, что у этих животных терморегуляции нет.
Дефект этого подхода очевиден: в таком эксперименте во внешней среде температура однородная и нет никакого ее разнообразия. Значит, животное помещается в такие условия, в которых терморегуляция, т. е. возможность изменить температуру тела, заведомо может быть только физиологической (за счет изменения внутренней теплопродукции). Поведенческая терморегуляция в таких условиях принципиально невозможна. Но причиной этого является не неспособность животного к терморегуляции, а отсутствие возможности такой регуляции из-за однородности среды, т. е. некорректность поставленного опыта.
Наши исследования термобиологии рептилий однозначно подтверждают наличие терморегуляции, например, у ящериц. Так, в пустынях Средней Азии живут сугубо ночные ящерицы – сцинковые гекконы, Teratoscincus scincus. Они появляются на поверхности почвы только после захода солнца (Богданов, 1965) и встречаются там при температурах внешней среды и тела примерно от 16 до 33° (рис. 6) (Черлин, 2013).
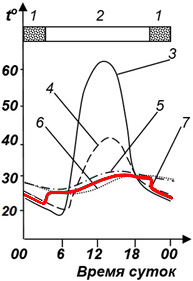
Рис. 6. Связь температуры тела сугубо ночного сцинкового геккона с температурными условиями среды в летний период (по: Черлин, 2013). t° – температура; формы активности: 1 – активность на поверхности в темное время суток, 2 – сон / отдых (дневной); температуры: 3 – поверхности почвы; 4 – воздуха на высоте 3 см, 5 – почвы на глубине 20 см, 6 – почвы на глубине 30 см, 7 – (красная линия) тела гекконов
Fig. 6. Relationship of body temperature of a purely nocturnal Turkestan plate-tailed gecko with ambient temperature conditions in summer (according to Cherlin. 2013). t° – temperature; forms of activity: 1 – activity on the soil surface at night, 2 – sleep / rest (daytime); temperatures: 3 – soil surface; 4 – air at a height of 3 cm, 5 – soil at a depth of 20 cm, 6 – soil at a depth of 30 cm, 7 – (redline) body temperature of geckos
Все дневное, светлое время суток сцинковые гекконы проводят в норах. Если посмотреть на динамику различных климатических параметров на поверхности и в глубине почвы, то становятся очевидными важные закономерности. Суточная динамика температуры тела сцинковых гекконов неслучайна. Несмотря на то, что температура тела гекконов сильно зависит от температурных условий внешней среды (как в описанных выше экспериментах физиологов), она кроме того подчиняется и своим внутренним закономерностям. Даже в различных климатических условиях, в разные сезоны года динамика температуры тела гекконов имеет вполне определенный характер и ее суточные графики изо дня в день повторяются. При этом суточный ход температуры тела сцинковых гекконов не повторяет суточную динамику ни одного из температурных параметров ни одной точки внешней среды. Значит, динамика температуры тела ящериц направленно задается определенными внутренними требованиями и фактически формируется их определенными поведенческими реакциями в разнообразных условиях внешней среды. Другими словами, у них проявляется хорошо развитая терморегуляция. Такая же ситуация ясно видна и на примерах со среднеазиатской эфой и ушастой круглоголовкой (см. рис. 3 и 4).
Таким образом, если терморегуляцию понимать как свойство, присущее только теплокровным животным, то современные рептилии, естественно, оказываются типично холоднокровными животными. Если же термин «терморегуляция» понимать корректно, то термобиологический статус современных рептилий оказывается в определенной степени «промежуточным» между холоднокровными и теплокровными. И при этом становится очевидно, что терморегуляционные реакции рептилий настроены так, что они являются эффекторными механизмами, реализующими феномен псилотермии.
В результате, если понятия «холоднокровность» и «хамилотермия» по отношению к рыбам и амфибиям оказываются практически идентичными, то понятия «теплокровность» и «псилотермия» различаются. Теплокровные животные – те, которые поддерживают высокую температуру тела преимущественно за счет высокого уровня термометаболизма, причем несократительного, а псилотермы проявляют направленность своих поведенческих и физиологических реакций (в т. ч. и термометаболизма) на периодическое или постоянное повышение и стабилизацию высокой температуры тела выше 28–30° любыми доступными путями (экзогенным или эндогенным поступлением тепла, физиологической и/или поведенческой терморегуляцией). Рептилии вместе с типичными теплокровными животными являются псилотермами. Реакция псилотермии объединяет их биологию на глубоком, базовом уровне.
Следовательно, рептилий как класс позвоночных животных во всем своем огромном разнообразии вымерших и современных форм можно рассматривать как животных со сложным и неоднозначным термобиологическим статусом. Внутри общности, которую мы называем рептилиями, имеются всевозможные варианты реализации псилотермии, которые приводили к появлению групп с физиологией от типичных эктотермов до классических эндотермов с различной степенью термометаболизма, правда, видимо, с недостаточно еще развитой системой регуляции теплопродукции (Черлин, 2024). У современных же рептилий имеется определенная «двоякость» термобиологического статуса. С одной стороны, они получают тепло преимущественно извне организма, т. е. должны считаться типично эктотермными животными. С другой стороны, у них четко проявляется свойство псилотермии, которое определенно направляет их к периодическому повышению и поддержанию достаточно высоких температур тела (в зависимости от видов от 28–30 до 44–45°). Другими словами, современные рептилии – эктотермные псилотермы. Именно в связи с этим А. В. Рюмин удачно назвал их «потенциально теплокровными» животными (Рюмин, 1939).
Рассматривая распределение псилотермии и хамилотермии среди позвоночных животных, можно заключить, что все анамнии – хамилотермы, а все амниоты – псилотермы. Смысл этой связки еще предстоит изучить и понять.
Таким образом, мы описали наличие биологического феномена псилотермии. Это свойство устанавливает определенную систему взаимоотношений животных с внешней средой, их экологию, принципы организации и функционирования систем внутренних органов, морфофизиологию потомков и т. п.
Морфофизиологическая организация позвоночных как группы животных, систем их внутренних органов несет в себе их важные базовые характеристики, которые отражаются на разных сторонах биологии всех групп, начиная по крайней мере с рептилий. Но самих этих характеристик мы до сих пор не знаем. Причем, как мы уже отметили, биологические особенности псилотермии входят в биологическую базу амниот в целом, млекопитающих, и человека в частности. А мы до сих пор не знаем, что представляет собой псилотермия, какой она охватывает круг биологических характеристик животных, в чем состоит ее биологический механизм, каких еще внутренних систем она касается, что она значит в биологии разных групп, и человека в частности.
[1] Термины «терморегуляция» и «регулирование температуры» («Thermoregulation» и «Temperature regulation») – синонимы, что специально подчеркнуто в словаре (Glossary…, 2003, p. 101).
Заключение
Анализ конкретных данных по температурам тела, принципам организации и развития термобиологических статусов в разных группах позвоночных животных (вымерших и современных) дал возможность выявить и описать базовое свойство позвоночных животных, появившееся впервые у рептилий. Это псилотермия – терморегуляционная реакция, направляющая животных на то, чтобы постоянно или периодически поднимать и удерживать температуру тела (чаще всего выше 28–30°) посредством тепла, поступающего или извне организма (эктотермная псилотермия некоторых вымерших и всех современных рептилий), или изнутри (эндотермная псилотермия большинства вымерших архозавроморфов, архозавров, ихтиоптеригий и некоторых других диапсид, продвинутых синапсид, птиц и млекопитающих). Исходным состоянием является хамилотермия – тип термобиологического статуса, при котором у животных нет реакции псилотермии, они имеют температуру тела, мало отличающуюся от температур внешней среды, чаще всего ниже 28–30° (рыбы и амфибии). Псилотермия способствовала повышению температуры тела, усилению основного обмена, улучшению качества активности и экологической конкурентоспособности позвоночных животных.
Описанный нами механизм псилотермии является важнейшим, базовым, фундаментальным свойством всех групп позвоночных животных начиная с рептилий. Именно его развитие привело к появлению и совершенствованию комплексов свойств эктотермных (холоднокровных) и эндотермных (теплокровных) животных.
Действие этого механизма проявляется у млекопитающих, а следовательно, и у человека. Поскольку этот механизм базовый, значит, должен иметь огромное значение в функционировании физиологических систем организма. Для человека это должно иметь большое значение, поскольку этот механизм принципиально важен как с точки зрения физиологической, так и с позиций терапевтической медицины. Но о нем, о его устройстве и функционировании в нервной системе, значении в организации и работе систем терморегуляции и других физиологических систем и функций организма в норме и патологии, в процессе адаптаций пока вообще ничего не известно. На наш взгляд, изучение этого механизма должно сыграть серьезную роль в развитии наших представлений о деятельности организма человека, о возможности на нее воздействовать, менять уровень адаптабельности и исправлять патологические состояния.
Библиография
Бахметьев П. И. Собственная температура пчел и вообще насекомых СПб.: Тип. В. Демакова, 1899. 11 с.
Березов Т. Т., Коровкин Б. Ф. Биологическая химия М.: Медицина, 1998. 704 с.
Биологический энциклопедический словарь М.: Советская энциклопедия, 1986. 831 с.
Биохимия / Ред. Е. С. Северин. М.: Изд. дом "ГЭОТАР-МЕД", 2004. 784 с.
Богданов О. П. Экология пресмыкающихся Средней Азии Ташкент: Наука, 1965. 259 с.
Большой энциклопедический словарь 2000 URL: https://bcoreanda.com/ShowTreck.aspx?ID=459 (дата обращения: 12.03.2025).
Вильсон П. В. Дыхательные ферменты М.: Иностранная литература, 1952. 416 с.
Гаврилов В. М. Экологические, функциональные и термодинамические предпосылки и следствия возникновения гомойотермии на примере исследования энергетики птиц // Журнал общей биологии. 2012. Т. 73, № 2. С. 88–113.
Голованов В. К. Температурные критерии жизнедеятельности пресноводных рыб М.: Полиграф-Плюс, 2013. 300 с.
Коросов А. В. Простая модель баскинга обыкновенной гадюки (Vipera berus L.) // Современная герпетология. 2008. Т. 8, вып. 2. С. 118–136.
Коштоянц Х. С. Основы сравнительной физиологии Т. 1. М.; Л.: Изд-во АН СССР, 1950. 524 с.
Рубин А. Б. Биофизика клеточных процессов Кн. 2. М.: Высшая школа, 1987. 303 с.
Рюмин А. В. Значение температуры в онтогенезе и филогенезе животных // Успехи современной биологии. 1940. Т. 12, № 3. С. 504–515.
Рюмин А. В. Температурная чувствительность позвоночных животных и биологический путь происхождения теплокровных форм // Сборник студенческих научных работ МГУ. 1939. Вып. 6. С. 55–84.
Слоним А. Д. Физиология терморегуляции Л.: Наука. Ленингр. отд-ние, 1984. 378 с.
Слоним А. Д. Экологическая физиология животных М.: Высшая школа, 1971. 448 с.
Стрельников И. Д. Значение солнечной радиации в экологии высокогорных рептилий // Зоологический журнал. 1944. Т. 23, № 5. С. 250–256.
Стрельников И. Д. Значение солнечной радиации и взаимодействия физико-географических факторов в экологии животных различных ландшафтов (К вопросу о значении физической географии в экологии животных) // Проблемы физической географии. 1948. № 13. С. 145–155.
Стрельников И. Д. Свет как фактор в экологии животных. Статья первая. Действие солнечной радиации на температуру тела некоторых пойкилотермных животных (к экологии животных пустыни Кара-Кумы) // Известия Научного института имени П. Ф. Лесгафта. 1934. Т. 17–18. С. 313–372.
Черлин В. А. Значение изменений интенсивности сопряженного и несопряженного дыхания митохондрий в эволюции позвоночных животных // Успехи современной биологии. 2017. Т. 137, № 5. С. 479–497.
Черлин В. А. Рептилии: температура и экология. Saarbrücken: Lambert Academic Publishing, 2014. 442 с.
Черлин В. А. Связь между эктотермией и эндотермией в эволюции позвоночных животных // Журнал общей биологии. 2024. Т. 85, № 3. С. 244–266.
Черлин В. А. Стабилизация высокой температуры тела в эволюции позвоночных животных // Успехи современной биологии. 1990. Т. 109, № 3. С. 440–452.
Черлин В. А. Сравнение термобиологии сцинкового (Teratoscincus scincus scincus) и гребнепалого (Crossobamon eversmanni) гекконов // Вестник Тамбовского университета. Серия: Естественные и технические науки. 2013. Т. 18, вып. 6. С. 3110–3112.
Черлин В. А. Эволюция термобиологических статусов у позвоночных животных. Статья 2. Развитие отношений с температурой у позвоночных животных // Журнал общей биологии. 2021. Т. 82, № 6. С. 459–477.
Черлин В. А., Целлариус А. Ю. Зависимость поведения песчаной эфы, Echis multisquamatus Cherlin 1981 от температурных условий в Южной Туркмении // Фауна и экология амфибий и рептилий палеарктической Азии. Л.: Наука, 1981. С. 96–108 (Труды Зоологического института АН СССР, т. 101).
Шмидт-Нильсен К. Физиология животных Т. 1. М.: Мир, 1982. 414 с.
Arrhenius S. A. Über die Dissociationswärme und den Einfluß der Temperatur auf den Dissociationsgrad der Elektrolyte // Z. Phys. Chem. 1889. Vol. 4, issue 1. S. 96–116.
Bergmann C. Uber die Verhältnisse der Wärmeökonomie der Tiere zu ihrer Größe. Göttingen, 1848. 117 S.
Bligh J., Johnson K. G. Glossary of terms for thermal physiology // J. Appl. Physiol. 1973. Vol. 35, No 6. Р. 941–961.
Cowles R. B. Semantics in biothermal studies // Science. 1962. No 135. P. 670.
Glossary of terms for thermal physiology. Third Edition revised by The Commission for Thermal Physiology of the International Union of Physiological Sciences (IUPS Thermal Commission) // Journal of Thermal Biology. 2003. No 28. Р. 75–106.
Jacobaeus O. De ranis et lacertis observations. Hafniae: Johannis M. Lieben., 1686. 174 p.
Legendre L. J., Davesne D. The evolution of mechanisms involved in vertebrate endothermy // Phil. Trans. R. Soc. B. 2020. Vol. 375 (1793). P. e20190136.
Martine G. Essays medical and philosophical. Milla: London, 1740. 392 p.
Pearson O., Brandford D. F. Thermoregulation of lizards and toad at high altitudes in Peru // Copeia. 1976. № 1. P. 155–170.
Réaumur R. Mémoirs pour servir à l’Histoire des Insectes. Tome 2. Paris: d’Imprimerie Royal., 1736. 514 p.
Senanayake U. I., Siriwardana S., Weerakoon D. K., Wijesinghe M. R. Combating Extreme Tropical Seasonality: Use of Rock Crevices by the Critically Endangered Frog Nannophrys marmorata in Sri Lanka // Herpetological Conservation and Biology. 2019. Vol. 14, No 1. P. 261–268.
Soetbeer F. Uber die Körperwärme der poikilothermen Wirbeltiere // Arch. f. exp. Pathol. Bd. 1898. Vol. 40. Р. 53–80.
Tigerstedt R. Die Production von Wärme und der Wärmehaushalt (S. 1–104) / Handbuch der vergleichenden Physiologie herausg. von Winterstein., 1910. 1060 S.
Van ‘T Hoff J. H. Études de dynamique chimique. Amsterdam: F. Muller & Co., 1884. 236 p.
Vernon H. M. The relation of the respiratory exchange of cold-blooded animals to temperature // J. Phisiol. 1897. Vol. 21 P. 443–496.
Благодарности
Благодарю кандидата биологических наук Валентину Николаевну Куранову, доцента Томского государственного университета, доктора биологических наук Бориса Дмитриевича Куранова, ведущего научного сотрудника Томского государственного университета, доктора биологических наук Георгия Аркадьевича Ладу, профессора Тамбовского государственного университета им. Г. Р. Державина, а также Игоря Юрьевича Барсукова и Игоря Леонидовича Окштейна за ценные замечания и советы.

 © 2011 - 2026
© 2011 - 2026