



Введение
Изучение и подробный экологический анализ приспособительных особенностей волосяного покрова мелких млекопитающих представляет большой научный интерес, и прежде всего с позиций эволюционной морфологии, экологической физиологии и таксономии этой весьма распространенной и многочисленной в природе группы животных. Массовое обитание их в лесной подстилке и ходах неглубоких нор в условиях постоянных термических дефицитов, как и свойственное им общее несовершенство терморегуляции, способствовали выработке в процессе их эволюции целого комплекса экологических и морфофизиологических приспособлений, направленных на сохранение оптимального энергетического баланса организма со средой при низких и крайне неустойчивых окружающих температурах. Отсюда важность углубленного исследования таких адаптивных механизмов физической терморегуляции мелких зверьков, как строение и теплозащитные свойства шерсти, микроструктура волос, географическая, сезонно-возрастная и популяционная изменчивость этих показателей, выявление особенностей строения шерстного покрова, связанных с обитанием в специфических температурных условиях приземного слоя воздуха, и т. п.
Надо учитывать также и общую относительно слабую изученность данного вопроса, тем более что имеющиеся в отечественной и зарубежной литературе сведения о строении и функционировании волосяного покрова мелких млекопитающих не только далеко не полны, но и во многом противоречивы. Так, если Н. С. Гудкова-Аксенова (1951), одна из первых осмелившаяся взяться за изучение волосяного покрова землероек-бурозубок, выделяет у этих зверьков только две категории волос, то В. Е. Соколов и О. Ф. Чернова (Соколов, 1973; Соколов, Чернова, 1998; Чернова, Целикова, 2004) – четыре, а П. Фогель и Б. Кепхен (Vogel, Köpchen, 1978) и P. Хуттерер и Т. Хёртер (Hutterer, Hurter, 1981) – три, то, по мнению С. Боровского (Borowski, 1973) и У. Скарена (Scaren, 1973), у землероек-бурозубок волосы одного и того же типа способны принимать разный вид в зависимости от конфигурации расширенной апикальной части стержня (концевой гранны). Отрицая дифференцировку волос у землероек-бурозубок на направляюшие, остевые и пуховые, С. Боровский обращает внимание на различную степень их сегментации и в связи с этим выделяет «следующие 3 типа волос: 1) летние волосы (S) с четырьмя сегментами, иногда с коротким кончиком (пятый короткий сегмент); 2) весенние волосы (Sр) с пятью сегментами, но иногда с шестым сегментом (около 5 % волосков); 3) зимние волосы (W) – шестисегментные, иногда с дополнительным седьмым концевым сегментом» (Borowski, 1973, с. 249). В действительности же, как убедительно показали и наши исследования, выполненные на обыкновенной бурозубке (Ивантер, 2021), летняя, весенняя и зимняя шерсть землероек состоит из направляющих, остевых и пуховых волос, а волосы с «дополнительным коротким концевым сегментом» есть не что иное, как пуховые.
Что же касается характерной для мелких насекомоядных млекопитающих сегментации волос, то в отличие от С. Боровского мы склонны считать это не примитивным атавистическим признаком, а важной, выработанной в процессе эволюции адаптацией к специфическим условиям существования. Здесь вполне уместно вспомнить замечание, содержащееся в давней статье И. С. Башкирова и И. В. Жаркова (1934), которые, говоря о пережимах на стержнях остевых и пуховых волос, свойственных представителям семейств Talpidae и Soricidae, не без оснований предполагают, что эти структурные особенности имеют большое приспособительное значение. Благодаря такой сегментации волосы насекомоядных могут сгибаться в разные стороны, не ломаясь и не свойлачиваясь, что позволяет зверькам легко передвигаться в норах не только головой вперед, но и пятиться назад. Кроме того, такая структура шерсти обусловливает удержание в ее толще так называемого инертного воздуха, придающего волосяному покрову зверьков наилучшие теплозащитные свойства.
Материалы
Материалом для наших исследований послужили шкурки зверьков, отловленных в разные сезоны 1972–2018 гг. преимущественно в центральных и южных районах Карелии.
Методы
Изучение расположения волос на шкурке, строение микроструктуры стержня отдельного волоса, его толщины, плотности и высоты (длины), дифференцировки и топографии шерсти проводили по общепринятым методикам (Кузнецов, 1932, 1952; Церевитинов, 1951; Borowski, 1952; Соколов, 1973; Соколов и др., 1986; Чернова, Целикова, 2004). Микроструктуру волоса исследовали под оптическим микроскопом при увеличении 15 х 90 (иммерсионный объектив). Отпечатки кутикулярного слоя волос делали на парфюмерном лаке №1 (Хмелевская, 1965). Определение теплоизоляционных свойств шкурок (по удельной теплопроводности) выполняли с помощью специального прибора ИТ-3, изготовленного по нашему заказу для измерения теплопроводности небольших по размеру неметаллических объектов в Киевском институте технической теплофизики АН УССР. Существенное преимущество этого прибора заключается в большой чувствительности и точности, а также в том, что все необходимые для расчета теплопроводности величины получают непосредственным измерением.
Завершая описание примененных методик, следует особо остановиться на правомерности или, скорее, уместности использования при описании архитектоники волос насекомоядных терминов «сегмент» и «перетяжка». Дело в том, что, хотя в световом микроскопе волосы землероек и кротов выглядят сегментированными, т.е. состоящими из более широких участков – сегментов и узких участков – перетяжек, в действительности это только кажущийся, чисто зрительный эффект. Если при изготовлении препарата волос случайно раскручивается, то хорошо видно, что он представляет собой сплошную постепенно расширяющуюся ленту, которая в нескольких местах поворачивается вдоль оси на 180°. Число таких поворотов строго определенное для каждой категории волос и изменяется посезонно. Впечатление же сегментированности создается за счет закономерно повторяющихся при поворотах расширений и сужений сердцевинного канала и неравномерности распределения в нем пигмента. Упоминание об этом мы находим и в ряде предшествующих публикаций (Keller, 1978; Hutterer, Hurter, 1981; Kapischke, Muhkle, 1988; Соколов, Чернова, 1998).
Результаты
Общие и видовые особенности волосяного покрова
Как показали многолетние исследования, выполненные нами на отдельных фоновых видах мелких млекопитающих тайги, и в частности обыкновенной, малой и средней бурозубках (Sorex araneus L., S. caecutience Laxm. и S. minutus L.), европейском кроте (Talpa europea L.), водяной куторе (Neomys fodiens Penn.), лесной мышовке (Sicista betulina Pall.), европейской рыжей полевке (Miodes glareolus Schr.) и полевке-экономке (Microtus oeconomus Pall.) (Ивантер, Левина, 1980, 1981; Ивантер и др., 1985; Ивантер, 2021; Ивантер, Моисеева, 2022), их волосяной покров характеризуется не только общими, характерными для всех представителей этой группы животных свойствами (дифференциация волос на три или четыре категории, трехслойность стержня, относительно небольшая длина и толщина волос, зональность окраски, своеобразие сезонной изменчивости соотношения слоев волоса и других морфологических показателей волосяного покрова), но и специфическими особенностями, имеющими важное приспособительное значение и представляющими поэтому особый интерес для сравнительно-экологических исследований. Несмотря на внешнее, во многом кажущееся сходство, в строении волосяного покрова у различных видов мелких млекопитающих обнаруживаются существенные отличия и в морфогистологии отдельного волосяного стержня, и в характере расположения в нем пигмента, и в густоте, длине и толщине волос.
Например, у всех насекомоядных волосы имеют сегментированное строение, обеспечивающие укладку волос в любую сторону, тогда как у грызунов такая сегментация отсутствует и волосы образуют «потоки» в направлении от головы к хвосту. Кроме того, если у насекомоядных волосы растут перпендикулярно к поверхности кожи и расположены поодиночке, то у грызунов они лежат наклонно и образуют группы – «пучки». Имеются отличия и в морфологии отдельного волоса, и в характере расположения в нем пигмента, и в густоте, длине и толщине волос.
Обнаруживаются характерные особенности и в микроструктуре стержня волос. Так, на концевых сегментах (граннах) остевых волос насекомоядных (в отличии от грызунов) имеются характерные продольные желобки, отчего в поперечнике волосы приобретают вид двутаврового среза. Если у насекомоядных сердцевина волоса прерывистая и однорядная, то у грызунов она сплошная и в гранне 3–4-рядная.
Окраска покрова определяется наличием и различной концентрацией меланина и липохрома (черного и рыжего пигментов). При этом липохром располагается в граннах, а меланин – в остальных участках волос. Общей же для всех исследованных видов особенностью является темная, с преобладанием бурых тонов окраска дорсальной стороны тела и светлая, грязно-белая (за исключением крота) вентральной. У большинства представителей имеется хорошо выраженная переходная зона на боках, однако у некоторых (кутора, крот) она отсутствует. В группе мелких грызунов особняком стоит лесная мышовка. Этот самый северный представитель впадающих у нас в зимнюю спячу тушканчиков выделяется особой окраской шкурки с отчетливой черной полоской на хребте, одиночным расположением волос на шкурке, а также хорошо выраженной сегментацией пуховых волос и сравнительно небольшой (наименьшей среди грызунов) толщиной волос всех категорий.
Предположение С. Боровского (Borowski, 1973) об отсутствии у мелких насекомоядных деления волос на направляющие, остевых и пуховые наши исследования не подтвердили. Отрицая у них дифференцировку волос по функционально-морфологическому принципу, С. Боровский классифицирует их по длине и степени сегментации стержня и выделяет на этой основе три типа волос: 1) летние, четырехсеменные, иногда с коротким кончиком (с пятым сильно укороченным сегментом); 2) весенние, с пятью, реже шестью сегментами; 3) зимние, шестисегментные, иногда с дополнительным седьмым концевым сегментом. Как уже указывалось (Ивантер, 2021), такую классификацию нельзя признать удачной. Волосы с «дополнительным коротким сегментом» в действительности представляют собой не что иное, как пуховые, которые по достижении нормальных размеров, подобно волосам всех других категорий, приобретают постоянное количество сегментов. Не исследуя специально луковицу волоса, вряд ли можно утверждать, как это делает С. Боровский, что пятисегментные волосы – обязательно весенние. Эти «весенние» волосы вполне могут превратиться в семисегментные «зимние»; их с таким же основанием можно назвать и «осенними», тем более что именно осенью идет увеличение числа сегментов растущих волос. Кстати, о том, из каких волос состоит осенний мех землероек, С. Боровский вообще не упоминает. Разъясняя этот вопрос, следует учитывать, что С. Боровский имел дело со сбритыми, а не с выпавшими волосами, а светлая мездра еще не доказательство того, что рост волоса уже успешно завершился. При микроскопировании такой мездры часто можно увидеть еще имеющиеся скопления пигмента в функционирующих фолликулах и открытый сердцевинный канал в прикорневой части волоса, что явно свидетельствует о росте волос. Кроме того, на шкурке одной и той же особи в межсезонные периоды (весна, осень) могут одновременно присутствовать (если пользоваться классификацией С. Боровского) и зимние, и весенние, и летние волосы: одни еще не выпали, другие на стадии роста, а третьи закончили рост.
Линейные размеры, число сегментов, форма и микроструктура позволяют выделить у исследованных насекомоядных (крота, куторы, землероек-бурозубок) четыре категории волос: направляющие, остевые I и II типов и пуховые, а у грызунов (лесной мышовки, рыжей полевки и полевки-экономки) – три: направляющие, остевые и пуховые.
Направляющий волос – самый длинный, толстый и упругий, возвышающийся над остальными и хорошо заметный даже невооруженным глазом при рассматривании шкурки на свет (рис. 1). Стержень у него почти прямой, веретенообразный, в поперечном сечении округлый, основание его не извитое, сегменты и перетяжки отсутствуют, концевая гранна хорошо развита и сильно растянута вдоль всего волоса, занимая более половины его длины. К вершине направляющий волос постепенно утончается и образует заостренный нитевидный и почти бесцветный кончик; уменьшается толщина стержня и по направлению к корню. Благодаря проходящим по обеим или одной из сторон гранны продольным желобкам (первое характерно для насекомоядных, второе – для грызунов) поперечный срез направляющего волоса имеет либо вид двутаврового рельса (крот, землеройки), либо форму овала с отчетливым боковым углублением (полевки). Окраска волос рассматриваемого типа темная, однотонная, прикорневая часть и кончик бесцветны. Количество направляющих волос в мехе насекомоядных незначительное: 1–2 на 200–300 остевых и пуховых. У грызунов их несколько больше: 3–4 % от общего числа волос.
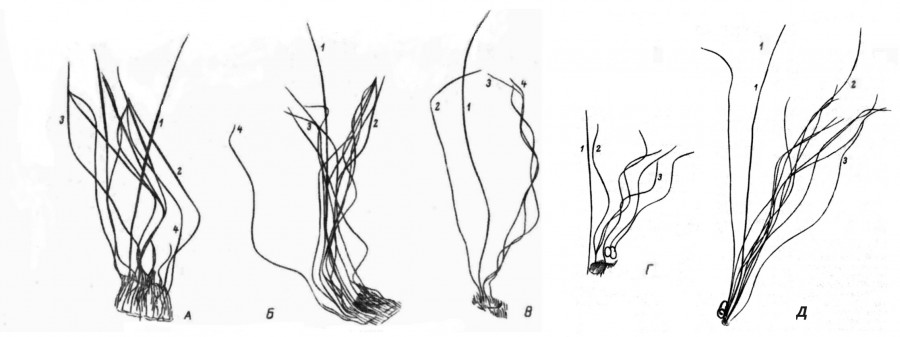
Рис. 1. Летние волосы мелких насекомоядных млекопитающих: обыкновенной бурозубки (А), куторы (Б), крота (В), лесной мышовки (Г) и европейской рыжей полевки (Д); 1 – направляющий; 2 – остевой I; 3 – пуховой; 4 – остевой II
Fig. 1. Summer hair of small insectivorous mammals: common brown-toothed (A), cooter (Б), mole (В), forest mouse (Г) and European red vole (Д); 1 – guide; 2 – guard I; 3 – down; 4 – guard II
В функциональном отношении направляющие волосы, очевидно, связаны с осязанием. Как указывает Б. Ф. Церевитинов (1951), «сумки направляющих волос по сравнению с сумками других волос залегают в более глубоких слоях дермы, где сильнее развито нервное сплетение. Поэтому малейшее давление на стержень направляющего волоса как на длинный и упругий рычаг передается корню и оплетающим волосяную сумку нервным окончаниям. Прямая неизогнутая форма направляющего волоса способствует лучшему восприятию различных раздражений. Но их нельзя сравнить с настоящими осязательными волосами – вибриссами, сумки которых снабжены нервными окончаниями и окружены венозными синусами» (Церевитинов, 1951, с. 8). Соглашаясь с тем, что направляющие волосы участвуют в осязании животного, Б. А. Кузнецов (1952) считает, что они выполняют также и защитную функцию: вместе с остевыми поддерживают массу пуховых волос, образуя как бы каркас мехового покрова и предохраняя его от свойлачивания.
Остевые волосы несколько короче и тоньше направляющих и характеризуются ланцетовидной формой с хорошо развитой уплощенной гранной. У насекомоядных млекопитающих стержень ости отчетливо сегментирован, имеет вид ленты, продольно повернутой в местах сужения сердцевинного канала на 180°, и состоит из нескольких расширенных сегментов, разграниченных «перетяжками», образующими изгибы ости под тупым углом. Наиболее развит концевой сегмент (гранна), он длиннее и шире остальных и имеет по бокам две продольные бороздки, благодаря чему остевой волос очень напоминает по форме сплющенный и продольно повернутый в нескольких местах железнодорожный рельс. В кончике волоса и в местах сгиба и поворота эти бороздки замкнуты. Как выяснилось, такое строение характерно для остевых волос всех изученных видов насекомоядных млекопитающих и может служить достаточно четким таксономическим признаком данного отряда. Напомним, что у грызунов продольных желобков или вовсе нет, или имеется лишь один, придающий поперечному срезу остевого волоса характерную бобовидную форму с глубокой выемкой на одной из сторон.
Окраска остевых волос зональная. В концевой гранне сконцентрирован относительно светлый ржаво-коричневый пигмент, в нижележащих частях темный, черно-бурый, а в перетяжках он или почти совсем отсутствует (насекомоядные), или темно-серого, почти черного цвета (полевки).
У грызунов мы выделяем один тип остевых волос, а у насекомоядных – два, отличающиеся друг от друга линейными размерами и числом сегментов. У грызунов сегментации волос нет, но перед гранной и в основании волоса имеется сужение сердцевины, правда, обычно без нарушения ее клеточной структуры (клетки лишь вытягиваются), столь типичного для представителей насекомоядных. Бесцветный (депигментированный) кончик остевого волоса как у насекомоядных, так и у грызунов гораздо короче, чем направляющих. Отличаются они и по количеству: на 4 мм² шкурки у различных видов мелких млекопитающих насчитываются от 50–100 до 300–400 остевых, т.е. в несколько десятков раз больше, чем направляющих.
Основная функция остевых волос сводится к защите более нежных пуховых от механических повреждений, предохранению их от свойлачивания и амортизации меха в целом. Кроме того, отгибаясь и прикрывая подпушь, утолщенные концевые сегменты остевых волос способствуют образованию в мехе теплозащитной воздушной прослойки и тем самым обеспечивают животному более совершенную терморегуляцию.
Пуховые волосы — самые тонкие и короткие и выполняют в основном термоизоляционную функцию. У насекомоядных они отчетливо сегментированы, а у грызунов лишь волнообразно изогнуты. Их цилиндрические стержни обычно имеют примерно одинаковую толщину по всей длине волоса и в отличие от остевых характеризуются слабо развитой, укороченной и тонкой концевой гранной. Зато характер распределения пигмента точно такой же, как у остевых волос: концевые гранны окрашены светлее остальных и в более рыжие тона, нижележащие участки содержат окисленный меланин и имеют более темный, черно-бурый цвет, в перетяжках же пигмент или отсутствует, тогда они бесцветны (у землероек и крота), или, наоборот, сконцентрирован в том же количестве, что и в ножке волоса, что придает этим участкам интенсивный, почти черный цвет (грызуны). В количественном отношении пуховые волосы – самые многочисленные: на 4 мм² шкурки их насчитывается от 200 до 350, что составляет 60–90 % от общего числа волос всех категорий.
Завершая общую характеристику выделяемых нами трех основных типов волос, следует подчеркнуть, что они крайне неоднородны и сильно варьируют в пределах каждого типа. Особенно изменчив остевой волос насекомоядных млекопитающих, в нем отчетливо выделяются по длине, толщине и числу сегментов две морфологические категории: ости I и II порядка. Несколько меньше подвержен вариации пуховой волос, а самые однородные – направляющие. Что же касается соотношения волос разных категорий, то оно существенно различается не только у разных видов животных, но и у одной и той же особи на разных участках шкурки и в разные сезоны.
Вместе с тем в сравнении с другими млекопитающими, особенно хищными, зайцеобразными и некоторыми крупными видами грызунов, дифференциация волосяного покрова выражена у мелких насекомоядных и полевок в гораздо меньшей степени. Это, очевидно, объясняется не только их более древним происхождением и приспособлением к подземному образу жизни, но и приобретенной в процессе эволюции адаптивной универсализацией (совмещением) функций волос разных категорий (все они в более или менее равной степени участвуют и в механической, и в тепловой защите тела). Полифункциональность волос привела к улучшению теплоизоляционных и механических качеств меха, но без увеличения его толщины и общей массы, совершенно не совместимого с активным передвижением мелких животных.
В микроструктуре стержня волоса выявляется три четко отграниченных концентрических слоя: кутикулярный (или чешуйчатый), корковый и сердцевинный (рис. 2). Образующая наружную оболочку волоса кутикула построена из одного слоя ороговевших чешуйчатых клеток некольцевидного типа со свободным краем, направленным к вершине волоса. Лишенные пигмента полупрозрачные чешуйки кутикулы на протяжении волоса закономерно меняются по форме и размерам. В прикорневой зоне и в местах перетяжек стержня они сильно удлинены, имеют у насекомоядных ровные, округлые, а у грызунов – зубчатые края и, относительно неплотно налегая друг на друга, обеспечивают необходимую гибкость волоса. По мере расширения стержня в гранну клетки кутикулы становятся короче и на концевой гранне превращаются в очень короткие и широкие чешуйки, зазубренные по краям и так плотно охватывающие стержень волоса, что не позволяют ему сгибаться. Чем толще гранна, тем шире и короче чешуйки и лучше выражена зазубренность края. У грызунов чешуйчатые клетки в области гранны приобретают вид узких, слегка волнистых лент, плотно опоясывающих стержень и придающих ему своеобразную поперечную исчерченность.
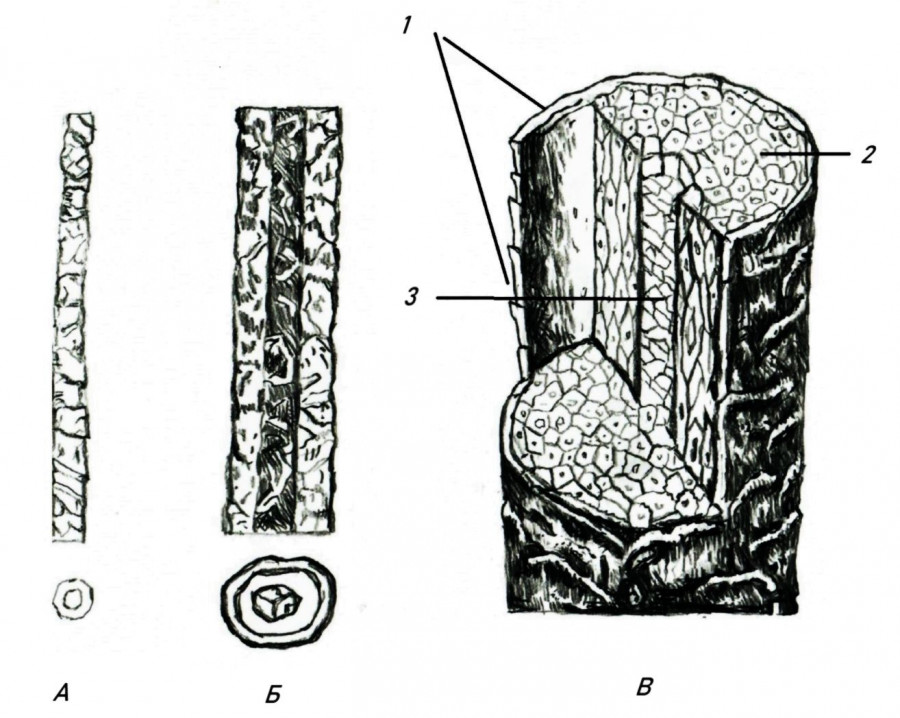
Рис. 2. Схема гистологической структуры волос мелких млекопитающих: А – пух, Б – ость, В – общее строение волосяного стержня: 1 – чешуйчатый слой; 2 – корковый слой; 3 – сердцевина
Fig. 2. Diagram of the histological structure of the hair of small mammals: A – fluff, Б – awn, В – the general structure of the hair shaft: 1 – scaly layer; 2 – cortical layer; 3 – core
Как показали проведенные исследования, в пределах одного вида характер рисунка кутикулы зимнего и летнего волоса отличается большим постоянством. Здесь надо помнить о том, что для мелких млекопитающих, выбранных нами в качестве объекта изучения, сезонная изменчивость показателей является одновременно и возрастной (в связи с коротким жизненным циклом зверьков). Сравнивая кутикулу летних и зимних волос насекомоядных и грызунов, удалось отметить лишь некоторые сезонные отличия в форме и размерах чешуек при сохранении общего типа рисунка. Чешуйки кутикулы зимних волос землероек несколько менее однородны, чем у летних, имеется значительное количество удлиненных чешуек. На более короткие чешуйки летних волос у ондатры указывает Е. А. Павлова (1955), у нутрии – Е. В. Фадеев (1955), у белки – Н. В. Хмелевская (1965). Видимо, сезонные изменения в строении кутикулы можно обнаружить у видов с довольно резкими сезонными различиями в общем характере всего волосяного покрова. Однако и в этих случаях тип строения кутикулы остается неизменным, варьирует главным образом лишь длина чешуек.
Аналогичный характер имеет возрастная изменчивость кутикулы. Она также невелика по масштабам и сводится в основном к различиям в размерах чешуйчатых клеток. Так, К. Люринг (Liihring, 1928) при сопоставлении кутикулы волос взрослой и новорожденной белок вообще не нашел отличий. В то же время у ондатры и нутрии молодые животные имеют более короткие чешуйки кутикулы, чем взрослые (Павлова, 1955; Фадеев, 1955). По Б. Бергерсену (Bergersen, 1931), кутикула волос новорожденных тюленей значительно отличается от таковой взрослых: свободные края чешуек у них более гладкие и высота клеток несколько больше, чем у взрослых.
Сравнивая кутикулу волос представителей разных систематических групп, обитающих в близких условиях, мы не находим каких-либо общих черт, характерных для того или иного экологического (адаптивного) типа. В то же время значительно отличающиеся по биологии, но близкие в систематическом отношении роды и виды имеют сходное строение кутикулы. Например, рисунок кутикулы волос типичных наземных животных, землероек-бурозубок и лесных полевок, во многом различен, тогда как при сравнении кутикулы крота, ведущего подземный образ жизни, куторы – типичного полуводного животного и любого вида бурозубок прежде всего заметны общие для всех насекомоядных черты рисунка, в частности правильная форма чешуек с округлым, ровным свободным краем.
Согласно Н. В. Хмелевской (1965), рисунок кутикулы волос водных грызунов – ондатры и бобра – совершенно различен, тогда как кутикула волос ондатры и относящейся к тому же подсемейству серой полевки имеет общие для полевок черты рисунка. Точно так же не удалось обнаружить сходства в строении кутикулы волос у далеких в систематическом отношении, но близких по образу жизни пустынных и подземных грызунов. Это говорит о том, что строение кутикулы волоса, по-видимому, не имеет непосредственного приспособительного значения и находится в коррелятивной зависимости от каких-то общих морфологических особенностей.
Многие авторы считают, что строение кутикулы специфично в основном не для вида, а для рода (Brown, 1942; Williamson, 1951; Benedict, 1957) или даже семейства (Cole, 1924). Ж. Вильямсон (Williamson, 1951), исследуя волосы оленей, выяснил, что отдельные виды по строению кутикулы не отличаются, но представители каждого рода имеют вполне определенный рисунок кутикулярных чешуй. А. Бенедикт (Benedict, 1957) при изучении волос летучих мышей всей мировой фауны установил, что 14 видов космополитически распространенного рода Myotis имеют сходное строение волос, тогда как виды разных родов, обитающих в одних и тех же условиях, отличаются по строению кутикулы. И. В. Назарова (1958) не нашла видовых отличий в строении кутикулы волос серых полевок и считает, что разные виды рода Microtus можно отличить по особенностям строения сердцевинного слоя и по характеру пигментации.
Вместе с тем выполнен целый ряд исследований, доказывающих возможность видовой диагностики млекопитающих по рисунку кутикулы. Это, в частности, работы Смита (Smith, 1933) по землеройкам, Г. Я. Содолевской (1957) по мышам, Л. Н. Скурат (1980), В. Е. Соколова, Л. Н. Скурат и Ж. М. Никифоровой (1981) по семействам летяг и беличьих, Х. Аппельта (Appelt, 1979) по летучим мышам, А. Келлера (Keller, 1981) по семействам мышиных и хомяковых. Г. Я. Содолевская, дифференцируя два близких вида мышей рода Apodemus, выяснила, что у желтогорлой мыши ромбовидные чешуйки кутикулы прикорневой зоны остевых волос расположены по спирали, а у лесной мыши волнообразно. Однако этот вывод опровергается исследованиями Н. В. Хмелевской (1965), показавшей, что у обоих видов можно обнаружить тот и другой тип расположения чешуек даже на одном волосе в зависимости от того, по какой линии соединять вершины чешуек.
В обстоятельной работе В. Е. Соколова, Л. Н. Скурат и Ж. М. Никифоровой (1981) рассмотрены возможности использования микроструктуры кутикулы и формы поперечных срезов остевых волос для идентификации представителей семейств Pteromyidae и Sciuridae и предлагается ключ к определению животных до рода (сурки и суслики) или до вида (летяга, белка, бурундук и тонкопалый суслик). При этом авторы обращают особое внимание на то, что для более точного определения видовой принадлежности волос необходимо учитывать комплекс признаков, в частности, помимо рисунка кутикулы и формы поперечных срезов, рекомендуется использовать для этой цели промеры волос и тип сердцевины. К близким выводам приходит и А. Келлер (Keller, 1981). По морфологии, форме чешуек и рисунку поперечных срезов кутикулы им составлен видовой определитель для 16 представителей семейств Cricetidae и Muridae, в том числе рыжей, обыкновенной и темной полевок, водяной крысы, ондатры, желтогорлой, лесной и домовой мышей, серой и черной крыс, мыши-малютки и др.
Заслуживают внимания публикации, касающиеся строения волос домашних животных и их ближайших родственников. Согласно исследованию М. Сауэра (Sauer, 1922), у домашних животных корковый слой волоса шире, сердцевина уже, а чешуйки кутикулы гуще расположены, чем у их диких сородичей. Аурин (Aurin, 1933) нашел отличия в строении кутикулы не только у зайца и кролика, но и у диких и домашних кроликов. Полученные результаты свидетельствуют о том, что под влиянием одомашнивания, наряду с общим увеличением пластичности и изменчивости всего организма животного, очевидно, увеличивается и степень изменчивости тонкого строения волоса.
Как уже указывалось, мы не обнаружили достаточно четких отличий в строении кутикулярного слоя волос у видов рода Sorex. Все они имеют вполне определенный, но очень схожий рисунок кутикулы, не позволяющий с уверенностью дифференцировать их до вида. К аналогичному выводу относительно родов Citellus и Ochotona приходит Н. В. Хмелевская (1965). Тем не менее это не противоречит рассмотренным выше исследованиям, констатирующим возможность видовой диагностики представителей одного и того же рода млекопитающих по строению кутикулы. Просто то, что нереально для очень близких, морфологически и экологически сходных форм, относящихся к слабо дивергирующему роду, оказывается вполне возможным, когда требуется разграничить виды, более далекие и хорошо различающиеся по строению и образу жизни. Зато представители разных родов, а тем более семейств и отрядов дифференцируются по морфологии волос достаточно четко. Правда, и здесь для более точного выявления отличий необходимо, наряду с микроструктурой кутикулы, учитывать и другие особенности строения волос: их форму, размеры, характер сегментации, дифференциацию на категории, расположение на шкурке, соотношение слоев стержня, распределение пигмента, тип сердцевины и т.д.
Между кутикулярным и сердцевинным слоями располагается довольно мощный корковый слой, состоящий из веретеновидных, плотно соединенных между собой кератинизированных клеток и обеспечивающий прочность волоса на разрыв. Он хорошо просматривается по всей длине волоса и образует трубчатый футляр центрального канала. У насекомоядных млекопитающих корковый слой лишен пигмента, а у грызунов содержит гранулы меланина и наряду с сердцевиной определяет окраску волос. Кроме того, у грызунов он относительно более тонкий и равномернее распределен по длине стержня.
Окраска и толщина волоса зависят в основном от развития и строения сердцевинного слоя: от размеров и расположения слагающих его чечевицеобразных клеток, наличия пигмента, внутри- и межклеточных воздушных полостей и т.д. Диаметр сердцевины меняется по мере созревания волоса и неодинаков в различных его частях. Наибольшего развития этот слой достигает в верхних граннах, где рыхло расположенные клетки сердцевины содержат многочисленные пигментные зерна, лежащие в виде монетных столбиков и обусловливающие окраску волос. В самой широкой части концевой граны клетки сердцевины расположены в 2–3 (у насекомоядных) или даже 4–6 рядов (у грызунов), а по направлению к вершине и основанию волоса (и ближе к перетяжке) сердцевина постепенно переходит в двухрядную и затем однорядную. В тонких, изогнутых и покрытых самыми длинными чешуйками частях волоса насекомоядных (между сегментами и в прикорневой части растущего волоса) сердцевина очень узкая, нитевидная, местами прерывистая и с диффузным распределением пигмента. В то же время у грызунов она обычно сохраняет клеточную структуру и в области шейки, хотя, конечно, и здесь сильно истончена и состоит из одного ряда клеток. В самом кончике и прикорневой части зрелого волоса сердцевина и пигмент практически отсутствуют, и, наоборот, сердцевинный слой очень хорошо развит и богат пигментом в корне и прикорневой части растущего волоса, особенно в начальный период роста, когда волосы находятся еще в толще кожи. Появившийся в основании волоса нитевидный, постепенно расширяющийся сердцевинный канал сначала не имеет четкой клеточной структуры и характеризуется диффузным распределением пигментных зерен, а затем в нем становится все более заметно строгое чередование воздушных полостей и заполненных пигментом клеток, достигающих наибольшего развития в концевых граннах волоса.
Таковы наиболее характерные особенности микроструктуры стержня волос млекопитающих. Однако в рамках этой общей схемы наблюдаются многочисленные вариации и отклонения, связанные, в частности, с типом (категорией) волоса, степенью его зрелости, местоположением на шкурке, сезоном года, видом животного. При этом наиболее изменчиво строение сердцевины. Так, если у направляющих и остевых волос она одно-двухрядная в наиболее тонкой части и многорядная в гранне, то у пуховых либо вовсе не выражена, либо прерывиста и однорядна на всем протяжении стержня и, кроме того, содержит большое количество воздуха. Различно у волос разных категорий и соотношение коркового и сердцевинного слоев. По развитию коркового слоя на первом месте стоят остевые, затем следуют направляющие и пуховые, сердцевина же, наоборот, лучше выражена у пуховых. Еще заметнее сезонные изменения. Как правило, для зимних волос по сравнению с летними характерны относительно более толстая сердцевина и более тонкий корковый слой. При этом зимнее утолщение сердцевинного канала происходит в основном за счет увеличения воздушных полостей, что ведет к улучшению термозащитных качеств шерсти.
О возможностях использования рисунка кутикулы для определения видов и родов млекопитающих достаточно подробно говорилось выше. Что же касается сердцевинного слоя, то его строение менее видоспецифично и для диагностических целей вряд ли пригодно. Тем не менее выявляются достаточно четкие отличия между представителями разных отрядов. Например, для остевых волос грызунов характерна многорядность сердцевины в граннах и сохранение ее клеточной структуры в перетяжках, тогда как для насекомоядных – однорядность сердцевины, а в местах сужений – нарушение структуры вплоть до исчезновения центрального канала (прерывистая сердцевина).
Один из наиболее изменчивых показателей структуры меха – плотность или густота волос. По этому признаку четко дифференцируются как отдельные группы, так и различные виды одной и той же группы животных (табл. 1). Мех насекомоядных как зимой, так и летом гуще, чем у грызунов. Среди Insectivora наиболее густым мехом обладает крот, плотность волос у которого (летом – 369–521, зимой – 538–729 на 4 мм²) в 1.5 раза больше, чем у малой бурозубки (соответственно 266–364 и 396–463). Наиболее редкий мех оказался у рыжей полевки – 225–300 летом и 284–422 зимой, зато лесная мышовка по густоте летнего меха превосходит даже крота – 382–576 на 4 мм².
Таблица 1. Плотность волосяного покрова мелких млекопитающих Карелии
| Вид | Сезон | n | Спина | Бок | Брюшко |
| Обыкновенная бурозубка | Лето | 25 | 442.7 ± 7.3 | 395.6 ± 7.9 | 479.8 ± 4.6 |
| Зима | 25 | 523.1 ± 7.2 | 429.6 ± 4.4 | 565.4 ± 9.0 | |
| Средняя бурозубка | Лето | 25 | 389.9 ± 6.8 | 376.3 ± 7.7 | 332.8 ± 7.6 |
| Зима | 25 | 538.5 ± 8.9 | 432.7 ± 8.7 | 460.9 ± 9.1 | |
| Малая бурозубка | Лето | 14 | 364.0 ± 6.7 | 328.3 ± 6.0 | 266.4 ± 4.3 |
| Зима | 15 | 463.4 ± 5.0 | 413.1 ± 5.1 | 395.9 ± 4.7 | |
| Обыкновенная кутора | Лето | 18 | 467.4± 6.1 | 350.9 ± 5.3 | 368.1 ± 5.0 |
| Европейский крот | Лето | 16 | 521.0 ± 9.6 | 368.9 ± 9.2 | 457.9 ± 5.9 |
| Зима | 15 | 728.7 ± 9.8 | 538.1 ± 9.1 | 572.0 ± 9.2 | |
| Лесная мышовка | Лето | 18 | 576.3 ± 8.3 | 495.7 ± 3.1 | 381.9 ± 7.0 |
| Рыжая полевка | Лето | 15 | 299.9 ± 3.0 | 288.8 ± 3.1 | 225.4 ± 1.8 |
| Зима | 13 | 422.5 ± 4.6 | 399.2 ± 4.1 | 284.7 ± 2.1 | |
| Полевка-экономка | Лето | 15 | 334.0 ± 2.9 | 288.0 ± 1.9 | 317.6 ± 2.1 |
| Зима | 15 | 478.7 ± 6.1 | 453.6 ± 5.6 | 460.2 ± 5.8 |
Несмотря на относительную равномерность распределения волос по телу зверька, густота их на различных участках шкурки неодинакова (см. табл. 1). При этом у большинства изученных видов наибольшей густотой отличается мех на спине. Что же касается топографии плотности меха на остальных частях тела, то она зависит от экологии вида и, прежде всего, от условий обитания: у крота наиболее редкие волосы растут на боках, у бурозубок и полевок – на брюшке, у куторы существенных отличий в опушении боков и брюшка не обнаружено.
Более заметны сезонные изменения, в частности, изменения зимнего покрова: он в 1.2–1.4 раза гуще летнего, причем плотность его варьирует в основном за счет пуховых волос, тогда как количество остевых и направляющих более постоянно. Наибольшей густотой отличается мех линяющих животных, у которых еще не выпавшие прочно удерживающиеся в коже старые волосы соседствуют с интенсивно подрастающими новыми. Зависит плотность меха и от возраста животных. Подтверждение этому – обратная связь между рассматриваемым показателем и размерами тела зверьков, говорящая об уменьшении густоты волос с ростом особи.
Толщина волоса (табл. 2), соотношение его слоев и общие особенности строения стержня у разных видов животных неодинаковы (рис. 3). Наиболее толстые и грубые волосы характерны для полевок (см. табл. 2). Они у них к тому же наиболее непрочные на разрыв, что объясняется относительно слабым развитием коркового слоя, составляющего вместе с кутикулой 6.5–14.3 % от общей толщины стержня. У насекомоядных этот показатель возрастает до 17–25 %. Самые тонкие остевые волосы у лесной мышовки (17.4 мкм), причем на долю сердцевины у нее приходится 79.9 % общей толщины стержня. У грызунов обращает внимание относительное постоянство диаметра сердцевинного канала при значительной сезонной изменчивости общей толщины волоса. Между тем у насекомоядных в разные сезоны года изменение этих показателей происходит асинхронно. Так, летом с увеличением общей толщины волоса в нем наблюдается сужение сердцевины, а зимой, наоборот, утончение стержня сопровождается относительным утолщением мозгового слоя.
Таблица 2. Толщина волос спины мелких млекопитающих Карелии
| Остевые | Пуховые | ||||||
| Вид | Сезон | n | толщина волоса, мкм | относительная толщина сердцевины, % от общей толщины волоса | n | толщина волоса, мкм | относительная толщина сердцевины, % от общей толщины волоса |
| Обыкновенная бурозубка | Лето | 25 | 40.0 ± 0.4 | 58 | 25 | 12.6 ± 0.1 | 63 |
| Зима | 20 | 30.1 ± 0.6 | 63 | 24 | 7.2 ± 0.3 | 77 | |
| Средняя бурозубка | Лето | 50 | 31.1 ± 0.6 | 64 | 28 | 11.0 ± 0.8 | 62 |
| Зима | 28 | 27.0 ± 0.4 | 62 | 26 | 7.9 ± 0.3 | 79 | |
| Малая бурозубка | Лето | 25 | 27.6 ± 0.4 | 55 | 29 | 10.0 ± 0.3 | 61 |
| Зима | 24 | 23.8 ± 0.2 | 65 | 33 | 6.1 ± 0.3 | 80 | |
| Обыкновенная кутора | Лето | 16 | 27.6 ± 0.5 | 50 | 18 | 9.8 ± 0.1 | 72 |
| Обыкновенный крот | Лето | 25 | 32.4 ± 0.3 | 51 | 19 | 10.9 ± 0.2 | 54 |
| Зима | 17 | 27.8 ± 0.5 | 66 | 25 | 9.7 ± 0.2 | 70 | |
| Лесная мышовка | Лето | 20 | 17.4 ± 0.2 | 80 | 18 | 15.8 ± 0.2 | 81 |
| Рыжая полевка | Лето | 22 | 49.1 ± 0.3 | 80 | 20 | 28.1 ± 0.2 | 78 |
| Зима | 18 | 20.1 ± 0.2 | 71 | 18 | 11.9 ± 0.2 | 80 | |
| Полевка-экономка | Лето | 10 | 49.1 ± 0.6 | 87 | 10 | 22.8 ± 0.7 | 54 |
| Зима | 9 | 48.6 ± 0.6 | 88 | 9 | 16.4 ± 0.3 | 82 | |
Примечание. У насекомоядных сравниваются остевые I порядка.
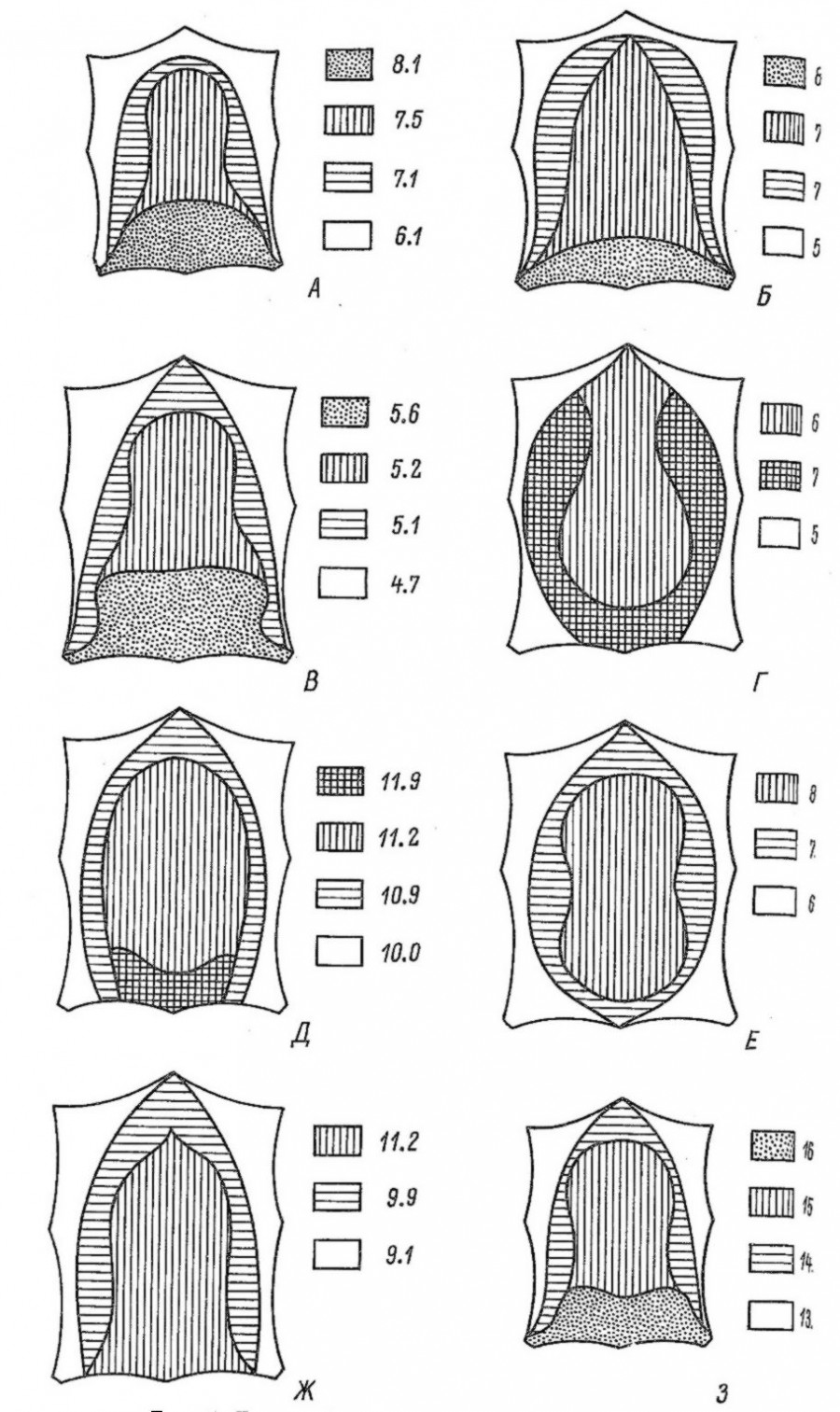
Рис. 3. Топография высоты волосяного покрова мелких млекопитающих (мм); А – обыкновенная бурозубка; Б – средняя бурозубка; В – малая бурозубка; Г – кутора; Д – крот; Е – лесная мышовка; Ж – рыжая полевка; З – полевка-экономка
Fig. 3. Topography of the hairline height of small mammals (mm); A – common shrew; Б – masked shrew; В – pygmy shrew; Г – kutora; Д – mole; Е – northern birch mouse; Ж – red-backed vole; З – root vole
У большинства исследованных видов наиболее толстые волосы встречаются на спине, а наиболее тонкие – на брюшке. Исключение составляют кутора и крот: у них на брюшке волосы несколько толще, чем на боках и спине, что объясняется, видимо, спецификой их образа жизни. Из отдельных категорий волос наибольшей толщиной обладают направляющие. Несколько уступают им по этому показателю остевые, а самые тонкие – пуховые волосы. В то же время диаметр сердцевинного канала изменяется в обратном порядке: максимальной относительной величины он достигает у пуховых, средней – у остевых, минимальной – у направляющих. Таким образом, чем тоньше волос, тем относительно большую долю занимает в нем сердцевинный слой и меньшую корковый.
Длина летних остевых волос рассматриваемых видов мелких млекопитающих (табл. 3) колеблется от 3.3 (малая бурозубка) до 12.2 мм (полевка-экономка). Наибольшей длиной характеризуются волосы полевок, их мех в 2 раза выше, чем у бурозубок, в 1.4 раза превосходит мех куторы и в 1.2 раза крота. Неодинакова длина волос и на отдельных участках шкурки. Согласно номенклатуре Б. Ф. Церевитинова (1958), изученные нами виды насекомоядных и грызунов по степени опушенности отдельных частей тела могут быть отнесены к следующим пяти типам топографии волосяного покрова: сакральному (полевка-экономка), дорсальному (рыжая полевка и лесная мышовка), сакро-эквальному (бурозубки), эквалатеральному (кутора) и эквальному (крот). Для первого из них, присущего в основном наземным млекопитающим, типично уменьшение длины волос от крестца к передней части тела. При дорсальном типе наиболее высокий мех расположен на спине, откуда его длина уменьшается в направлении головы, брюшка и крестца; это свойственно большинству настоящих норных животных. Сакро-эквальный тип характеризуется более или менее равномерным распределением волос по телу зверька с некоторым увеличением их длины в области крестца, а эквалатеральный – постепенным уменьшением высоты меха с крестца и боков на спину и брюшко (последнее типично для полуводных млекопитающих). Наконец, эквальный тип волосяного покрова, свойственный подземным формам, отличается относительно равномерной высотой меха по всему телу.
Таблица 3. Длина волос мелких млекопитающих Карелии
| Вид | Сезон | Направляющие | ||
| спина | бок | брюшко | ||
| Обыкновенная бурозубка | Лето Зима |
5.1 ± 0.06 (17) 8.5 + 0.09 (16) |
4.7 ± 0.03 (16) 8.5 ± 0.06 (15) |
4.4 ± 0.04 (18) 7.9 ± 0.09 (17) |
| Средняя бурозубка | Лето Зима |
5.9 ± 0.03 (67) 8.5 ± 0.09 (25) |
5.4 ± 0.02 (50) 8.0 ± 0.06 (20) |
4.9± 0.01 (25) 7.9± 0.04 (18) |
| Малая бурозубка | Лето Зима |
4.0 ± 0.02 (18) 5.5 ± 0.01 (17) |
4.5 ± 0.02 (16) 5.7 ± 0.01 (16) |
3.5 ± 0.02 (17) 4.9 ± 0.01 (19) |
| Обыкновенная кутора | Лето | 8.0 ± 0.02 (17) |
8.5 ± 0.08 (16) |
7.5 ± 0.08 (14) |
| Обыкновенный крот | Лето Зима |
8.5 ± 0.04 (18) 11.8 ± 0.07 (16) |
8.3 ± 0.06 (16) 11.2 ± 0.06 (14) |
7.1 ± 0.03 (15) 10.9 ± 0.08 (15) |
| Лесная мышовка | Лето | 9.0 ± 0.02 (26) |
8.5 ± 0.12 (25) |
7.8 ± 0.03 (25) |
| Рыжая полевка | Лето Зима |
12.2 ± 0.04 (18) 13.1 ± 0.02 (20) |
11.8 ± 0.02 (16) 12.7 ± 0.04 (20) |
10.2 ± 0.03 (18) 11.4 ± 0.02 (22) |
| Полевка-экономка | Лето Зима |
14.6 ± 0.04 (9) 17.4 ± 0.07 (10) |
13.3 ± 0.06 (10) 15.8 ± 0.04 (10) |
12.3 ± 0.08 (9) 14.9 ± 0.06 (10) |
| Остевые | ||||
| спина | бок | брюшко | ||
| Обыкновенная бурозубка | Лето Зима |
4.2 ± 0.01 (25) 7.5 ± 0.08 (20) |
4.3 ± 0.04 (24) 7.1 ± 0.05 (16) |
3.8± 0.04 (25) 6.3± 0.07 (24) |
| Средняя бурозубка | Лето Зима |
4.0 ± 0.05 (50) 7.5 ± 0.07 (28) |
4.0 ± 0.04 (60) 7.7 ± 0.04 (29) |
3.7 ± 0.03 (50) 5.2 ± 0.05 (26) |
| Малая бурозубка | Лето Зима |
3.3 ± 0.01 (25) 5.2 ± 0.02 (24) |
3.2 ± 0.01 (25) 5.1 ± 0.01 (25) |
2.9 ± 0.01 (24) 4.7 ± 0.01 (25) |
| Обыкновенная кутора | Лето | 6.5 ± 0.11 (25) |
7.0 ± 0.06 (25) |
5.9 ± 0.07 (18) |
| Обыкновенный крот | Лето Зима |
7.9 ± 0.02 (25) 11.2 ± 0.05 (19) |
7.7 ± 0.01 (18) 10.9 ± 0.04 (18) |
6.4 ± 0.04 (18) 10.0 ± 0.06 (19) |
| Лесная мышовка | Лето | 8.1 ± 0.09 (25) |
7.1 ± 0.09 (25) |
6.7 ± 0.04 (25) |
| Рыжая полевка | Лето Зима |
8.4 ± 0.03 (22) 11.2 ±0.03 (18) |
8.0 ± 0.03 (26) 9.9 ± 0.03 (22) |
7.5 ± 0.02 (28) 9.1 ± 0.03 (26) |
| Полевка-экономка | Лето Зима |
12.2 ± 0.05 (14) 15.1 ± 0.05 (9) |
11.9 ± 0.05 (11) 14.7 ± 0.05 (10) |
10.1 ± 0.06 (9) 13.8 ± 0.06 (16) |
| Пуховые | ||||
| спина | бок | брюшко | ||
| Обыкновенная бурозубка | Лето Зима |
3.6 ± 0.02 (25) 6.2 ± 0.03 (24) |
3.4 ± 0.01 (26) 5.9 ± 0.02 (21) |
3.3 ± 0.01 (23) 4.6 ± 0.02 (22) |
| Средняя бурозубка | Лето Зима |
3.5 ± 0.02 (25) 5.2 ± 0.03 (26) |
3.3 ± 0.01 (25) 5.0 ± 0.01 (25) |
3.1 ± 0.01 (40) 3.8 ± 0.02 (23) |
| Малая бурозубка | Лето Зима |
2.8 ± 0.04 (29) 4.2 ± 0.02 (33) |
2.8 ± 0.04 (27) 4.0 ± 0.04 (25) |
2.4 ± 0.01 (25) 3.6 ± 0.05 (27) |
| Обыкновенная кутора | Лето | 6.0 ± 0.02 (18) |
6.2 ± 0.03 (24) |
5.3 ± 0.02 (25) |
| Обыкновенный крот | Лето Зима |
6.5 ± 0.02 (29) 10.1 ± 0.02 (25) |
6.3± 0.03 (25) 10± 0.01 (20) |
5.1 ±0.01 (25) 8.9 ±0.02 (25) |
| Лесная мышовка | Лето | 7.5 ± 0.11 (27) |
6.5 ± 0.08 (25) |
5.7 ± 0.04 (24) |
| Рыжая полевка | Лето Зима |
7.3 ± 0.02 (20) 9.4 ± 0.03 (18) |
7.0 ± 0.04 (22) 9.1 ± 0.03 (28) |
6.6 ± 0.02 (32) 7.9 ± 0.02 (24) |
| Полевка-экономка | Лето Зима |
10.9 ± 0.04 (16) 12.3 ± 0.07 (9) |
10.1 ± 0.03 (15) 11.9 ± 0.05 (10) |
8.6 ± 0.06 (10) 11.0 ± 0.02 (14) |
Хорошо выражены сезонные изменения данного показателя: зимний мех по всем категориям волос на 1.5–3.5 мм длиннее летнего (см. табл. 3). У направляющих волос этот прирост составляет на разных участках шкурки и у разных видов зверьков 0.9–3.8, в среднем 2.3 мм, или 7.4–80.9 %, в среднем 30 % от летних показателей, у остевых – 1.6–3.7, в среднем 2.7 мм (21.3–92.5, в среднем 42.4 %), у пуховых – 0.7–3.8, в среднем 2.0 мм (12.8–74.5, в среднем 36.6 %). Таким образом, от лета к зиме длина направляющих волос увеличивается в среднем почти на треть, остевых – на 42, а пуховых – на 37 %. Что же касается видовых особенностей этого процесса, то они сводятся в основном к его большей интенсивности у насекомоядных (особенно крота и обыкновенной бурозубки) по сравнению с полевками.
Теплозащитные свойства покровов и механизмы терморегуляции
В процессе эволюции и освоения среды обитания мелкие млекопитающие таежного Севера приобрели целый ряд адаптаций, направленных в первую очередь на наиболее целесообразное, с наименьшими энергетическими затратами поддержание теплового баланса организма со средой. Поскольку сохранение тепла за счет физической терморегуляции не требует дополнительных пищевых ресурсов, оно является основным способом экономии энергетических затрат (Jansky, 1967). В связи с этим эволюция терморегуляции идет в направлении замены химических механизмов физическими (Слоним, 1952, 1964, 1971; Шилов, 1962), о чем говорит и более отчетливая выраженность физической терморегуляции у типичных субарктических и высокогорных видов (Шварц и др., 1968; Большаков, 1972).
Как известно, в системе приспособления млекопитающих к пониженным температурам важное место занимают реакции волосяного покрова. Многочисленными исследованиями показано, что теплоизоляция пропорциональна густоте и толщине мехового покрова и у мелких млекопитающих она ниже, чем у более крупных (Bonsma, Pretorins, 1943; Калабухов, 1951, 1957, 1969; Слоним, 1952; Марвин, 1966а, 1966б; Прасолова и др., 1967; Большаков, Некрасов, 1968; Уманцева, 1980; Давыдов, 1982). Чтобы иметь возможность передвигаться, мелкие животные должны обладать тонким и легким шерстным покровом, а то, что волосы у них по необходимости короче, снижает их теплоизоляционные свойства (Шмидт-Ниельсен, 1982).
Согласно современным представлениям, термозащитные качества волосяного покрова зависят главным образом от образующегося в его толще слоя «неподвижного» и «инертного» воздуха, исключающего конвекционные токи и создающего эффективную тепловую изоляцию (Соколов, 1973). Чем пышнее и выше мех, гуще и тоньше волосы, тем больше воздушная прослойка между волосками и, следовательно, лучше сохраняется тепло. Теплопроводность зависит также от воздуха, находящегося в сердцевине волос (она тем меньше, чем сильнее развита сердцевина) и в порах кожной ткани, а значит, от толщины и плотности кожи. Имеют значение и особенности строение волос – наличие продольных бороздок на стержне, его конфигурация, число сегментов, способ укладки и т.п.
Связь термоизоляционных свойств меха с морфологией и общим состоянием наружных покровов млекопитающих подтверждается экспериментальным определением коэффициента теплопроводности сухих коллекционных шкурок зверьков, добытых в разные сезоны года (табл. 4). Согласно этим данным, интегральная теплопроводность меха имеет достаточно четкие таксономические и сезонные различия, обусловленные структурными особенностями волосяного покрова. Чем длиннее, тоньше и гуще волосы и чем толще кожа, тем ниже коэффициент теплопроводности и, следовательно, выше термоизоляционные свойства шкурки. При этом шкурка с наиболее длинными и наиболее густыми волосами не обязательно обладает наименьшим коэффициентом теплопроводности (при межвидовом сравнении). Зато каждый вид в отдельности демонстрирует отчетливую сезонную изменчивость и зависимость коэффициента теплопроводности от длины, толщины и густоты волос. Мех насекомоядных млекопитающих значительно уступает меху грызунов по высоте, но превосходит его по плотности. Кроме того, характерное для насекомоядных сегментированное строение волос, предотвращая сминание и обеспечивая укладку меха в любом направлении, способствует удержанию в нем неподвижного воздуха, что положительно сказывается на термоизолирующих свойствах шкурки в целом. У куторы и крота, обладающих к тому же наиболее длинным и густым мехом, теплозащитные качества шкурки лучше, чем у бурозубок, почти в 2 раза. Среди изученных нами грызунов наиболее «теплую» шкурку имеет полевка-экономка (см. табл. 4), затем следуют лесная мышовка и рыжая полевка. В той же последовательности изменяются у названных видов показатели длины и густоты волос.
Таблица 4. Коэффициент теплопроводности (10–3 Вт/м • °К) сухих шкурок мелких млекопитающих Карелии
| Вид | Сезон | n | Пределы | М ± m |
| Обыкновенная бурозубка | Лето | 16 | 38.2–50.0 | 47.6 ± 0.8 |
| Зима | 15 | 35.0–46.4 | 37.4 ± 0.6 | |
| Осень | 12 | 33.1–41.2 | 34.7 ± 0.5 | |
| Средняя бурозубка | Лето | 16 | 50.0–60.0 | 54.0 ± 1.2 |
| Зима | 15 | 38.3–42.6 | 40.1 ± 0.5 | |
| Осень | 13 | 37.1–44.8 | 39.4 ± 0.3 | |
| Малая бурозубка | Лето | 15 | 49.0–57.2 | 53.5 ± 0.9 |
| Зима | 14 | 38.0–49.0 | 42.2 ± 0.5 | |
| Осень | 9 | 36.0–45.0 | 39.0 ± 0.4 | |
| Обыкновенная кутора | Лето | 18 | 30.0–42.1 | 32.5 ± 0.9 |
| Осень | 14 | 28.0–39.0 | 30.3 ± 0.8 | |
| Обыкновенный крот | Лето | 9 | 24.2–30.0 | 26.7 ± 0.3 |
| Зима | 8 | 19.0–22.4 | 20.5 ± 0.3 | |
| Осень | 8 | 18.9–21.0 | 19.1 ± 0.3 | |
| Лесная мышовка | Лето | 22 | 48.0–50.3 | 48.6 ± 0.3 |
| Осень | 4 | 37.9–38.2 | 38.1 ± 0.4 | |
| Рыжая полевка | Лето | 20 | 45.2–53.1 | 49.8 ± 0.9 |
| Зима | 16 | 37.0–50.0 | 41.0 ± 0.8 | |
| Осень | 14 | 36.1–42.1 | 38.7 ± 0.8 | |
| Полевка-экономка | Лето | 11 | 46.7–53.2 | 47.8 ± 0.4 |
| Зима | 5 | 34.1–38.4 | 36.5 ± 0.4 | |
| Осень | 8 | 34.1–40.9 | 36.1 ± 0.6 |
Хорошо известно, что структура волосяного покрова млекопитающих зависит от сезона года: зимний мех гуще и длиннее и, как правило, лучше сохраняет тепло, чем более легкий (редкий и короткий) летний мех. Это сказывается на уровне энергетического обмена. Тем не менее более совершенная термоизоляция организма «зимних» зверьков определяет специфику их адаптации к холоду: последняя обычно сопровождается не повышением теплопродукции, а значительным ограничением теплоотдачи (Слоним, 1964; Смирнов, 1968). Таким образом, улучшение теплоизоляционных свойств покровов млекопитающих под воздействием холода и обусловленное им снижение химической терморегуляции представляют собой общую закономерность (Слоним, 1961). И хотя это больше свойственно крупным животным, улучшение термозащитных качеств зимней шерсти и соответствующие сезонные изменения уровня теплоотдачи отмечены и у мелких млекопитающих (Марвин, 1966а, 1974; Большаков, Некрасов, 1968; Калабухов, 1969; Соколов, 1973 и др.).
Согласно нашим измерениям (см. табл. 4), коэффициент теплопроводности летнего волосяного покрова по отношению к зимнему увеличивается (а теплоизоляция снижается) у обыкновенной бурозубки на 28.2 %, средней – на 34.6, малой – на 26.8, у крота – на 30.2, у рыжей полевки – на 21.5, у полевки-экономки – на 30.9 %. Это соответствует отмеченным выше сезонным изменениям длины и густоты меха. Показатель ранговой корреляции Спирмена (rs) между длиной волос и удельной теплопроводностью шкурки составил (по всему материалу) –0.41 (t = 2.3; р > 0.95), что говорит о средней по силе, но статистически вполне достоверной связи. Еще теснее зависимость коэффициента теплопроводности от густоты меха: rs = –0.66 (при t = 3.6; р > 0.99). Коэффициент общей (совокупной) корреляции, оценивающий связь теплозащитных свойств шкурки с длиной и густотой волос составил 0/90 {р > 0.999).
Проведенные исследования убедительно показывают, что шкурки зверьков в разгар линьки не только не уступают зимним по термоизоляционным качествам, но даже превосходят их, о чем свидетельствует низкий коэффициент теплопроводности (см. табл. 4). Как уже говорилось, это объясняется значительным утолщением кожи и увеличением густоты меха за счет совмещения невыпавших еще старых волос с подрастающими новыми. Таким образом, вопреки распространенному мнению во время линьки механизмы физической терморегуляции у мелких млекопитающих не только не утрачивают своего значения, а напротив, служат важным звеном в цепи гомеостатических реакций, направленных на снижение непроизводительных трат энергии и потери тепла в сложные для жизни зверьков переходные периоды. Более глубокая, чем в другие сезоны, реакция газообмена на воздействие холода указывает на то, что осенью грызуны и землеройки еще сохраняют определенные черты физиологической адаптации к высоким летним температурам, и воздействие низких температур, усиленных повышенной влажностью воздуха, вызывает у них обостренную реакцию химической терморегуляции (Смирнов, 1968). Как считает П. К. Смирнов, такая реакция требует повышенных затрат энергии на избыточную теплопродукцию, необходимую для покрытия увеличенной потери тепла и, следовательно, для поддержания теплового баланса на уровне, обеспечивающем нормальную жизнедеятельность. Улучшение теплоизоляционных свойств наружных покровов в период линьки в значительной мере компенсирует несовершенство химической терморегуляции и служит важным приспособлением для более рационального теплообмена организма при его физиологической перестройке в условиях неустойчивого температурного режима среды.
Обсуждение
Среди изученных нами видов мелких млекопитающих имеются представители различных адаптивных типов, приспособившихся к норно-наземному (землеройки-бурозубки и грызуны, причем среди последних один вид – лесная мышовка – зимоспящий), подземному (крот) и полуводному (кутора) образу жизни. Это позволяет, рассмотрев их в сравнении, выявить наиболее характерные, носящие явно адаптивный характер видовые особенности в строении волосяного покрова и оценить их с экологических позиций.
К группе норно-наземных млекопитающих мы относим в основном так называемых «полунорников» – мелких грызунов и насекомоядных, обитающих в лесной подстилке и ходах неглубоких нор, но нередко появляющихся и на поверхности почвы. По образу жизни, среде обитания, а значит, и по строению меха, они совмещают в себе многие черты, характерные как для настоящих наземных, так и для специализированных норных млекопитающих, и представляют собой переходный адаптивный тип. Обитая в умеренной и особенно холодной климатических зонах, в условиях постоянных термических дефицитов, эти зверьки приобрели легкий, не мешающий движениям, но достаточно «теплый», с высокими термоизоляционными свойствами, меховой покров, характеризующийся умеренной густотой и длиной волос, неравномерностью опушения отдельных частей тела, булавовидной формой концевой гранны, слабой извитостью основания волоса и несколько более сильным, чем у других групп, развитием сердцевинного канала.
Сами по себе волосы, как известно, не могут быть хорошим теплоизолятором из-за относительно высокой теплопроводности слагающего их рогового вещества. Решающее значение в теплозащите, помимо «инертного» воздуха, заключенного в сердцевине волос, имеет так называемый «неподвижный» воздух, образующийся в шерстном покрове между волосами благодаря максимальному исключению конвекционных токов (Соколов, 1973). У землероек-бурозубок удержанию теплозащитного слоя воздуха способствует сама структура меха – одиночное, перпендикулярное к поверхности кожи расположение волос, их сегментация, извитость, дифференциация на категории. Разделяя пух и прикрывая его сверху, остевые и направляющие волосы не только предохраняют мех от сминания и свойлачивания, но и препятствуют выдуванию воздуха. Следовательно, обладая сравнительно слабо (в сравнении с грызунами) развитой сердцевиной волос, но зато гораздо большей их прочностью (за счет утолщения коркового слоя), насекомоядные компенсируют недостаточное количество «инертного» воздуха в сердцевине повышенным содержанием «неподвижного» воздуха в толще меха. В результате коэффициент теплопроводности их шкурок даже меньше, чем у грызунов. С другой стороны, более низкие, чем у землероек, теплозащитные свойства меха грызунов не сказываются отрицательно на их жизнедеятельности, поскольку эти животные занимают более «теплую» экологическую нишу (более постоянные ходы и норы с относительно стабильным микроклиматом, утепленные гнезда и т.д.), да и химическая терморегуляция у них, как известно, более совершенная.
Характерная для норно-наземных млекопитающих неравномерность опушения отдельных участков тела служит важным дополнительным компонентом физической терморегуляции. Область спины, подвергающаяся у этих животных наибольшему охлаждению, покрыта самым высоким и густым мехом, а на более скрытой и, следовательно, лучше защищенной от холода брюшной стороне тела волосяной покров значительно реже и ниже, чем на хребте и боках. Целесообразность такой топографии становится очевидной, если вспомнить характерную позу, принимаемую зверьком во время сна: свертываясь в клубок, он предохраняет нижнюю часть тела от охлаждения и, кроме того, создает условия, способствующие уменьшению теплоотдачи с единицы поверхности.
У рассматриваемой группы млекопитающих волосяной покров выполняет также важную функцию защиты кожи от механических повреждений. Являясь хорошим амортизатором, он смягчает давление окружающего субстрата, которое постоянно приходится испытывать мелким зверькам при продвижении в узких ходах нор, лесной подстилке, переплетении трав. При этом функция механической защиты волосяного покрова вступает отчасти в противоречие с функцией термоизоляции (Соколов, 1973). Наибольшей прочностью обладают волосы со слабо развитой сердцевиной, однако с ее уменьшением ухудшаются их термоизоляционные свойства. Разрешается это противоречие несколькими путями. Один из них заключается в увеличении густоты таких тонкосердцевинных волос, что улучшает термозащитные качества волосяного покрова без ущерба для его механических свойств. Второй путь – дифференциация волос, при которой функцию механической защиты берут на себя направляющие и остевые волосы, а терморегуляционную – пуховые. Наконец, третий путь связан с неравномерностью строения волоса по его длине. При этом особое значение для защиты меха от вытирания имеют бессердцевинные кончики волос, обладающие более высокими механическими свойствами благодаря хорошо развитому корковому слою. Большой прочностью на излом отличаются и другие зауженные участки волоса с отсутствием или слабым развитием сердцевины. В этих местах стержень, не ломаясь, легко сгибается в любую сторону, обеспечивая волосу эластичность и предохраняя его от повреждений. У остевых волос грызунов таких участков (их называют перетяжками) обычно два – у основания и перед гранной, а у землероек больше – от 3 до 9 в зависимости от вида и сезона года.
Группа подземных млекопитающих представлена в нашем исследовании обыкновенным кротом. Этот зверек великолепно приспособлен к роющему образу жизни, что находит отражение и в строении его волосяного покрова. Как и у других типичных представителей данного адаптивного типа, мех у крота относительно низкий, ровный и густой, со слабо выраженным ворсом и почти без наклона к хвосту. Такая своеобразная структура меха позволяет зверьку свободно двигаться в узких ходах нор как головой вперед, так и пятясь назад. Весьма характерна относительная выравненность волосяного покрова. Это проявляется и в однообразии опушения отдельных частей тела (эквальный тип топографии по высоте и густоте меха), и в сближении волос разных категорий по линейным размерам.
Явно адаптивный характер имеют и некоторые особенности микроструктуры волоса. Специфичное для крота относительно слабое развитие сердцевинного (и соответственно, более сильное коркового) слоя волос способствует улучшению их механических свойств. А то, что это касается преимущественно лишь категорий кроющих волос и почти не затрагивает пуховых (у последних сердцевина толще, чем у остевых и направляющих), способствует сохранению теплозащитных качеств меха. Прикрытый сверху направляющими и остевыми волосами пух испытывает незначительные механические воздействия, поэтому ухудшение механических свойств пуха за счет значительного повышения теплозащитных качеств вполне оправданно (Соколов, 1973). Если учесть, что шкурка крота обладает к тому же и более густым, высоким мехом и кожа у него толще, чем у близких в систематическом отношении наземных животных (землероек-бурозубок), то не приходится удивляться более высоким теплозащитным свойствам его наружных покровов.
Несмотря на обильную жировую смазку, повышенную прочность волос и их способность сгибаться и укладываться в любом направлении, мех крота от постоянного соприкосновения с почвой довольно быстро вытирается. Восстанавливается он в процессе особой компенсационной линьки – важной адаптивной особенности, характерной только для подземных млекопитающих.
В строении волосяного покрова полуводных (земноводных) млекопитающих (обыкновенная кутора) прежде всего обращают внимание признаки, общие с таковыми родственных форм, ведущих наземный и подземный образ жизни. Так, волосы куторы, подобно волосам других насекомоядных, располагаются поодиночке, перпендикулярно к поверхности кожи, делятся на четыре категории (направляющие, остевые I и II порядка и пуховые) и имеют сегментное строение с определенным числом сужений и продольных поворотов. Из других общих признаков можно указать на зональность окраски волос, трехслойность стержня, правильную форму кутикулярных чешуек и т. д. Вместе с тем имеются и некоторые специфические черты, обусловленные приспособлением животных к водной среде обитания.
В отличие от других насекомоядных у куторы наиболее длинные волосы растут на боках, что, по-видимому, способствует удержанию тела на поверхности и в толще воды. Такие морфологические особенности ее мехового покрова, как наличие на боковых сторонах уплощенных остевых волос глубоких продольных желобков, относительно слабое (в сравнении с другими представителями отряда) развитие сердцевины в граннах кроющих волос, утончение и удлинение волос всех категорий (последнее за счет большего числа сегментов), а также увеличенная густота и пышность шерсти, обеспечивают при плавании сохранение воздушной прослойки в подпуши, несмачиваемость меха и отсюда более совершенные термоизоляционные его качества. Своеобразен и характер линьки: сезонная смена меха проходит у куторы постепенно и несколько растянута во времени. Наконец, кутора имеет типичную для большинства водных животных контрастную окраску тела – серебристый низ и темный верх без переходной зоны на боках. Согласно Котту (Сott, 1950), это связано с характерным для обитателей поверхностных слоев воды оптическим эффектом, призванным закамуфлировать контуры тела с помощью скрадывающей противотени.
Из других признаков, характеризующих кутору как типичного представителя полуводных (земноводных) млекопитающих, можно назвать более четкую дифференциацию волос на категории, эквалатеральный (равносторонний) тип топографии волосяного покрова, а также некоторые особенности в строении и форме волос. Для кроющих (направляющих и остевых) волос куторы характерна более расширенная и уплощенная, чем у наземных форм, концевая гранна. Возвышаясь над пухом и прикрывая его сверху, широкие и плоские гранны остевых волос образуют черепицеобразный покров и благодаря поверхностному натяжению воды сохраняют термоизолирующую воздушную прослойку в подпуши (Гудкова, Аксенова, 1951; Соколов, 1973). Этой же цели служит большая густота пуховых волос, волнистость их стержней и относительно сильное по сравнению с остевыми и направляющими волосами развитие сердцевинного слоя (72–73 % против 49–50 % у кроющих). Характерное для остевых волос куторы более плотное, чем у бурозубок, прилегание кутикулярных чешуек к стержню волоса и друг к другу обеспечивает более гладкую поверхность волос, их лучшую сомкнутость и более эффективную защиту подпуши от намокания.
Заключение
Итак, в Карелии в условиях северной периферии видовых ареалов волосяной покров изученных видов мелких мышевидных грызунов и насекомоядных млекопитающих, обитающих здесь в условиях постоянных термических дефицитов, как и свойственное им общее несовершенство терморегуляции, выработало у них целый комплекс экологических и морфофизиологических приспособлений, направленных на всемерное сохранение и усовершенствование оптимального энергетического баланса организма со средой при низких и крайне неустойчивых температурах севера. Особенно значимы соответствующие адаптивные особенности покровов, обнаруженные у них на всех показателях физической терморегуляции, в т.ч. и таких важнейших ее механизмах, как строение и теплозащитные свойства волосяного покрова и тонкая микроструктура волос и кожи, с их приспособительной сезонно-возрастной и популяционной изменчивостью, тесно связанной с обитанием в весьма специфических условиях приземного слоя воздуха. Проведенное нами исследование нельзя, конечно, считать исчерпывающим во всех аспектах затронутой проблемы, однако, как мы надеемся, оно позволит значительно активизировать продолжение работ в этой весьма перспективной области наук.
Библиография
Башкиров И. С., Жарков И. В. Биология и промысел крота в Татарии // Ученые записки Казанского государственного университета. 1934. Т. 94, вып. 3. С. 18–31.
Большаков В. Н. Пути приспособления мелких млекопитающих к горным условиям . М.: Наука, 1972. 199 с.
Большаков В. Н., Некрасов Е. С. О теплозащитных свойствах наружных покровов горных грызунов // Научные доклады высшей школы. Сер.: Биол. наук. 1968. № 10. С. 23–27.
Гудкова-Аксенова Н. С. Среда обитания и ее влияние на организацию некоторых водных насекомоядных и грызунов // Ученые записки Горьковского государственного университета. 1951. Вып. 19. С. 135–174.
Давыдов А. Ф. Гомойотермные организмы бореальной климатической зоны // Экологическая физиология животных. Л.: Наука, 1982. Ч. 3. С. 110–130.
Ивантер Э. В. Строение, теплозащитные свойства и линька волосяного покрова обыкновенной бурозубки (Sorex araneus) // Зоологический журнал. 2021. Т. 100, № 4. С. 419–433.
Ивантер Э. В., Ивантер Т. В., Туманов И. Л. Адаптивные особенности мелких млекопитающих. Эколого-морфологические и физиологические аспекты . Л.: Наука, 1985. 318 с.
Ивантер Э. В., Левина Р. В. Морфологическая характеристика и теплозащитные свойства шерстного покрова насекомоядных млекопитающих // Вид и его продуктивность в ареале: Материалы III Всесоюзного совещания. Вильнюс, 1980. С. 67–68.
Ивантер Э. В., Левина Р. В. Структура и теплоизоляционные свойства волосяного покрова средней бурозубки (Sorex caecutience Laxm.) // Экология наземных позвоночных Северо-Запада СССР. Петрозаводск, 1981. С. 110–119.
Ивантер Э. В., Моисеева Е. А. Строение, теплозащитные свойства и линька волосяного покрова европейского крота (Talpa europaea) // Труды КарНЦ РАН. 2022. № 3. С. 844–853.
Калабухов Н. И. Методика экспериментальных исследований по экологии наземных позвоночных . М., 1951. 177 с.
Калабухов Н. И. Предпочитаемая температура млекопитающих и ее связь с другими особенностями их терморегуляции // Грызуны и борьба с ними. Саратов, 1957. Т. 5.
Калабухов Н. И. Периодические (сезонные и годичные) изменения в организме грызунов, их причины и последствия . Л.: Наука, 1969. 248 с.
Кузнецов Б. А. Товароведение пушно-мехового сырья СССР . М.: Заготиздат, 1932. 464 с.
Кузнецов Б. А. Основы товароведения пушно-мехового сырья . М.: Заготиздат, 1952. 508 с.
Марвин М. Я. Строение волосяного покрова полевок рода Microtus Среднего Урала // Ученые записки Уральского государственного университета. Сер.: биол. Свердловск, 1966а. Вып. 3. С. 52–57.
Марвин М. Я. Влияние внешней среды на строение меха млекопитающих // Ученые записки Уральского государственного университета. Сер.: биол. Свердловск, 1966б.
Марвин М. Я. Строение волосяного покрова грызунов Урала // Научные труды Свердловского государственного педагогического института. 1974. Сб. 235. С. 11–22.
Назарова И. В. Эколого-морфологический очерк серых полевок Волжско-Камского края // Известия Казанского филиала АН СССР. Сер.: биол. 1958. № 6. С. 27–41.
Павлова Е. А. Возрастная и сезонная изменчивость меха ондатры // Труды ВНИО. М., 1955. Вып. 15. С. 59–93.
Прасолова Л. А., Киселев Ю. А., Выставной А. И., Уманцева Н. Д. Межпородные различия в термоизолирующих свойствах шерстного покрова крупного рогатого скота // Видовые и природно-климатические адаптации организма животных. Новосибирск, 1967. С. 273–276.
Скурат Л. Н. Строение волосяного покрова грызунов семейства Беличьих // Грызуны. М., 1980. С. 111–113.
Слоним А. Д. Животная теплота и ее регуляция в организме млекопитающих . М.; Л.: Госиздат, 1952. 327 с.
Слоним А. Д. Основы общей экологической физиологии млекопитающих . М.; Л.: Биомедгиз, 1961. 432 с.
Слоним А. Д. О физиологических механизмах адаптаций животных и человека . Л., 1964. 64 с.
Слоним А. Д. Экологическая физиология животных . М.: Наука, 1971. 448 с.
Смирнов П. К. Эколого-физиологическое исследование некоторых видов грызунов . Л., 1968. 135 с.
Содолевская Г. Я. О строении чешуйчатого слоя волос у близких видов мышей // Доклады и сообщения Ужгородского университета. 1957. № 1. С. 35–37.
Соколов В. Е. Кожный покров млекопитающих . М.: Наука, 1973. 487 с.
Соколов В. Е., Скурат Л. Н., Никифорова Ж. М. Морфологическая характеристика волос грызунов семейства летяг (Pteromyidae) и беличьих (Sciuridae) // Зоологический журнал. 1981. Т. 60, вып. 7. С. 1058–1069.
Соколов В. Е., Скурат Л. Н., Степанова Л. В. и др. Руководство по изучению кожного покрова млекопитающих . М.: Наука, 1986. 279 с.
Соколов В. Е., Чернова О. Ф. Новые данные об архитектонике волос землероек и кротов (Insectivora: Soricidae, Talpidae) // Доклады Академии наук. 1998. Т. 360, № 5. С. 717–720.
Уманцева Н. Д. Адаптивные особенности терморегуляции полевок тундровой зоны : Автореф. дис. … канд. биол. наук. М., 1980. 23 с.
Фадеев Е. В. Строение меха нутрии // Труды ВНИО. М., 1955. Вып. 15. С. 93–104.
Хмелевская Н. В. Строение кутикулы волос грызунов, ее изменчивость и значение для систематики // Зоологический журнал. 1965. Т. 44, вып. 7. С. 1064–1074.
Церевитинов Б. Ф. Дифференцировка волосяного покрова пушных зверей // Труды ВНИО. М., 1951. Вып. 10. С. 6–17.
Церевитинов Б. Ф. Топографические особенности волосяного покрова пушных зверей // Труды ВНИИЖП. М., 1958. Вып. 17. С. 256–307.
Чернова О. Ф., Целикова Т. Н. Атлас волос млекопитающих . М.: Товарищество научных изданий КМК, 2004. 429 с.
Шварц С. С., Смирнов В. С., Добринский Л. Н. Метод морфо-физиологических индикаторов в экологии наземных позвоночных . Свердловск, 1968. 387 с.
Шилов И. А. О некоторых понятиях, связанных с изучением проблемы терморегуляции // Зоологический журнал. 1962. Т. 41, вып. 4. С. 491–497.
Шмидт-Ниельсен К. Физиология животных. Приспособление и среда: В 2 кн. . М.: Мир, 1982. 785 с.
Appelt H. Lichtmikroskopische untersuchungen an fledermaushaapen // Abh. Und Ber. Naturk. Mus “Mauritianuym”, Altenburg. 1979. Bd. 10, No 3. P. 279–295.
Aurin W. Ist die Unterschridung von Hasen- und Kaninchenhauren durch Untersuchungder Cuticula und der Marksulsanz moglich // Dtsch. Ges. Gerichtgl. Med. 1933. Bd. 20, No 4. P. 54-58.
Benedict A. F. Hair structure as a generic character in bats // Univ. Calif. Publ. Zoolk. 1957. Vol. 59. P. 285–548.
Bergersen B. Beitrage zur Kenntnis der Haut einiger Pinnipedien. Oslo, 1931. 146 p.
Bonsma J., Pretorins A. Influence of colour and cover adaptability of cattle // Farming in South Africa. 1943. Vol. 18. P. 101–120.
Borowski S. Sezonowe zmiany uwlosienia Soricidae // Ann. Univ. M. Curie Sklod. Sect. C. 1952. Vol. 7, No 2. S. 65–117.
Borowski S. Variations in coat and colour in representatives of the genera Sorex L. and Neomys Kaaup. // Acta theriol. 1973. Vol. 18, No 14. P. 247–279.
Brown T. M. The microscopy of mammalian hair for Antropologist // Proc. Amer. Philos. Soc. 1942. Vol. 3. P. 250–274.
Cole H. J. Taxonomic value of hair in Chiroptera // Philippine J. Sci. 1924. Vol. 24, No 1. P. 117–121.
Hutterer R. H., Hurter T. Adaptiuve Hair Structuren bei Wasserspitzmausen (Insectivora, Soricidae) // Z. Saqugetierk. 1981. Bd. 46. S. 1–11.
Kapischke I. J., Muhkle H.-J. Zur Morphologie der Waldspoitzmaus (Sorex araneus) // Staatl. Mus. Tierk. Dresden. 1988. Bd. 44, No 1. S. 71–74.
Keller A. Determination des mammiferes de la Suisse par leur pelage: Talpidae et Soricidae // Revue Suisse Zool. 1978. No 85, Fasc. 4. P. 758–761.
Keller A. Determination des mammiferes de la suisse par leur pelage. IV Cricetidae et Muridae // Rev. suisse zool. 1981. Vol. 88, No 2. P. 463–473.
Liihring K. Das Haarkleuid von Sciurus vulgaris L. und die Verteilung seiner Farivarianten in Deutachland // 8 Z. Morphol. Okol. Tiere. 1928. Bd. 11, H. 5. S. 668–762.
Sauer M. Die Bedeutung der Oberhautzeichung fur die Unterscheidung von Haaren vershidener Herkunft und eine Farbung zur Darstellung dieser Zeichnung // Disch. Z. Gerichil.-Med. Berlin, 1922. S. 98–107.
Scaren U. Spring moult and onset of the breeding season jn the common shrew (Sorex Araneus L.) in Central Finland // Acta theriol. 1973. Vol. 18, No 23. P. 443–458.
Smith H. The relationships of themedullae and cuticular scales of the hair shafis of the Soricidae // J. Morphol. 1933. Vol. 55, No 1. P. 137–149.
Vogel P., Köpchen B. Besondere Haarstructuren der Soricidae (Mammalia, Instctivora), und ihre taxonomishe Deutung // Zoomorphologia. 1978. Bd. 89. S. 47–56.
Williamson J. Determination of hairs by impressions // J. Mammals. 1951. Vol. 32, No 1. P. 80–84.

 © 2011 - 2026
© 2011 - 2026