



Введение
Обширный ареал обыкновенной гадюки позволяет предположить наличие изменчивости многих адаптивных черт вида. Одна из особенностей, обеспечивающая проникновение вида далеко на север, – эффективная терморегуляция, которая позволяет змее поддерживать высокую температуру тела днем в условиях инсоляции. Для этого гадюка меняет позу, субстрат, место для баскинга. Такое поведение включается, когда текущая температура тела превышает некое критическое значение, играющее роль параметра терморегуляции. Главный вопрос нашего сообщения – меняются ли параметры терморегуляции вида с продвижением на север. Рассматривая вопрос об изменчивости термобиологических характеристик рептилий, В. А. Черлин (Черлин, 2012) высказал положение о стабильности ключевых видовых параметров терморегуляции в пределах всего ареала. К сожалению, эти положения не были доказаны статистически. Разрешение этого вопроса становится возможным в связи с изменением метода измерения температуры тела животного – с помощью вживленных температурных датчиков. Он дает возможность существенно увеличить объем выборок и подойти к статистической оценке параметров терморегуляции. Очевидно, для контрастного сравнения нужно было взять территории, на которых теплообеспеченность существенно отличается. Для исследуемых районов Карелии сумма активных температур оценивается в пределах 1000–1500 °C, а в Пермском крае – 1500–2000 °C (Уланова, 1975). Кроме этого, интересно было выяснить, отличаются ли параметры терморегуляции у особей с разной окраской и массой, а также по сезонам.
Цель нашей работы состоит в численной оценке и сравнении корректных термофизиологических параметров терморегуляции у разных групп обыкновенной гадюки.
Материалы
Работы выполнялись в 2014 г. в Пермском крае в окрестностях деревни Киселево (N57°7'23", E57°21'15") и в 2017–2019 гг. в Карелии на биологическом стационаре в д. Гомсельга (N62°3'19", E33°57'45") в конце весны и в первой половине лета – с 10 мая по 20 июля.
В Пермском крае регистрировалась температура тела у двух самок гадюки светлой и черной окраски (масса 100–110 г). Температуру тела фиксировали с помощью логгера типа ds1921 (Lourdais et al., 2013; Herczeg et al., 2007). Охлажденным до 5 °С животным посередине туловища над ребрами со стороны спины делали поперечный надрез кожи шириной около 1 см, под кожей формировали лакуну глубиной 1 см по направлению к голове, помещали в нее логгер и зашивали одним-двумя стежками. Гадюк выпускали в естественные условия; в конце лета их отыскивали, изымали логгер, гадюк выпускали, данные считывали. Запись данных велась летом 2014 г.: в мае – июне у светлой особи и в июне – июле у черной. Период считывания значений температуры тела был задан для светлой особи в 70 минут, для темной – 30 минут. С помощью логгеров синхронно фиксировалась температура воздуха в траве. База данных имеет объем около 3000 записей, из них 298 значений температуры тела выше 33 °С.
В Карелии на биостационаре в д. Гомсельга был построен вольер размером 8 × 8 м, представляющий собой полиэтиленовый забор высотой 1 м, нижняя часть которого вкопана в почву на 10 см. В центре вольера разместили груду камней диаметром 1.5 м и высотой 70 см – типичное укрытие гадюк в этой местности. Змей для опытов отловили на о. Кижи (на той же широте в 100 км на восток по прямой). Логгеры типа ds1921 вшивали под кожу спины посередине туловища. Имплантация под кожу была выбрана как менее травмирующая по сравнению с внутрибрюшинной, а также из-за опасения, что иначе змеи не смогут питаться. Опыт показывает, что ранка обычно полностью заживает через 2 недели. Периодически змей осматривали, но явные воспалительные процессы с выделениями зафиксированы не были. С логгерами на спине гадюки питались в природе, некоторые – и в вольере. Метод имплантации и тип датчика у всех змей из Пермского края и Карелии были идентичны, что позволяет выполнять сравнения их показаний. Температуру тела регистрировали в 2017 г. в мае и июне у 11 гадюк, в 2018 г. в июне у 6 гадюк, в 2019 г. в июне – июле у 5 гадюк с разной окраской (всего 23 особи массой 100–120 г). После вживления логгеров змей выпускали в вольер на 8–25 дней, затем логгеры изымали, считывали данные, змей выпускали в природу. Температуру тела у трех особей (2019 г.) регистрировали через одну минуту, у остальных – через 2 минуты; общая продолжительность наблюдений в отдельный месяц у разных змей составила от 8 до 25 дней. Параллельно с помощью логгеров измерялась температура воздуха в траве. В результате получена база данных с числом записей для каждой особи от 3000 до 15000. Число значений температуры тела выше 33 °С у разных змей составляло от 140 до 3000. Над вольером была установлена видеокамера, фиксирующая перемещения части меченых змей в светлое время суток.
Целью работы было оценить не «термоэкологические» усредненные характеристики температуры тела, зависящие от условий среды, но «термофизиологические» показатели, характеризующие терморегуляторные способности вида (Черлин, 2012). По этой причине из всего диапазона температур рассматривались только высокие значения температуры, близкие к максимальным, которые гадюки совершенно явно избегают.
Методы
Из числа разработанных нами параметров терморегуляции (Коросов, Ганюшина, 2020) мы использовали два – «средняя температура баскинга» и «типичная максимальная температура». Они имеют ясный физиологический смысл, хорошо статистически обоснованны и позволяют выполнять сравнение с использованием статистических критериев. Каждая змея получила оценки обоих показателей.
Средняя температура баскинга (To) рассчитывается как среднее арифметическое из всех температур тела гадюки, которые зафиксированы за все периоды времени, когда температура окружающей среды (то есть температура воздуха в траве) превышала 23 °С (рис. 1). В расчет принимали только те значения температуры тела, когда змеи пребывали на поверхности субстрата и были зафиксированы камерами слежения. Температура в траве играет роль всего лишь индикатора уровня солнечной радиации, при 23 °С он достаточно высок и вынуждает змею осуществлять терморегуляторное поведение. Как показывает наблюдение за змеями в природе и расшифровка видеозаписей слежения, в это время гадюка осуществляет активное терморегуляторное поведение, направленное на поддержание высокой температуры тела, но не превышающей некоторого порога. Варьирование температуры около этого уровня можно рассматривать как в каком-то смысле «оптимальную» для гадюки. Оценки To получены только для тех карельских змей, за которыми велись видеонаблюдения. Выборки значений по каждой гадюке для Карелии и Пермского края имеют объемы от 68 до 218 вариант. Они позволяют рассчитать среднюю величину, ее ошибку и построить доверительный интервал. Для оценки ошибки средней показателей средней оптимальной температуры применяли обычную формулу: mTo = STo / (n ^ 0.5) – отношение стандартного отклонения (STo) к корню из объема выборки.
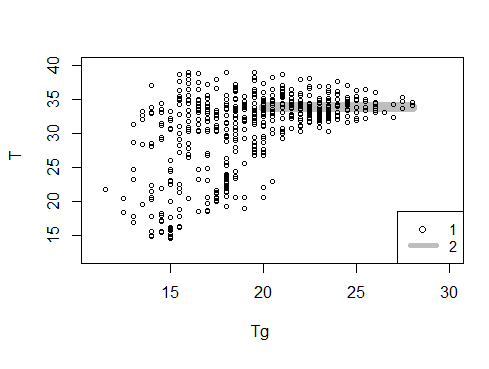
Рис. 1. Соотношение между температурой тела гадюки № 15 (T, °С) и температурой воздуха в траве (Tg, °С) за все время наблюдений (1 – значения температуры тела во время пребывания змеи на поверхности, 2 – границы доверительного интервала MTo ± 2mTo)
Fig. 1. The ratio between the body temperature of the adder No. 15 (T, ° С) and the air temperature in the grass (Tg, ° С) for the entire observation period (1 – body temperature values during the snake's stay on the surface, 2 – the boundaries of the confidence interval MTo ± 2mTo)
Типичная максимальная температура (Tmt) рассчитывается как некий предел в правой части полигона распределения значений температуры тела гадюки. На температурной шкале это такая точка, правее которой температура тела гадюки обычно не встречается.
Показатели максимальной температуры привлекательны тем, что интуитивно понятны, поскольку они характеризуют физиологическую верхнюю границу толерантного диапазона. К сожалению, единичные замеры максимальной температуры тела отдельных животных в природе несут на себе сильный отпечаток случайности, не обладают устойчивостью и их статистическое сравнение невозможно. Гораздо лучше статистически обоснованны квантильные границы, отсекающие, например, 95 % от выборки. Однако для пойкилотермных животных распределение температуры тела абсолютно не соответствует нормальному, поскольку вся левая часть состоит из температур, никак не характеризующих физиологические потребности животных, но является отражением температурных условий окружающей среды. По такому распределению нельзя построить двухсторонние квантильные границы. С другой стороны, нам важна только правая сторона распределения температур, значит, необходимо сконструировать показатель, соответствующий какой-либо квантиле или ее аналогу.
Для оценки типичной максимальной температуры использовались два метода – расчет с помощью квантилей и линейной регрессии. В качестве исходных данных для каждой гадюки послужило распределение температуры тела в течение всех дней наблюдений. От этого распределения отбрасывали часть, находящуюся левее модального класса (имеющего максимальную частоту), который всегда приходился на 34 °С. Общий объем значений температуры выше 33 °С для разных змей варьировал от 140 до 2900 вариант, поскольку продолжительность наблюдений и частота считывания температуры тела в разных опытах различались. Оставшуюся правую часть распределения разбивали на несколько классовых интервала (от 10 до 20 в зависимости от объема выборки) и анализировали. Всего получены показатели для 23 гадюк из Карелии и 2 гадюк из Пермского края. Поскольку объемы выборок для пермских гадюк были невелики, их данные объединили и получили общую выборку из 298 значений. Предварительный анализ распределений для этих змей показал их большое сходство и возможность объединения данных.
Метод определения квантильных границ состоял в следующем. Исходными данными каждой змеи служили два ряда значений температуры тела (T и интервалом около 0.4 °С) и частоты их встречаемости (f) (рис. 2).
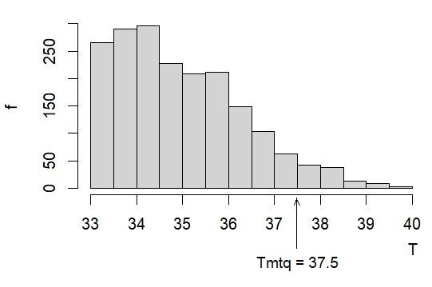
Рис. 2. Типичный вариант распределения значений температуры тела гадюки (№ 18) и положение границы Tmtq
Fig. 2. Typical variant of the distribution of the body temperature values of the adder (No. 18) and the position of the Tmtq border
Общий объем выборки для каждой змеи составлял n. Поскольку от этой выборки предстояло найти границу, отделяющую 95 % особей от 5 % особей, находили число особей, составляющих искомые 5 %, – n5 = n * 0.05, для выборки из 300 значений получаем n5 = 300 * 0.05 = 15. Далее ранжируем все значения по убыванию и из полученного ряда берем значение на рассчитанном пятнадцатом месте – это и будет искомая квантильная граница, максимальное типичное значение Tmtq, соответствующее доверительной вероятности 90 %.
Метод определения границ с помощью линейной регрессии состоял также в использовании правого ската распределения температур – от модального значения (34 °С) до максимального (рис. 3).

Рис. 3. Частотное распределение значений температуры тела гадюки (T, °С) и график зависимости температуры тела от частоты ее встречаемости (1 – частоты значений температуры, 2 – линия регрессии)
Fig. 3. Frequency distribution of viper's body temperature values (T, ° С) and the graph of the dependence of body temperature on its frequency (1 – frequency of temperature values, 2 – regression line)
Для каждой змеи вдоль по скату распределения уравнение регрессионной зависимости между значениями температуры тела (T) и частотой их встречаемости (f):
T = a * f + b.
По уравнению отыскивали то значение температуры, при котором частота встречаемости должна быть равна нулю T = b при f = 0. Это значение брали как максимальную типичную регрессионную температуру (Tmtr). Обычно за пределами этой границы с правой стороны остается небольшая группа особенно высоких значений температуры («хвост»), однако по сравнению с основной массой доля этих значений обычно незначительна и существенно не влияет на положение точки пересечения линии регрессии с осью абсцисс.
Для оценки статистической ошибки типичной максимальной температуры (при использовании обоих методов) использовали ресамплинг (Шитиков, Розенберг, 2013). Процедура состояла в том, что из исходной выборки значений температуры многократно (по 100 раз) извлекали некоторую часть данных ограниченного объема (вторичная выборка), для которой выполняли процедуру расчета значения типичной максимальной температуры. Каждую новую вторичную выборку составляли из случайно отобранных значений исходной выборки. По этой совокупности из 100 оценок типичной максимальной температуры для данной змеи рассчитывали, во-первых, среднее значение максимальной типичной температуры (MT), во-вторых, стандартное отклонение (mT), которое имеет смысл ошибки средней.
Для оценки значимости различий между параметрами для разных гадюк мы апробировали разные методы, но решили остановиться на самом простом и иллюстративном. Сравнение параметров терморегуляции проводили, сопоставляя доверительные интервалы.
Доверительные интервалы строились вокруг значения Mtt, полученного при объеме вторичной выборки n = 500 для уровня значимости p = 0.05 по формуле MT ± 2 * mt. Как известно, перекрывание доверительных интервалов более чем на 1/4 означает отсутствие значимых отличий между сравниваемыми средними, а хиатус – их наличие (Cumming, Finch, 2005). Зависимость от факторов была изучена с помощью GLM-регрессии (Мастицкий, Шитиков, 2015). Все расчеты выполнялись в среде программы R.
Результаты
Летним утром гадюки выходят из ночных укрытий (в 6–8 часов), только когда солнце начинает припекать; они принимают специфические позы и формы тела и быстро увеличивают температуру своего тела до уровня 32–35 °С (Коросов, 2008). В течение нескольких следующих часов температура субстрата постоянно растет, но температура тела гадюки остается относительно стабильной и колеблется около значения 34 °С (рис. 4). Для описания этого результата терморегуляторного поведения, направленного на поддержание высокой температуры тела, но не превышающей некоторого порога, и предложены наши показатели.
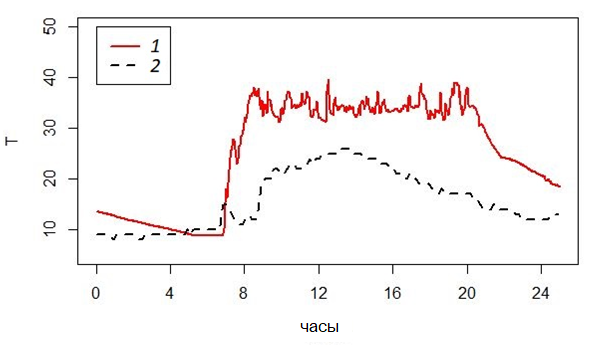
Рис. 4. Ход температуры (°С) тела гадюки (1) и воздуха в траве (2) в течение суток 29.06.2019
Fig. 4. Temperature variation (°С) of the adder's body (1) and air in the grass (2) during the day 29.06.2019
Первый вопрос, на который предстояло ответить, состоит в том, отличаются ли параметры терморегуляции у гадюк, отловленных в разных частях ареала. С этой целью на диаграммах были отображены доверительные интервалы, построенные вокруг средних значений оптимальной и типичной максимальной температуры для всех наблюдавшихся гадюк (рис. 5, 6).
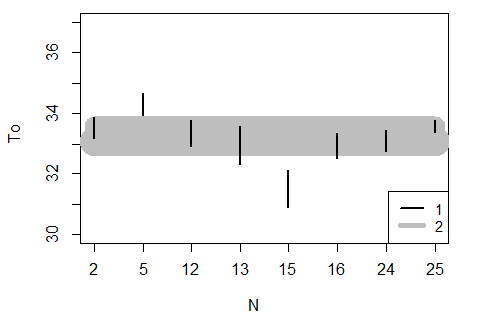
Рис. 5. Доверительные интервалы некоторых значений средней температуры баскинга гадюк (N – номер змеи, 1 – доверительные интервалы для змей из Карелии (2–16) и Пермского края (24, 25), 2 – доверительный интервал для гадюк из Пермского края)
Fig. 5. Confidence intervals for some values of the average basking temperature of adders (N – snake number, 1 – confidence intervals for snakes from Karelia (2–16) and Perm region (24, 25), 2 – confidence interval for adders from Perm region)
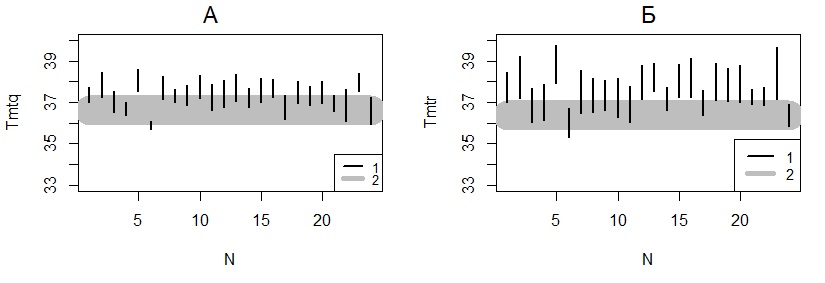
Рис. 6. Доверительные интервалы для оценок типичной максимальной температуры, оцененной методом регрессии (А, Tmtr) и методом квантилей (Б, Tmtq) (N – номер змеи, 1 – доверительные интервалы для змей из Карелии (1–23) и Пермского края (24) (p = 0.05, mT = 0.3 °С), 2 – доверительный интервал для гадюк из Пермского края)
Fig. 6. Confidence intervals for estimates of the typical maximum temperature evaluated by the regression method (A, Tmtr) and the quantile method (B, Tmtq) (N - snake number, 1 - confidence intervals for snakes from Karelia (1–23) and the Perm region (24) (p = 0.05, mT = 0.3 °С), 2 – confidence interval for adders from the Perm region)
Значения средней оптимальной температуры (To) для гадюк из Карелии и Перми укладываются в диапазоне 3 °С – от 31.5 до 34.3 °С, с общей средней 33.2 °С (таблица).
Значения средней оптимальной температуры тела и ее изменчивости для некоторых особей обыкновенной гадюки
| № | Topt | sd | m | n |
| 2 | 33.53 | 2.14 | 0.18 | 147 |
| 5 | 34.29 | 1.83 | 0.18 | 106 |
| 12 | 33.35 | 3.30 | 0.22 | 218 |
| 13 | 32.93 | 3.33 | 0.32 | 111 |
| 15 | 31.49 | 3.12 | 0.30 | 104 |
| 16 | 32.93 | 2.86 | 0.21 | 186 |
| 24 | 33.1 | 1.54 | 0.18 | 68 |
| 25 | 33.59 | 1.48 | 0.10 | 205 |
Стандартное отклонение (STo) для многих выборок составляет около 1.5 °С, ошибка средней не высокая, поэтому показатель «средняя температура баскинга» является достаточно устойчивым.
На фоне изменчивости значений особей из Карелии пермские гадюки занимают устойчивое срединное положение (см. рис. 5). Значимые отличия между значениями средней оптимальной температуры для групп гадюк из разных частей ареала не выявлены. Причины существенного варьирования средней оптимальной температуры у карельских змей пока остаются неизвестными.
Типичные максимальные температуры, оцененные у разных особей гадюки, варьируют в пределах 3 °С: Tmtr – от 35.9 до 39.1 °С (общая средняя составила 37.8 °С), Tmtq – от 35.9 до 38.1 °С (общая средняя – 37.3 °С).
Как можно видеть на диаграммах (см. рис. 6), диапазон значений типичной максимальной температуры для пермских гадюк (37 °С) находится внутри разброса значений карельских гадюк, хотя и вблизи от минимальных значений. Большинство змей из Карелии имеют достаточно высокие значения, но часть из них имеет низкие значения, такие же и даже меньше, чем у змей из Пермского края. Данные показывают, что значимые различия между изученными показателями терморегуляции гадюк из разных регионов не обнаруживаются. Мы не смогли опровергнуть гипотезу В. А. Черлина.
Вместе с тем очевидно, что у разных особей гадюки из Карелии эти параметры значимо отличаются – их доверительные интервалы в некоторых случаях вообще не перекрываются, что особенно ярко видно на примере особей 5 (Tmtq = 38.1 °С) и 6 (Tmtq = 35.9 °С). Однако причины этих различий остаются непонятными. Расчет коэффициентов корреляции между значениями максимальной типичной температуры и такими характеристиками 23 змей, как окраска (темная или светлая, значения 0 и 1), пол (самцы и самки, значения 0 и 1), масса тела (в диапазоне 50–140 г), ни в одном случае не дал значимых величин, которые даже не достигали уровня 0.2 (p > 0.1). Анализ влияния указанных факторов (регион, пол, окраска, масса, длина) с помощью GLM-регрессии также не дал значимых результатов (уровень значимости не опускался ниже p = 0.2). Таким образом, причины индивидуальных отличий параметров терморегуляции пока остаются непонятными.
Обсуждение
Лейтмотивом этой статьи, и вообще деятельности соавторов в последнее время, является поиск корректных термофизиологических показателей, соответствующих или тесно коррелирующих с физиологическими параметрами терморегуляции. В значительной степени наша работа является проверкой гипотезы, высказанной В. Черлиным (Черлин, 2012), о стабильности и неизменности физиологических параметров терморегуляции данного вида на всем своем ареале, которая не кажется бесспорной из-за методической недоработанности. Несмотря на строгие требования, которые этот автор предъявляет к себе и другим исследователям, его характеристики терморегуляторных способностей рептилий не снабжены статистическими ошибками, и выводы о сходстве параметров терморегуляции статистически не обоснованны. Есть у нас вопросы и к методам идентификации типов терморегуляторного поведения, количество которых представляется избыточным. В данном сообщении мы предприняли попытку сравнения параметров терморегуляции, во-первых, упростив схему вычленения терморегуляторных типов поведения и, во-вторых, обеспечив повторяемость наблюдений (рассчитав статистическую ошибку). Это позволило нам подойти к статистически обоснованным выводам по результатам сравнения параметров терморегуляции, имеющих, на наш взгляд, физиологический смысл.
Предложенная конструкция использованных параметров является результатом продолжительных исследований по термобиологии рептилий. К формированию показателя средней оптимальной температуры соавторы шли разными путями. С одной стороны, предлагался параметр «точка абсолютного оптимума», представляющий собой пересечение линий трендов температуры тела и температуры среды (Литвинов и др., 2013). С другой стороны, строилась модель суточной динамики температуры тела с единственным параметром – максимальная добровольная температура (Коросов, 2008). К сожалению, в обоих случаях анализ материала давал широкий диапазон возможных значений с плохим статистическим обоснованием. Выход из ситуации наметился, когда при выборе значений для расчетов оптимальной температуры начали ориентироваться на поведение особи, к чему давно призывал В. А. Черлин. И визуальное наблюдение в поле, и расшифровка непрерывных видеозаписей позволили сформулировать новые методические принципы, согласно которым из всех наблюдаемых значений температуры тела животного нужно брать в расчет только те, которые явно связаны с отчетливым терморегуляторным поведением (Коросов, 2015; Ганюшина и др., 2019). Значения температуры тела, непосредственно связанные с терморегуляторными реакциями рептилии, следует назвать термофизиологическими (Черлин, 2012), поскольку такие значения будут характеризовать внутренние физиологические пороговые критерии, запускающие терморегуляторные реакции.
Важно было сконструировать формальные критерии для отбора нужных значений. Один из таких критериев был получен при сопоставлении температуры тела гадюки и температуры окружающей среды (см. рис. 1). На рисунке хорошо видно, что при температуре воздуха в траве выше 23 °С значение температуры тела резко снижает свою изменчивость, и связано это именно с тем, что в условиях достаточного теплообеспечения животное может активно выбирать нужную температуру своего тела. Значения температуры воздуха в траве ниже 23 °С относятся к менее благоприятным условиям инсоляции, к переменной облачности, сильному ветру, темному времени суток и пр. Любые факторы, меняющие тепловой поток, тем самым увеличивают изменчивость температуры тела гадюки. Параметры, полученные в этих условиях, будут характеризовать не способность животного к терморегуляции, но феномен ограничения теплообеспеченности среды его обитания. Любые характеристики, прямо не связанные с терморегуляторными реакциями рептилий, по терминологии В. А. Черлина (Черлин, 2012), следует называть термоэкологическими характеристиками, широко варьирующими в зависимости от влияния средовых факторов; они нас не интересовали. При определении параметров терморегуляции выяснился ряд их особенностей. Оценки средней оптимальной температуры имеют довольно широкие доверительные интервалы. За редким исключением они перекрываются более чем на 1/3, т. е. в основном показатели средней оптимальной температуры для разных змей отличаются незначимо. Высокая изменчивость значений связана, видимо, с тем, что в выборку неизбежно попадают довольно низкие значения температуры тела змеи, которая недавно выбралась из укрытия на солнцепек и еще не успела нагреться. Конечно, такие моменты можно было бы отследить по кадрам видеорегистрации, однако специальный подбор данных для получения более высоких показателей, в общем-то, противоречит идее поиска объективных показателей. Это соображение заставляет признать, что корректная оценка средней оптимальной температуры, видимо, возможна только при большом объеме данных (более 200 значений), наших материалов для этого в некоторых случаях было недостаточно. Тем не менее показатель является устойчивым и, при уточнении его интерпретации, рекомендуется к использованию.
Показатели максимальной типичной температуры тела призваны охарактеризовать физиологическую верхнюю границу толерантного диапазона благоприятных для гадюки температур. Они имеют смысл термофизиологических показателей, тесно связанных с параметрами терморегуляции рептилий. Два метода расчета дали очень близкие результаты, хотя и не идентичные. Уровень корреляции между Tmtg и Tmtq равен 0.88 (p < 0.01). На наш взгляд, варьирование показателей связано с особенностями методов оценки. Квантильный метод статистически более обоснован и, судя по невысокой величине ошибки, более точен, нежели регрессионный метод. Основную погрешность вносит возможность случайного смещения модального класса, от которого зависит общий объем выборки, а значит, и доля в 5 %. Метод регрессионной оценки значения температуры с нулевой частотой дает достаточно однородные результаты, но только для гладкого распределения при большом объеме выборки (500 наблюдений температуры выше 33 °С). На наш взгляд, оба этих метода годятся для указанных целей и могут быть рекомендованы для использования.
Причины значимых отличий параметров терморегуляции у отдельных особей гадюки, которые пока не ясны и рассматриваются как случайные, могут быть связаны с тонкими физиологическими различиями, например, со стадией переваривания пищи, возрастом особи, скрытыми болезнями, стадией линьки и пр. Теперь задача состоит в том, чтобы выявить факторы, ответственные за варьирование индивидуальных параметров терморегуляции, а также расширить спектр таких параметров.
Заключение
- Оценки параметров терморегуляции у особей обыкновенной гадюки из разных частей ареала, удаленных на 1500 км, значимо не отличаются.
- Оценки максимальной типичной температуры у некоторых особей обыкновенной гадюки из Карелии отличаются значимо.
- Варьирование значений максимальной типичной температуры у разных особей обыкновенной гадюки из Карелии не связано с полом, окраской и массой тела.
- Показатель «типичная максимальная температура» легко определяется по наборам данных, имеет невысокую изменчивость для выборок объемом 300–500 вариант и обладает статистической ошибкой, что позволяет выполнять статистические сравнения параметров терморегуляции рептилий.
Библиография
Ганюшина Н. Д., Коросов А. В., Литвинов Н. А., Четанов Н. А. Наружная и внутренняя температуры тела обыкновенной гадюки // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2019. № 2. С. 17–27.
Коросов А. В. Новое слово в термобиологии рептилий? // Принципы экологии. 2015. № 1. С. 77–85. DOI: 10.15393/j1.art.2015.4162
Коросов А. В. Простая модель баскинга обыкновенной гадюки (Vipera berus) // Современная герпетология. 2008. Т. 8. Вып. 2. С. 118–136.
Коросов А. В., Ганюшина Н. Д. Методы оценки параметров терморегуляции рептилий (на примере обыкновенной гадюки, Vipera berus L.) // Принципы экологии. 2020. № 4. С. 88–103. DOI: 10.15393/j1.art.2020.11322
Литвинов Н. А., Четанов Н. А., Ганщук С. В. Принципы оценки термоадаптационных возможностей рептилий / Вестник ТГУ. 2013. Т. 18. Вып. 6. С. 3035–3038.
Мастицкий С. Э., Шитиков В. К. Статистический анализ и визуализация данных с помощью R. М.: ДМК Пресс, 2015. 495 с.
Уланова Е. С. Агрометеорологические условия и урожайность озимой пшеницы. Л.: Гидрометеоиздат, 1975. 303 с.
Черлин В. А. Термобиология рептилий. Общая концепция. СПб.: Русско-Балтийский информационный центр «Блиц», 2012. 362 с.
Шитиков В. К., Розенберг Г. С. Рандомизация и бутстреп: статистический анализ в биологии и экологии с использованием R. Тольятти: Кассандра, 2013. 314 с.
Cumming G., Finch S. Inference by Eye: Confidence Intervals and How to Read Pictures of Data // American Psychologist. 2005. Vol. 60 (2). P. 170–180.
Herczeg G., Saarikivi J., Gonda A., Perälä J., Tuomola A., Merilä J. Suboptimal thermoregulation in male adders (Vipera berus) after hibernation imposed by spermiogenesis. // Biological Journal of the Linnean Society. 2007. No. 92. P. 19–27.
Lourdais O., Guillon M., DeNardo D., Blouin-Demers G. Cold climate specialization: Adaptive covariation between metabolic rate and thermoregulation in pregnant vipers // Physiology & Behavior. 2013. No. 119. P. 149–155.

 © 2011 - 2026
© 2011 - 2026