



Введение
В течение последних десятилетий понимание того, какие условия содержания животных в неволе соответствуют их потребностям, сильно изменилось. Когда-то было достаточно просто демонстрировать животных публике, но сегодня животные экспонируются в условиях, воспроизводящих их природную среду, им предоставляется множество возможностей взаимодействия с разнообразными предметами и объектами, которые должны стимулировать их естественное поведение и умственную деятельность. В основу зоопарковской биологии (Zoo biology) был положен главный принцип – улучшить жизнь животного в неволе возможно, изменив его окружение. Реализация этой идеи легла в основу подхода обогащения среды (environmental enrichment), который был создан на базе практики по нормализации поведения животных в зоопарке. К настоящему времени в рамках обогащения среды решаются такие задачи, как повышение разнообразия естественных форм поведения и общей двигательной активности, снижение патологического поведения и увеличение площади использования доступного пространства. Выполнение вышеперечисленных задач направлено на повышение благополучия животных, изучение которого (в сельском хозяйстве, лаборатории и зоопарках) сформировало новую научную дисциплину – науку о благополучии (Animal welfare science). Результаты исследований благополучия оказывают существенное влияние на законодательство и практику содержания животных в неволе, а также формируют облик современного зоопарка. Задачами зоологических парков стали: создание надлежащих условий содержания, поддержание высокого благополучия животных и изучение механизмов поддержания благополучия в неволе (Подтуркин, 2013).
Агонистические факторы окружающей среды, которые влияют на физиологию и психологию животных, описываются как стрессоры. Голод, жажда, недоедание, болезни, паразиты, враждебные контакты и неблагоприятные погодные условия – это естественные стрессоры. Эти факторы могут быть ограничены в случае медведей, содержащихся в неволе под опекой человека (отметим, что вследствие этого большинство видов животных в зоопарках живет значительно дольше, чем в естественных условиях). Однако животные в неволе сталкиваются с другими стресс-факторами: недостаток активности, посетители, работники зоопарка, искусственные рационы, неестественность суточного режима на ежедневной основе. Боль и голод – это факторы физического стресса, тогда как страх, паника, печаль, сенсорный стресс из-за громких шумов, климат, давление, социальный стресс из-за избытка животных в вольере, болезней, отравлений, социальных структур – это психологические факторы стресса (Кнёпфер, 2015).
Медведи – группа хищных млекопитающих, которым свойственно проявлять в условиях неволи высокий уровень патологического поведения, что делает этих животных удобным объектом для исследования причин нежелательного поведения диких животных в зоопарках (Подтуркин и др., 2015).
Цель настоящей работы заключается в выявлении особенностей поведенческой активности бурых медведей (Ursus arctos) в условиях зоопарка.
Материалы
Материал для исследования собирался на базе экопарка «Зоолесье» (г. Петрозаводск) в июне – июле 2021 г. Также в статье использованы аналогичные данные двухлетней давности (2019 г.).
Объекты исследования – два бурых медведя:
1) взрослый самец по кличке Барни, возраст 5 лет. В экопарк «Зоолесье» прибыл в возрасте 1 года в 2017 г.
2) взрослая самка по кличке Топтыжка, возраст 2 года (в 2019 г. она была еще медвежонком).
Площадь вольера самца 40 м2, самки – около 50 м2. Вольеры и у самки, и у самца разделены на две части, с помощью запирающихся ворот медведей кормят и чистят вольер. Питание Барни и Топтыжки двухразовое: утром в 11 и вечером в 18 часов.
Методы
Этограмма – это перечень поведенческих патернов – двигательных актов и фиксируемых положений тела, свойственных данному животному (Попов, Ильченко, 2008). В процессе предварительного наблюдения за животными мы выявили поведенческие паттерны, наиболее свойственные данным медведям, и на основе этого составили этограммы, которые отражают всю последовательность поведенческих актов у подопытных медведей, что дает возможность досконально изучить их поведенческий репертуар (табл. 1). Как видно из табл. 1, медведям, живущим в экопарке «Зоолесье», свойственны как нормальные для данного вида животных формы поведения, так и несвойственные медведям в дикой природе формы активности – «просит корм стоя и дует в трубу», «ходит по кругу» (пэйсинг).
Таблица 1. Этограммы медведей (самца и самки)
| Формы поведения | Поведенческие паттерны | |
| Самец | Самка | |
| Нет двигательной активности | 1. Сидит-наблюдает; 2. Стоит-наблюдает (на четырех лапах); 3. Лежит |
1. Сидит-наблюдает; 2. Стоит-наблюдает (на четырех лапах); 3. Лежит |
| Нормальная двигательная активность | 4. Ходит туда-сюда; 5. Стоит на задних лапах; 6. Переходит из одной клетки в другую; 7. Чешется |
4. Ходит туда-сюда; 5. Стоит на задних лапах; 6. Переходит из одной клетки в другую; 7. Чешется |
| Двигательные стереотипы | 8. Ходит по кругу (пэйсинг) | 8. Ходит по кругу (пэйсинг) |
| Подкорм и другие контакты с посетителями | 9. Просит корм стоя (смотрит на посетителей и тянется к ним); 10. Дует в трубу |
9. Дует в трубу |
| Активность при обогащении среды шиной | 11. Держит шину; 12. Грызет шину; 13. Качает шину |
10. Держит шину; 11. Грызет шину; 12. Качает шину |
Далее мы провели эксперимент по наблюдению за поведением бурых медведей в условиях зоопарка с помощью метода временных срезов. Суть этого метода – в «точечных» или «мгновенных» описаниях состояния наблюдаемого объекта, производимых через равные промежутки времени (Попов, Ильченко, 2008).
Мы проводили измерения в течение 30 дней в вечернее и утреннее время до кормления животного, по 2 измерения в день (самец и самка). Каждое измерение – в течение 30 минут с промежутком через 1 минуту. Из них 10 дней измерения проводили в вечернее время, 10 дней – в утреннее, 10 дней – в вечернее время с обогащением «подвешенная на цепях шина». Всего проведено 60 сессий, по результатам измерений составлено 30 регистрационных таблиц поведенческих паттернов медведей (в каждой таблице – самец и самка). Пример представлен в табл. 2 (за 21 июня).
Таблица 2. Регистрационная таблица поведенческих паттернов самки (красный крестик) и самца (голубой крестик) бурого медведя. 21.06.2021, вечер
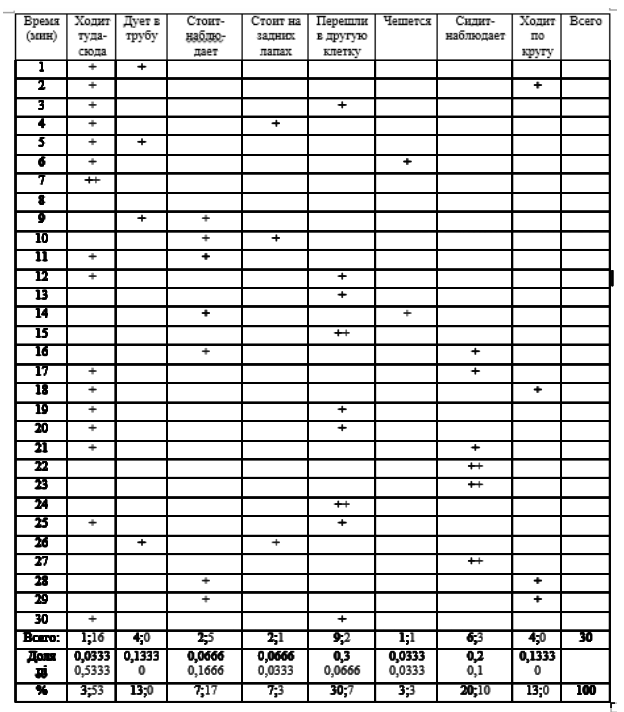
Полученные в работе результаты обработаны статистически с помощью U-критерия. Для оценки разнообразия поведения мы применили индекс разнообразия Шеннона. Этот индекс обычно используется для оценок видового разнообразия в экологических исследованиях; однако с его помощью можно оценить и разнообразие поведения. Индекс подсчитывают по формуле: H = -Σ pi * ln (pi), где pi – доля данной формы поведения в бюджете времени.
Результаты
На основе полученных данных для каждого из двух медведей мы определили бюджеты времени – системы показателей, характеризующие распределение затрат времени по видам его использования, и вывели по ним сравнительные характеристики. Для подсчета бюджета времени использовали доли регистраций соответствующей формы поведения в числе всех регистраций (срезов) (Попов, Ильченко, 2008). Результаты представлены на рис. 1, 2, 3, 4.
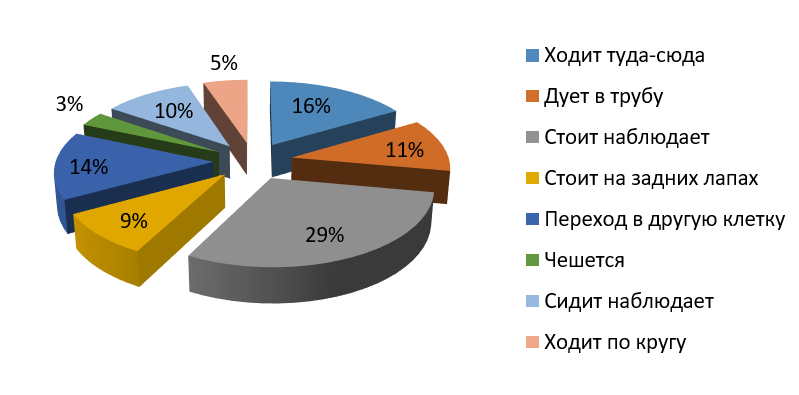
Рис. 1. Бюджет времени самца бурого медведя (вечер)
Fig. 1. Time budget of the male brown bear (evening)
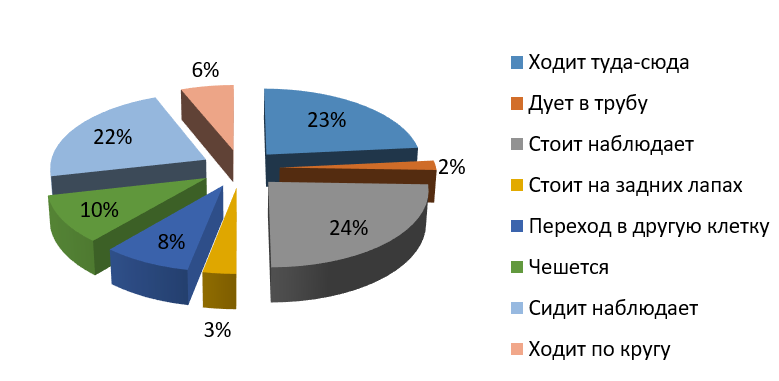
Рис. 2. Бюджет времени самца бурого медведя (утро)
Fig. 2. Time budget of the male brown bear (morning)
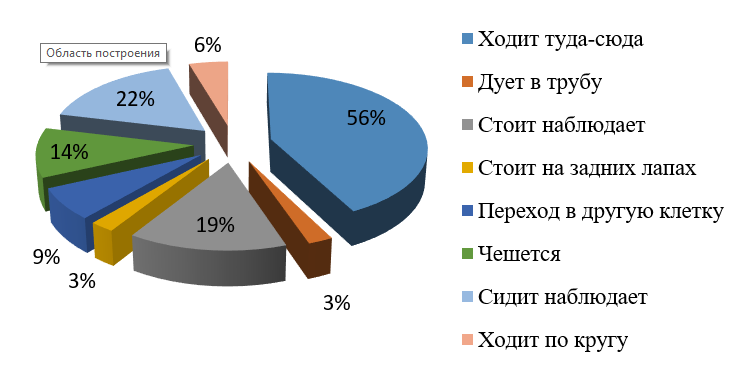
Рис. 3. Бюджет времени самки бурого медведя (вечер)
Fig. 3. Time budget of the female brown bear (evening)
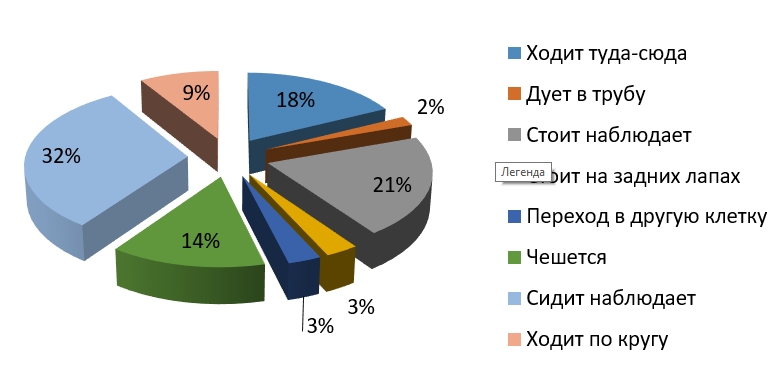
Рис. 4. Бюджет времени самки бурого медведя (утро)
Fig. 4. Time budget of the female brown bear (morning)
Как видно из рис. 1–4, у самца вечером наибольшее количество времени приходится на наблюдения и составляет примерно 29 %. Медведица вечером больше половины времени тратит на ходьбу в разные стороны клетки, 41 % – на наблюдения стоя или сидя. В основном и самец, и самка наблюдают за посетителями зоопарка, отвлекаются на посторонние звуки, такие как плач детей, пение птиц, шум машин. У самца 14 % приходится на попрошайничество (дует в трубу) и 3 % на чесание об решетку. У самки 3 % приходится на попрошайничество, 14 % на чесание (чесание может быть связано с меткой территории или зудом от укуса комаров).
Утром большую часть времени самец отдыхает и наблюдает – около 46 % (из них стоит-наблюдает 24 %, сидит-наблюдает 22 %), однако довольно много времени двигается – 23 %. Самка утром менее активна: стоит 21 %, сидит 32 % от общего бюджета времени. Скорее всего, это связано с тем, что ее клетка находится на солнечной стороне, тень не такая большая, как в клетке у самца. И самка, и самец по утрам тратят лишь по 2 % времени на попрошайничество (возможно, это связано с отсутствием посетителей зоопарка по утрам). На чесания приходится 14 % у самки и 10 % у самца.
Проведя анализ четырех диаграмм (см. рис. 1, 2, 3, 4), мы составили общие диаграммы сравнительной характеристики бюджета времени для самки и самца в утреннее (рис. 5) и вечернее время (рис. 6); для самки в утреннее и вечернее время (рис. 7) и для самца в утреннее и вечернее время (рис. 8). Результаты обработаны статистически с помощью U-критерия.
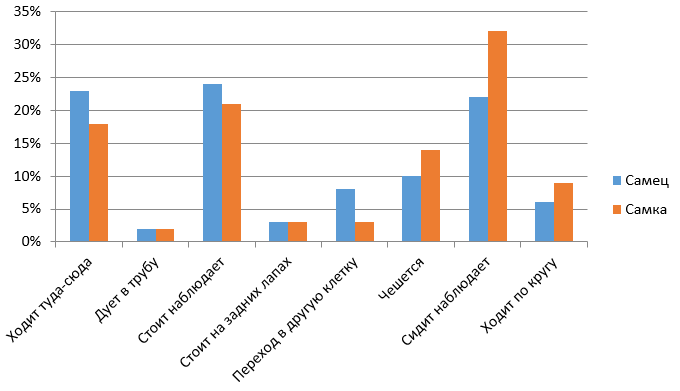
Рис. 5. Сравнительная характеристика бюджетов времени самки и самца в утреннее время
Fig. 5. Comparative characteristics of the time budgets of the female and male in the morning
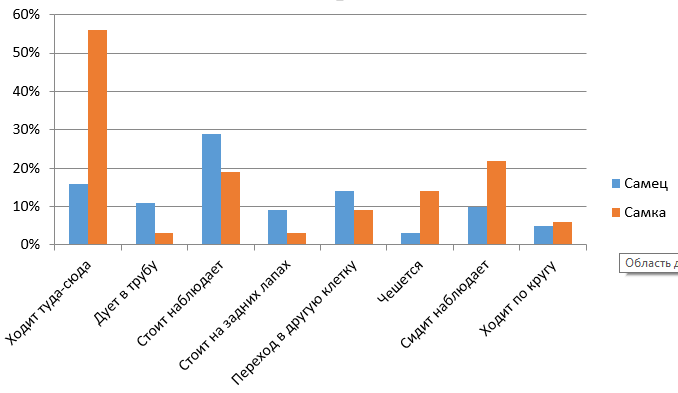
Рис. 6. Сравнительная характеристика бюджетов времени самки и самца в вечернее время
Fig. 6. Comparative characteristics of the time budgets of the female and male in the evening
Из рис. 5 и 6 видно, что нормальная активность самца бурого медведя, к которой следует относить ходьбу по сторонам клетки, чесание об решетку и наблюдения стоя либо сидя, составляет 79 % утром и 52 % вечером, т. е. самец более активен утром, чем вечером (различия недостоверны). У самки нормальная активность составляет 85 % утром и 11 % вечером. На нежелательную активность, к которой относим подуть в трубу, стояние на задних лапах и круговую ходьбу, в процентном соотношении большее количество времени тратит самец, примерно 30 %, а самка – около 20 %.
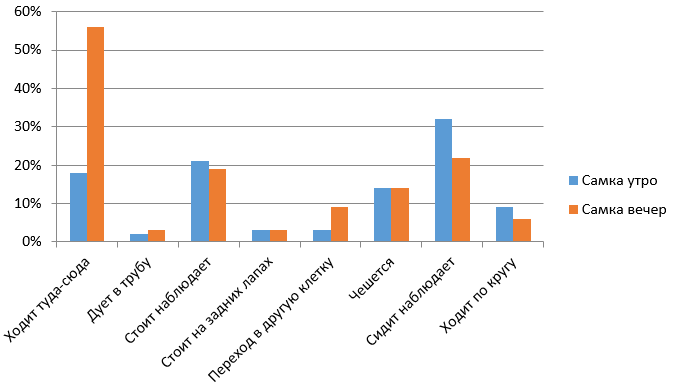
Рис. 7. Сравнительная характеристика бюджетов времени самки в утреннее и вечернее время
Fig. 7. Comparative characteristics of the time budgets of the female in the morning and in the evening
По рис. 7 видно, что в вечернее время самка активнее, чем утром. Преобладающим показателем является ходьба по сторонам.
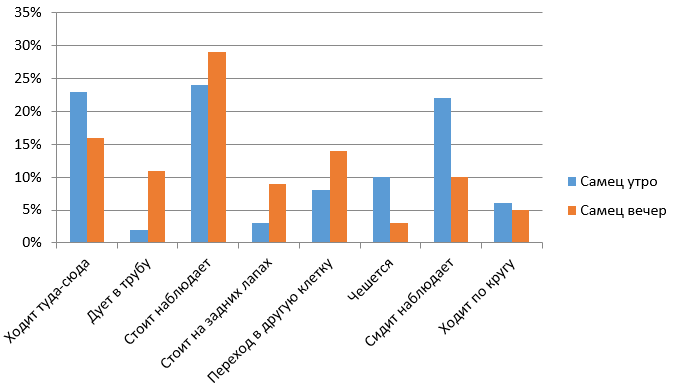
Рис. 8. Сравнительная характеристика бюджетов времени самца в утреннее и вечернее время
Fig. 8. Comparative characteristics of the time budgets of the male in the morning and in the evening
На рис. 8 видно, что вечером у самца преобладают наблюдение стоя, ходьба по сторонам и переход из клетки в клетку. А утром он больше всего ходит по сторонам, наблюдает и сидя, и стоя. Вечером у самца возрастает показатель попрошайничества (дует в трубу). Мы связываем это с приростом посетителей зоопарка.
Вторая серия опытов была направлена на исследование изменения поведенческой активности медведей при часто применяемом в зоопарках предметном обогащении среды в виде подвешенной на цепях шины (рис. 9, 10). Так как замеры активности медведей с шиной проводились в вечернее время, то для контроля мы взяли вечерние замеры активности медведей без шины. 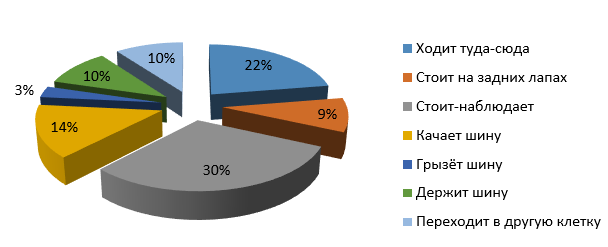
Рис. 9. Бюджет времени самки бурого медведя при обогащении среды подвешенной на цепях шиной
Fig. 9. Time budget of the female brown bear when enriching the environment with a tire suspended on chains
На рис. 9 видно, что большую часть бюджета времени самка Топтыжка наблюдала стоя (30 %), чуть меньше времени тратила на ходьбу по сторонам (22 %). Около 27 % от бюджета времени самка была увлечена шиной, а именно – качала шину (14 %), грызла шину (3 %) и держала шину (10 %). Около 10 % времени медведица потратила на переход из одной клетки в другую.
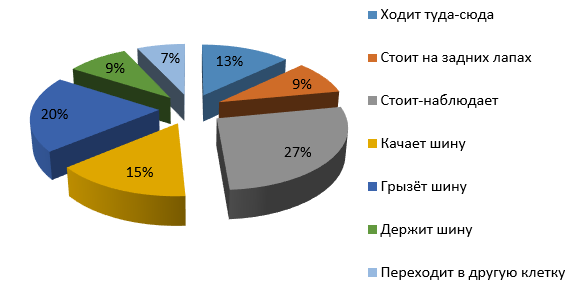
Рис. 10. Бюджет времени самца бурого медведя при обогащении среды подвешенной на цепях шиной
Fig. 10. The time budget of the male brown bear when enriching the environment with a tire suspended on chains
При обогащении среды шиной (см. рис. 10) почти треть времени (27 %) самец Барни тратил не на шину, а на наблюдения за окружающей обстановкой, примерно столько же времени (35 %) был отвлечен на шину: 20 % от бюджета времени грыз шину и 15 % качал шину. Остальное время он ходил по сторонам (13 %), переходил из клетки в клетку (7 %) и стоял на задних лапах (9 %).
С появлением шины и у самки, и у самца изменились виды активностей (рис. 11, 12). А именно такие активности, как дуть в трубу, чесаться, сидеть-наблюдать и ходить по кругу, перестали появляться. Возникли новые виды активностей – качать шину, грызть шину, держать шину.
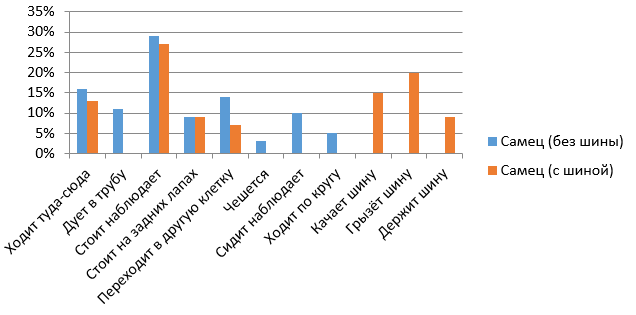
Рис. 11. Сравнительная характеристика бюджетов времени для самца в вечернее время (с шиной и без шины)
Fig. 11. Comparative characteristics of the time budgets of the male in the evening (with a tire and without it)
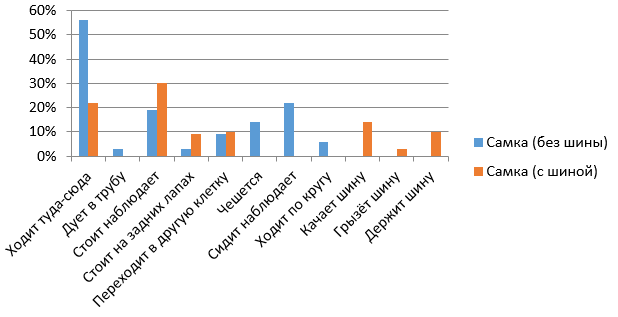
Рис. 12. Сравнительная характеристика бюджетов времени для самки в вечернее время (с шиной и без шины)
Fig. 12. Comparative characteristics of the time budgets for the female in the evening (with a tire and without it)
Для оценки разнообразия поведения мы применили индекс Шеннона. Как видно из рис. 13, 14, полученные в нашем исследовании коэффициенты разнообразия поведения медведей практически не отличаются – у самцов – самок и при сравнении показателей утро – вечер, что говорит о достаточно постоянном поведении на протяжении измеряемого периода. При этом коэффициенты вариации достаточно высокие, они лежат в промежутке от 2 до 3, что указывает на сравнительно большое разнообразие поведения медведей.
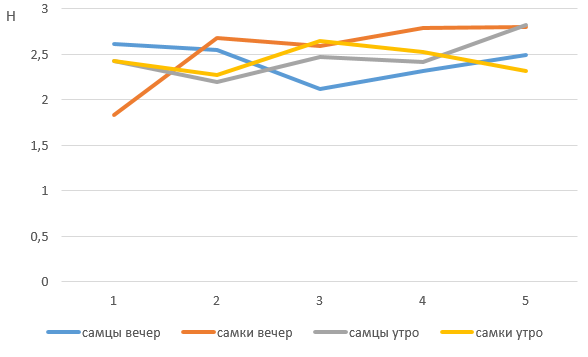
Рис. 13. Сравнение разнообразия поведения медведей с помощью коэффициента Шеннона (2022 г.)
Fig. 13. Comparison of the diversity of bear behavior using the Shannon's coefficient (2022)
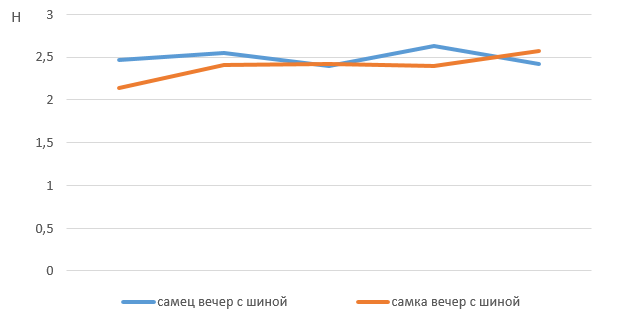
Рис. 14. Сравнение разнообразия поведения медведей с помощью коэффициента Шеннона при обогащении среды подвешенной на цепях шиной (2022 г.)
Fig. 14. Comparison of the diversity of bear behavior using the Shannon's coefficient when enriching the environment with a tire suspended on chains (2022)
Также мы сравнили результаты с ранее полученными коэффициентами вариации поведения данных медведей (двухлетней давности, лето 2019 г. (Moiseeva, 2021)). При сравнении диаграмм выяснилось, что ранее полученные коэффициенты вариации для Барни и медведицы были существенно ниже. Вероятно, можно говорить о повышении разнообразия поведения с возрастом (при сравнении данных самки с разницей в два года); разнообразие поведения Барни также повысилось.
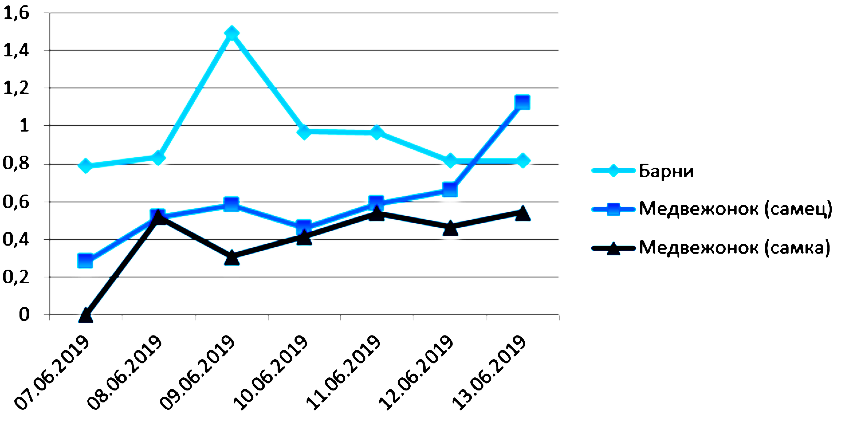
Рис. 15. Сравнение разнообразия поведения медведей с помощью коэффициента Шеннона (2019 г.)
Fig. 15. Comparison of the diversity of bear behavior using the Shannon's coefficient (2019)
Обсуждение
В бюджетах времени у обоих медведей при наблюдении без предметного обогащения преобладают две формы поведенческих активностей:
– ходьба по сторонам, являющаяся формой нежелательного поведения медведей и одновременно способом адаптации к условиям зоопарка, средством выплеснуть накопившуюся энергию;
– стояние на месте / наблюдение – отсутствие двигательной активности.
В утреннее время преобладают:
– ходьба по сторонам (больше у самца);
– наблюдения стоя и сидя.
В вечернее время также преобладают:
– ходьба по сторонам (больше у самки);
– наблюдения стоя и сидя.
Таким образом, утром активнее Барни, а вечером – Топтыжка. Преобладающим показателем активности самки является ходьба по сторонам. Мы предполагаем, что это связано с количеством посетителей в зоопарке (вечером их больше), а также с необходимостью выплеска энергии. Утром самка большее количество времени сидит и стоит-наблюдает – возможно, это связано с утренней жарой. Может быть, роль здесь играет также то, что вольер самки, в отличие от вольера самца, находится на солнечной стороне, и спастись от жары самка в период наших наблюдений могла лишь в одном из углов вольера. При сравнении показателей поведенческой активности утром и вечером достоверные отличия получены для самца по паттернам «дует в трубу» и «сидит-наблюдает», для самки – «стоит-наблюдает».
Другие формы двигательной активности, свойственные бурым медведям (вставание на задние лапы, попрошайничество, пэйсинг, чесание об решетку), представлены у исследуемых нами медведей из экопарка в меньшей степени (20–30 %).
В дикой природе бурые медведи встают на задние лапы в случаях возникновения опасности. В процессе наших наблюдений при появлении посторонних звуков, таких как сильный ветер или дождь, сигналы машин, оба медведя вставали на задние лапы, самка отводила на это 3 % своего времени, а самец – 9 %, но самец делал это также при виде людей для попрошайничества. То есть у самки вставание на задние лапы было связано только с реакцией на опасность и является нормальной двигательной активностью, а самец, который живет в зоопарке дольше самки, использует вставание на задние лапы для попрошайничества, что, вероятно, можно считать адаптацией к содержанию в неволе.
При детальном изучении попрошайничества у пары медведей-губачей в Московском зоопарке показано, что поведение выпрашивания не является исключительно способом получения еды, т. к. только половина подачек съедалась или обследовалась животными, а к остальным звери просто не приближались. Медведи научались манипулировать поведением посетителей, изменяя свое поведение в зависимости от числа людей перед вольером и их действий. Они развивали различные по эффективности поведенческие стратегии, позволяющие минимизировать усилия при выпрашивании (например, не начинали выпрашивать, если около вольера было меньше 5 человек) (Непринцева, Вощанова, 2009).
По наблюдениям Зубчаниновой, Попова (2010), поведение «выпрашивания» у изучаемого ими медведя-губача в Московском зоопарке было связано обратной зависимостью с проявлениями стереотипии: чем больше медведь выпрашивал подкормку у посетителей (и чем больше посетителей собиралось у вольера), тем реже он проявлял стереотипные формы поведения. По мнению этих авторов, существенная доля патологических форм поведения в бюджетах времени указывает на наличие стресс-факторов и перенапряжение нервной системы медведя. Такими факторами могут быть в первую очередь невозможность проявить естественное пищедобывательное поведение при общепринятых способах кормления, а также отсутствие положительных социальных контактов с партнерами. В многолетних наблюдениях этих авторов за медведем-губачом (2003–2008 гг.) доля патологических форм поведения в бюджетах времени существенно менялась от 4 до 55 %. Так, резкое снижение стереотипии в поведении медведя происходило после подселения самки в смежную с ним клетку после ее реконструкции (2005 г. – 42 %, 2006 г. – 38 %, 2007 г. – 14 %, 2008 г. – 4 %). Также в эти годы сотрудники зоопарка активно осуществляли кормовое и предметное обогащение, стимулировали игровое поведение медведя.
Отметим, что бывает достаточно сложно отличить нормальное поведение от патологического и попрошайничества, зачастую эти формы поведения могут иметь сходства. В частности, ходьба по сторонам с точки зрения психологии животных является показателем необходимости выплеска энергии, точно так же можно рассматривать ходьбу по кругу (пэйсинг), хотя пэйсинг обычно трактуют как типично патологическую форму поведения, свидетельствующую о том, что медведю не хватает двигательной активности. Постоянное или достаточно частое хождение по кругу свидетельствует о стремлении медведя оказаться на обширном пространстве с возможностью выплеска своей энергии. У исследуемых нами бурых медведей на пэйсинг отводится не так много времени (5–9 %).
В последние годы обогащение среды входит в практику содержания животных в зоопарках как обязательная составляющая (Подтуркин, Попов, 2012; Буянов и др., 2020). Однако, несмотря на широкое распространение внесения в вольеры различного рода «игрушек», имеется острый дефицит документированных исследований эффектов такого рода манипуляций (Подтуркин, Попов, 2010). Так, в Московском зоопарке взрослой медведице на протяжении полутора месяцев ежедневно предоставляли не менее трех способов обогащения среды из разных категорий: предметное, кормовое, когнитивное, запаховое, что привело к росту показателя разнообразия нормальной активности животного (Подтуркин, Сальникова, 2018). При этом сравнение рутинного (фон) и экспериментального обогащения среды показало большую эффективность последнего. Предоставление гепардам игрушки достоверно повышало их подвижность за счет снижения времени лежания, однако не имело долговременного сохранения (Подтуркин и др., 2008).
Эффективность обогащения среды принято оценивать по стандартным показателям: повышение разнообразия нормальной активности и снижение патологического поведения животного. В нашем эксперименте с появлением шины и у самки, и у самца изменились виды активностей. А именно такие активности, как дуть в трубу, чесаться, сидеть-наблюдать и ходить по кругу, перестали появляться, вместо них возникли новые виды активностей: качать шину, грызть шину, держать шину. У обоих медведей при появлении шины повысилась двигательная активность (самец 49 % и 66 %, самка – 86 % и 89%, но самец был более увлечен новым предметом, чем самка). Например, самец достоверно чаще самки грыз шину, а самка в период обогащения среды с помощью шины достоверно больше ходила туда-сюда.
Разнообразие поведения в процессе наших наблюдений и у самца, и у самки почти не менялось. Коэффициенты вариации достаточно высокие – лежат в промежутке от 2 до 3, что указывает на сравнительно большое разнообразие поведения медведей в данных условиях зоокомплекса. Мы склонны объяснить эти результаты достаточно благополучным окружением изучаемых нами медведей.
В заключение отметим, что нам представляется интересным проследить динамику поведения медведей по отношению к новым предметам (шина и другие предметы), т. к. очевидно, что достаточно высокий интерес к этому объекту был обусловлен новизной, и по прошествии времени произойдут изменения поведения.
Заключение
Таким образом, согласно нашим наблюдениям, двигательная активность самца бурого медведя не изменялась в течение дня, медведица же была значительно активнее вечером, чем утром. Вечером по сравнению с утром у самца достоверно возрастает попрошайничество, у самки не изменяется.
Проведенное нами обогащение среды с помощью подвешенной на цепи шины привело к изменению видов активностей медведей. Такие активности, как дуть в трубу, чесаться, сидеть-наблюдать и ходить по кругу, перестали появляться, вместо них возникли новые виды активностей, такие как качать шину, грызть шину, держать шину. При этом самец был более увлечен новым предметом, чем самка. Также наблюдалось повышение двигательной активности обоих медведей, что говорит об эффективности проведенного обогащения среды с помощью шины.
Коэффициенты разнообразия поведения медведей на протяжении измеряемого периода мало отличаются, что говорит о постоянном поведении обоих медведей. Коэффициенты вариации высокие, что указывает на достаточно большое разнообразие их поведения.
При сравнении коэффициентов разнообразия поведения медведей с полученными нами аналогичными показателями 2-летней давности для этих же медведей можно видеть, что разнообразие поведения и самца, и самки повысилось. Вероятно, это свидетельствует о повышении разнообразия поведения животных в зоопарке с возрастом при правильном содержании и уходе.
Библиография
Буянов И. Ю., Шушакова М. М., Третинникова Т. Н., Некипелова Е. О. Обогащение среды для семейства медвежьи в парке «Роев ручей» // Вестник КрасГАУ. 2020. № 3. С. 95–104.
Зубчанинова Е. В., Попов С. В. Мониторинг поведения медведя-губача (Melursus ursinus inornatus) в Московском зоопарке // Научные исследования в зоологических парках: Сборник научных статей. Вып. 26. М.: Московский зоопарк, 2010. С. 52–70.
Кнёпфер Ш. Рекомендации по содержанию бурого медведя (Ursus arctos) в неволе – обогащение и создание оптимальной среды . Австрия, FOUR PAWS, Центр спасения и реабилитации медведей, 2015. URL: https://kldzoo.ru/upload/medialibrary/493/obogashchenie-medvedey-ot-four-paws-perevod-na-russkiy-yazyk.pdf (дата обращения: 19.01.2023).
Непринцева Е. С., Вощанова И. П. Стратегии «попрошайничества» медведей-губачей в Московском зоопарке // Поведение и поведенческая экология млекопитающих: Материалы 2-й науч. конф. 9–12 ноября 2009 г., г. Черноголовка. М.: Тов-во научных изданий КМК, 2009. С. 88.
Подтуркин A. A., Попов С. В. Теоретические подходы к обогащению среды // Харьковский зоопарк. 2010. Вып. 5. С. 171–174.
Подтуркин A. A., Попов С. В. Влияние характеристик лабораторной среды обитания на исследовательское поведение полуденных песчанок (Meriones meridianus) // Зоологический журнал. 2012. Т. 91, № 2. С. 202–207.
Подтуркин А. А. Оптимизация среды обитания как способ повышения благополучия млекопитающих в условиях зоопарка : Автореф. дис. ... канд. биол. наук. М., 2013. 26 с. URL: https://new-disser.ru/_avtoreferats/01006733842.pdf (дата обращения: 29.01.2023).
Подтуркин А. А., Вощанова И. П., Непринцева Е. С. Выпрашивать нельзя, расхаживать – способы адаптации медведей к условиям неволи // От истоков к современности: 130 лет организации психологического общества при Московском университете: Юбилейная конференция. М., 2015. Вып. 5. С. 430–432.
Подтуркин А. А., Попов С. В., Непринцева Е. С. Опыт обогащения среды двух гепардов (Acinonyx jubatus) в Московском зоопарке // Научные исследования в зоологических парках. 2008. Вып. 24. С. 45–52.
Подтуркин А. А., Сальникова Т. Ю. Оценка эффективности обогащения среды самки бурого медведя (Ursus arctos beringianus) в Московском зоопарке // Научные исследования в зоологических парках. 2018. Вып. 33. С. 32–43.
Попов С. В., Ильченко О. Г. Методические рекомендации по этологическим наблюдениям за млекопитающими в неволе // Руководство по научным исследованиям в зоопарках. М.: Евроазиатская региональная ассоциация зоопарков и аквариумов: Московский зоопарк, 2008. С. 3–66.
Moiseeva, Т. А. Behavioral activity of Ursus arctos brown bear in zoo conditions // IOP Conference Series: Earth and Environmental Science. 2021. Vol. 677. P. 052068. URL: https://iopscience.iop.org/article/10.1088/1755-1315/677/5/052068/pdf.
Благодарности
Выражаем благодарность научному сотруднику Московского зоопарка, кандидату биологических наук Подтуркину Алексею Александровичу за ценные консультации, а также руководству и сотрудникам экопарка «Зоолесье» (г. Петрозаводск) за возможность проведения исследований.

 © 2011 - 2026
© 2011 - 2026