



Введение
Серая куропатка Perdix perdix (L., 1958) – широко распространенный вид курообразных, численность которого в большинстве стран Западной и Восточной Европы, а также в Северной Америке неуклонно сокращается (Kuijper et al., 2009; BirdLife International, 2015; Johnsgard, 2017). На сегодняшний день известно, что одним из главных факторов, регулирующих численность популяции и определяющих в целом границы ареала птиц, в том числе реинтродуцированной в США серой куропатки, являются погодные условия и климат (Potts, 1980; McCrow, 1982; Böhning-Gaese, Lemoine, 2004). При этом достаточно хорошо описан эффект влияния погоды на выживаемость птенцов серой куропатки, начиная с момента их вылупления (Gates, 1973; Panek, 1992; Potts, 2012; Potts, Aebischer, 1995; Kuijper et al., 2009; Aebischer, Ewald, 2012). Однако практически отсутствуют работы, посвященные влиянию погодно-климатических условий на сроки размножения, пребывания и весенне-осенних сезонных перемещений (региональных миграций) серых куропаток. Подобные исследования единичны и лишь косвенно подтверждают эту связь (Montagna, Meriggi, 1991). Так или иначе, серая куропатка не фигурирует в перечнях из нескольких десятков видов птиц, анализируемых с позиций долговременного влияния погоды и климата на сроки размножения (обзоры: Dunn, Winkler, 2010; Halupka, Halupka, 2017).
Погодные условия, в первую очередь температура и осадки, вносят существенные коррективы в ежегодные флуктуации численности серых куропаток (Potts, 1970; Panek, 1992; Гайдук, Абрамова, 2009; Faragó et al., 2012; Bro et al., 2014; Gruychev, Angelov, 2019), проявляются локально, имеют различную интенсивность и региональные особенности, например, даже на уровне стран Западной и Восточной Европы по мере нарастания континентальности с запада на восток (Tryjanowski et al., 2011). Следовательно, аналогичные региональные различия следует ожидать и при анализе влияния весенних метеопараметров на сроки размножения серых куропаток в различных частях ареала, причем не только в долготном, но и, по-видимому, в широтном направлении (Parmesan, 2007). Так, в Великобритании, по некоторым оценкам, низкие весенние температуры задерживают гнездовой цикл куропаток максимум на 3 дня (Potts, 1970). В то же время данные, полученные на севере Италии, указывают, что в некоторые годы низкие весенние температуры значимо отсрочивают начало откладки яиц и, соответственно, сроки вылупления птенцов (Montagna, Meriggi, 1991).
Мало что известно также о влиянии температуры и осадков на длительность пребывания серых куропаток в районе размножения и на сроки начала их сезонных перемещений в осенний период, что, очевидно, связано с оседлостью серых куропаток в большей части ареала, в частности на территории Европы. Между тем данные по другим группам птиц указывают на неоднозначность влияния потепления климата в Европе на длительность пребывания в районе размножения. Например, мигранты, преимущественно из отряда Воробьинообразные, в фазы потепления климата прибывали раньше в Англию и Прибалтику, но и отлетали осенью раньше, тем самым длительность пребывания в районе размножения существенно не менялась (Cotton, 2003; Соколов, 2006). Другие мигранты, наоборот, в теплые годы прилетали раньше и оставались в районе размножения в Центральной Европе дольше обычного (Berthold, 2001).
Целью настоящей работы является рассмотрение влияния весенних, летних, осенних и частично зимних погодных условий на сроки и длительность пребывания семейных групп серых куропаток, принадлежащих к малоизученному в экологическом отношении подвиду Perdix perdix robusta, в степях Южного Предуралья за долгосрочный период. Для этого была выбрана Буртинская степь – один из участков ФГБУ «Заповедники Оренбуржья» (Оренбургский и Шайтан-Тау), по которому накоплен многолетний статистический материал.
Буртинская степь – это место размножения серых куропаток. Уже с июня здесь начинают встречаться почти исключительно выводки и сопровождающие их взрослые птицы (семейные группы). Рассматриваемый в настоящей работе подвид Perdix perdix robusta (Homeyer et Tancre, 1883), населяющий район исследования (Потапов, 1987), является частично перелетным, и в ноябре начинаются откочевки птиц из данной местности предположительно в южные районы. В малоснежные теплые зимы на участке возможна зимовка лишь незначительной части птиц (Barbazyuk, Chibilyov, 2018). Предполагается, что со второй половины лета идет активное движение выводков через участок в силу его большей биотопической привлекательности, поэтому учетные маршруты отражают ситуацию с ежегодным обилием куропаток не только в Буртинской степи, но и в окружающей местности в целом.
Материалы
Буртинская степь площадью 45 км2 (4500 га) находится в степной зоне Оренбургского Предуралья (координаты кордона 51.228783 N, 56.666500 E), примерно в 30 км от границы с Казахстаном. В 1989 г. вошла в состав заповедника «Оренбургский». Климат – выраженный континентальный с холодной суровой зимой (средняя температура января -15.8 °С) и сухим жарким летом (средняя температура июля +22 °С). Среднегодовое количество осадков 327 мм. Устойчивый снежный покров высотой от 10 до 120 см держится примерно с декабря по конец марта. Участок лежит в подзоне разнотравно-типчаково-ковыльных (настоящих) степей на южных черноземах. Участок представляет собой холмистую открытую степь с мелкими березово-осиновыми рощицами в складках холмов и черноольшаниками по поймам ручьев. Максимальная абсолютная высота на плато Муюлды в центре участка составляет 420.9 м над уровнем моря (Чибилев, 2014). Участок окружают безлесные равнинные пастбища и сельскохозяйственные угодья, значительная часть которых представлена залежами.
Данные по численности серой куропатки в Буртинской степи получены за период 1992–2019 гг. с пропуском 1993 г. Учеты куропаток проводили и проводят до настоящего времени несколько постоянных и бессменных хорошо подготовленных учетчиков, которые являются инспекторами участка.
Методы
Учетные маршруты представляют собой полевые дороги, которые проходят по степным ландшафтам через гнездовые и кормовые стации куропаток. Кроме того, учеты проводили по всему периметру участка, который выделен на местности опашкой. Весь участок находится в условиях строгого заповедного режима, и по полевым дорогам передвигается только патрульная машина.
Интенсивные учеты куропаток на постоянном и второстепенных маршрутах представляли собой комбинацию автомобильных учетов с остановками в постоянных пунктах наблюдения (кордон, ручей Кайнар, плотины Белоглинка и Таволгасай) (рис. 1). Периметр осматривали несколько раз за сезон во время опашки участка. Данная схема учетов не менялась и продолжает сохраняться до настоящего времени. Многолетние личные наблюдения автора в Буртинской степи показали, что малопосещаемые полевые дороги и опашка на заповедном участке, по которым проходили учетные маршруты, являлись местом притяжения выводков серых куропаток. Возможно, это связано с большей доступностью кормов в придорожной полосе, лучшим обзором и контролем выводка взрослыми, более безопасными условиями пребывания с позиций своевременного обнаружения наземных хищников (повышается дистанция обнаружения).
Все данные, полученные для настоящего исследования, представлены в двух наборах. Первый набор данных включает в себя только основной учетный маршрут протяженностью 14.9 км (рис. 1), который целиком осматривается с периодичностью 3–5 раз в неделю с ранней весны до конца года. Второй набор содержит данные с основного маршрута плюс данные с второстепенных полевых дорог (обследуются фрагментарно периодичностью 3–5 раз в неделю, общая протяженность 12.3 км), весь периметр участка (полный осмотр за день осуществляется 2–3 раза в год, общая протяженность 33.5 км), а также другие места Буртинской степи (отдельные встречи). Зимой количество посещений заповедника резко сокращалось до нескольких в месяц. Таким образом, в первом наборе данных учитывались встречи только с основного маршрута, а во втором наборе – все имеющиеся встречи на участке. Основная масса встреч получена в ходе учетов на основном маршруте. Тем не менее данные с других мест участка в отдельные годы дополняли и уточняли общую картину пребывания куропаток на участке, например, существенно расширяя размах встреч куропаток за счет единичных более ранних или более поздних встреч по периферии участка. В статье приводятся отдельно эти два набора данных под названием «маршрут» и «участок».
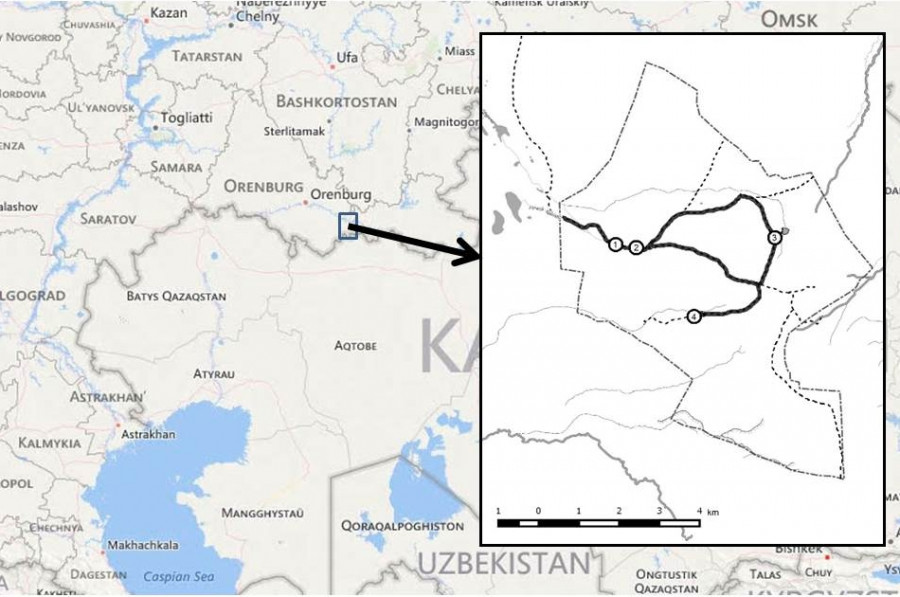
Рис. 1. Район исследования. Толстая черная линия – многолетний основной дорожный маршрут с остановками (протяженность 14.9 км); пунктир – границы заповедника, или периметр (33.5 км), и второстепенные дорожные маршруты в его пределах (12.3 км). Остановки на основном маршруте: 1 – кордон, 2 – ручей Кайнар, 3 – плотина Белоглинка, 4 – плотина Таволгасай
Fig. 1. Study area: Burtinskaya Steppe, Orenburg Nature Reserve, Southern Cis-Urals, Russia. Thick black line indicates the primary main road route with stops (length 14.9 km); dotted line represents the boundaries of Burtinskaya Steppe, or the perimeter (33.5 km), and secondary road routes within its boundaries (12.3 km). Stops on the main route: 1 – ranger station, 2 – Kaynar Spring, 3 – Beloglinka Dam, 4 – Tavolgasay Dam
Ширина учетной полосы не превышала 100 м, по 50 м в каждую сторону от дороги. Во время учетов встреченных птиц подсчитывали на земле или в воздухе при вспугивании стаи с дороги, опашки или в пункте остановки. Фиксировали следующие параметры: количество особей (одиночки, пары или группы более 2 птиц), дата, время и место встречи, по возможности пол, возраст, состояние молодых (бегают, подлетывают, в пуху, оперенные). В зимние месяцы дополнительно фиксировались следы куропаток на снегу и их количество. За один дневной учет могло быть 1–3 и более встреч куропаток. В общей сложности за период 1992–2019 гг. зафиксировано 1332 встречи серых куропаток в Буртинской степи, включая одиночек весной, стаи в зимний сезон и следы птиц на снегу. В настоящей работе акцент сделан на рассмотрении сроков встреч серых куропаток в Буртинской степи, а не на анализе численности птиц. Тем не менее в целях демонстрации общей картины динамики численности за многолетний период в качестве примера приводится средняя численность птиц с июня по декабрь на основании дневных учетов семейных групп на маршруте и на всем участке в перерасчете на квадратный километр от общей площади Буртинской степи.
В исследовании применялся исключительно корелляционный анализ. В связи с распределением дат встреч куропаток по закону, отличному от нормального, использовался непараметрический коэффициент корелляции Спирмена.
В пределах сезона встречи куропаток анализировали на периодах июнь – ноябрь и июнь – декабрь, которые соответствовали началу появления выводков и концу календарной осени, года. Еще один анализируемый период сезона представлял собой полный межгодовой цикл встреч семейных групп куропаток в Буртинской степи: с июня и до конца апреля следующего года (до конца цикла), т. е. до полного прекращения встреч семейных групп в районе исследования, до майского гэпа и начала встреч новых семейных групп в июне.
При построении линейных регрессий метеопараметров и других переменных с указанием коэффициента регрессии (В) проверяли распределение остатков тренда на нормальность по критерию хи-квадрат и Колмогорова – Смирнова, а отсутствие автокорелляции в остатках, в частности, по критерию Дарбина – Уотсона для временных рядов. Непараметрический корелляционный анализ и построение регрессий осуществлялись в программе Statistica 10.
Посуточные метеоданные за 1992–2019 гг. получены в государственной метеостанции (пункт Беляевка), расположенной в 25 км к северо-западу от Буртинской степи.
Для оценки влияния засухи на даты встреч серых куропаток использовался один из отечественных количественных показателей засухи – гидротермический коэффициент Г. Т. Селянинова (ГТК), близкий аналог стандартизованного индекса осадков (SPI) и стандартизованного индекса осадков и испаряемости (SPEI) (Черенкова, Золотокрылин, 2016). ГТК представляет собой относительный показатель увлажненности территории – отношение суммы осадков (R) в мм со средней температурой воздуха выше 10 °С (Т > 10 °С) к сумме средних суточных температур, уменьшенных в 10 раз, за этот же период:
![]()
Расчет ГТК, характеризующего соотношение тепла и влаги, производился на основе отбора среднесуточных температур воздуха выше 10 °С и суточных сумм осадков в эти даты за каждый месяц с мая по август за 28 лет. Согласно Г. Т. Селянинову, для упрощенной оценки режима увлажнения «внутри» вегетационного периода за засушливый период принимается ГТК, равный менее 1.0, а за сухой – менее 0.5 (Грингоф, Павлова, 2013).
Результаты
Многолетнее изменение средней температуры воздуха, осадков и гидротермического коэффициента
В целом можно утверждать, что за исследуемый период времени средняя температура воздуха росла, а количество осадков и ГТК снижались. При этом тенденции были неодинаковые. При анализе изменения средней температуры за отдельные сезоны года линейно устойчиво росли только летние температуры (y = -145.899 + 0.0832x, B = 0.44, R2 = 0.19, F1,26 = 6.13, р = 0.020, n = 28; нормальное распределение остатков, DW = 1.89: автокорреляция отсутствует) и весенние температуры (y = -189.694 + 0.0973x, B = 0.39, R2 = 0.15, F1,26 = 4.55, р = 0.04, n = 28; нормальное распределение остатков, DW = 2.66: автокорреляция не определена) (рис. 2). Наилучшая связь средней температуры воздуха со временем наблюдалась при объединении нескольких периодов года, максимальная связь зафиксирована в апреле – октябре (y = -118.9085 + 0.0667x, B = 0.50, R2 = 0.24, F1,26 = 8.39, р = 0.008, n = 28; нормальное распределение остатков, DW = 1.77: автокорреляция отсутствует). Достоверная линейная зависимость осадков от времени весной и осенью отсутствовала (см. рис. 2) и оказалась достоверной только в июне – августе (y = 125.1675-2.5103x, B = -0.40, R2 = 0.16, F1,26 = 4.82, р = 0.04, n = 28; нормальное распределение остатков, DW = 1.81: автокорреляция отсутствует). В 1992–2019 гг. наблюдалось устойчивое снижение ГТК за весь рассчитываемый период, т. е. с мая по август (y = 118.3481-0.0579x, B = -0.38, R2 = 0.15, F1,26 = 4.49, р = 0.04, n = 28; нормальное распределение остатков, DW = 2.12: автокорреляция отсутствует) (см. рис. 2).
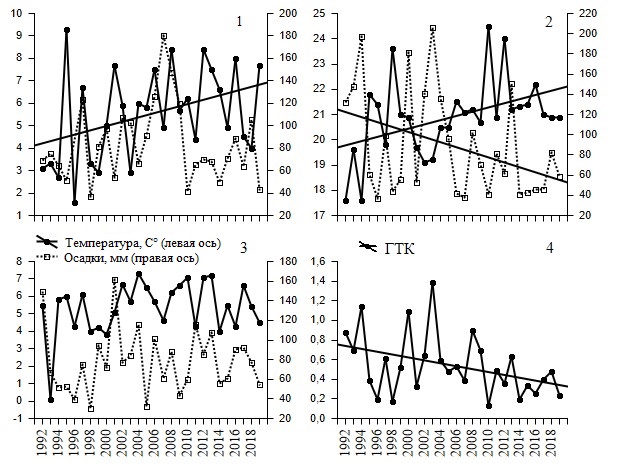
Рис. 2. Средняя температура воздуха (ºС) и сумма осадков (мм) за март – май (1), июнь – август (2), сентябрь – ноябрь (3), а также ГТК за май – август (4) в 1992–2019 гг. в Буртинской степи заповедника «Оренбургский». Для значимых линейных регрессий показан тренд
Fig. 2. Average air temperature (ºС) on left axis and precipitation (mm) on right axis March – May (1), June – August (2), September – November (3), and also HTC May – August (4) in 1992–2019 in Burtinskaya Steppe, Orenburg Nature Reserve. Trend is shown for significant linear regressions
Распределение количества встреч по месяцам, изменение средней плотности птиц и средней даты встреч семейных групп
В 1992–2019 гг. серые куропатки в Буртинской степи в целом встречались на протяжении всех месяцев года. В апреле и мае это были преимущественно одиночки и пары птиц. В июне появлялись первые выводки. Количество встреч семейных групп (выводков со взрослыми) нарастало до октября, затем в ноябре и декабре резко падало: птицы покидали участок. В отдельные теплые годы мелкие группы куропаток (подросшие выводки) держались до февраля – марта, в двух случаях – до первой декады апреля. В мае семейные группы отсутствовали (рис. 3). После появления первых выводков одиночных птиц и пары регистрировали очень редко. Это были имитирующие травму особи, которые отводили опасность от гнезда, птенцов либо, вероятно, особи, иным образом связанные с семейными группами.
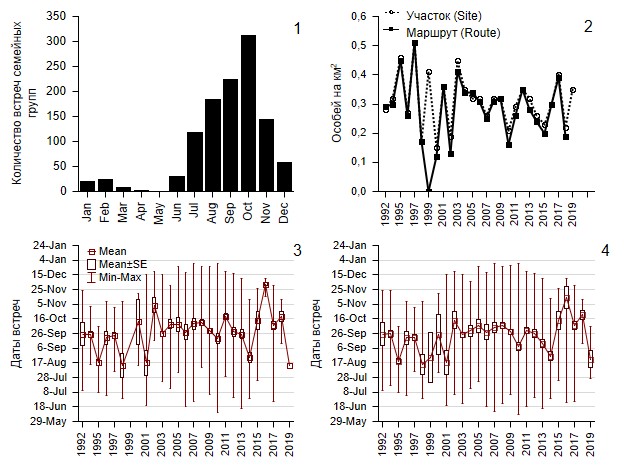
Рис. 3. Распределение количества встреч семейных групп в течение года (с января по декабрь) на участке в 1992–2019 гг. (1). Средняя численность птиц на маршруте и на участке с июня по декабрь в перерасчете на км2 (2). Даты встреч семейных групп на маршруте с июня по декабрь (3). Даты встреч семейных групп на участке с июня по декабрь (4). Данные учетов в Буртинской степи заповедника «Оренбургский», 1992–2019 гг.
Fig. 3. Annual distribution of family group sightings (January–December) for the entire site (1). Average bird density per square kilometer on the route and the entire site from June to December (2). Dates of family group sightings on the route from June to December (3). Dates of family group sightings for the entire site June – December (4). Census data in Burtinskaya Steppe, Orenburg Nature Reserve, 1992–2019
Средняя плотность серых куропаток в Буртинской степи в перерасчете на площадь участка сильно варьировалась по годам от 0 (1999) до 0.51 ± 0.08 (1997) особи/км2 на маршруте и от 0.15 ± 0.03 (2000) до 0.51 ± 0.08 (1997) особи/км2 на всем участке. Максимальные расхождения численности птиц на маршруте и на всем участке зафиксированы в 1999 г.: на маршруте птицы отсутствовали, а за его пределами встретились всего две семейные группы. В 2019 г. на маршруте зафиксировано лишь единичное наблюдение семейной группы в количестве 18 птиц, при этом на всем участке средняя численность составляла 15.6 ± 2.46 (n = 5), или 0.35 ± 0.05 особи/км2 (рис. 3). При этом наблюдалась слабая понижательная, статистически недостоверная линейная динамика средней плотности серых куропаток в Буртинской степи.
За исследуемый период средняя дата встреч серых куропаток до конца календарного года варьировалась в пределах 13 августа (1998) – 2 декабря (2016) на маршруте и в пределах 14 августа (1998) – 14 ноября (2016) на всем участке (см. рис. 3). Как уже отмечено выше, на маршруте в 1999 г. птиц не регистрировали, а в 2019 г. 13 августа зарегистрирована единичная встреча семейной группы. В целом на участке птиц регистрировали во все годы, количество встреч за год было всегда больше одной (см. рис. 3).
Влияние весенне-летних температур, осадков и ГТК на среднюю и медианную дату встреч семейных групп
В табл. 1 и 2 представлены корелляции метеопараметров за разные периоды времени со средней датой встреч и медианой встреч семейных групп серых куропаток на маршруте и на всем участке до конца осени и до конца календарного года.
Таблица 1. Значения корреляции Спирмена между средней температурой воздуха, суммой осадков и ГТК Селянинова и средней датой встреч семейных групп на маршруте (n = 26) и на всем участке (n = 27) до конца осени и до конца года. Данные учетов в Буртинской степи заповедника «Оренбургский», 1992–2019 гг.
| Средняя дата встреч, дней с июня | Период | ||||||||||||||
| Апрель | Апрель – Май | Апрель – Июнь | Апрель – Июль | Апрель – Август | Май | Май – Июнь | Май –Июль | Май – Август | Июнь | Июнь – Июль | Июнь – Август | Июль | Июль – Август | Август | |
| До конца года | Средняя температура воздуха (ºС) | ||||||||||||||
| Маршрут | 0.15 | -0.13 | -0.28 | -0.17 | -0.12 | -0.34 | -0.46 | -0.32 | -0.24 | -0.42 | -0.19 | -0.12 | 0.08 | 0.06 | 0.08 |
| Участок | -0.03 | -0.10 | -0.23 | -0.15 | -0.13 | -0.20 | -0.32 | -0.23 | -0.14 | -0.40 | -0.18 | -0.12 | -0.01 | -0.01 | 0.00 |
| До конца осени | |||||||||||||||
| Маршрут | 0.01 | -0.30 | -0.42 | -0.34 | -0.30 | -0.51 | -0.55 | -0.43 | -0.35 | -0.48 | -0.25 | -0.21 | -0.02 | -0.02 | -0.05 |
| Участок | -0.14 | -0.26 | -0.39 | -0.33 | -0.30 | -0.36 | -0.44 | -0.37 | -0.27 | -0.49 | -0.28 | -0.21 | -0.10 | -0.01 | -0.06 |
| До конца года | Сумма осадков (мм) | ||||||||||||||
| Маршрут | 0.31 | 0.57 | 0.59 | 0.47 | 0.45 | 0.39 | 0.48 | 0.37 | 0.39 | 0.47 | 0.28 | 0.23 | -0.06 | 0.01 | -0.05 |
| Участок | 0.37 | 0.56 | 0.52 | 0.38 | 0.39 | 0.35 | 0.36 | 0.26 | 0.34 | 0.31 | 0.13 | 0.15 | -0.08 | 0.01 | -0.01 |
| До конца осени | |||||||||||||||
| Маршрут | 0.23 | 0.52 | 0.56 | 0.52 | 0.49 | 0.38 | 0.44 | 0.44 | 0.43 | 0.46 | 0.41 | 0.30 | 0.13 | 0.15 | -0.06 |
| Участок | 0.25 | 0.53 | 0.54 | 0.49 | 0.46 | 0.41 | 0.38 | 0.39 | 0.40 | 0.37 | 0.32 | 0.21 | 0.15 | 0.11 | -0.10 |
| До конца года | ГТК | ||||||||||||||
| Маршрут | - | - | - | - | - | 0.34 | 0.46 | 0.33 | 0.36 | 0.49 | 0.30 | 0.24 | -0.07 | 0.04 | -0.07 |
| Участок | - | - | - | - | - | 0.28 | 0.33 | 0.22 | 0.27 | 0.32 | 0.15 | 0.15 | -0.08 | 0.03 | -0.03 |
| До конца осени | |||||||||||||||
| Маршрут | - | - | - | - | - | 0.37 | 0.46 | 0.44 | 0.42 | 0.48 | 0.41 | 0.31 | 0.12 | 0.17 | -0.06 |
| Участок | - | - | - | - | - | 0.39 | 0.38 | 0.37 | 0.35 | 0.38 | 0.31 | 0.23 | 0.15 | 0.13 | -0.10 |
Примечание. Средняя дата встреч рассчитывалась с 1 июня до конца календарного года (июнь – декабрь: с 1-го по 214-й день) и с 1 июня до конца осени (июнь – ноябрь: с 1-го по 183-й день). Корреляции, отмеченные полужирным шрифтом, значимы на уровне р ˂ 0.05, а отмеченные полужирным шрифтом и курсивом значимы на уровне р ˂ 0.01 (то же для табл. 2).
Таблица 2. Значения корреляции Спирмена между средней температурой воздуха, суммой осадков и ГТК Селянинова и медианой даты встреч семейных групп на маршруте (n = 26) и на всем участке (n = 27) до конца осени и до конца года. Данные учетов в Буртинской степи заповедника «Оренбургский», 1992–2019 гг.
| Медиана даты встреч, дней с июня | Период | ||||||||||||||
| Апрель | Апрель – Май | Апрель – Июнь | Апрель – Июль | Апрель – Август | Май | Май –Июнь | Май –Июль | Май – Август | Июнь | Июнь –Июль | Июнь – Август | Июль | Июль – Август | Август | |
| До конца года | Средняя температура воздуха (°С) | ||||||||||||||
| Маршрут | 0.28 | -0.08 | -0.13 | -0.06 | -0.01 | -0.37 | -0.33 | -0.22 | -0.13 | -0.23 | -0.01 | 0.05 | 0.16 | 0.18 | 0.18 |
| Участок | -0.03 | -0.14 | -0.23 | -0.17 | -0.16 | -0.30 | -0.30 | -0.27 | -0.16 | -0.35 | -0.17 | -0.08 | -0.06 | -0.01 | 0.01 |
| До конца осени | |||||||||||||||
| Маршрут | 0.20 | -0.14 | -0.20 | -0.13 | -0.09 | -0.44 | -0.39 | -0.29 | -0.20 | -0.28 | -0.07 | -0.03 | 0.08 | 0.12 | 0.11 |
| Участок | -0.07 | -0.21 | -0.30 | -0.25 | -0.23 | -0.37 | -0.36 | -0.33 | -0.23 | -0.39 | -0.23 | -0.14 | -0.11 | -0.05 | -0.05 |
| До конца года | Сумма осадков (мм) | ||||||||||||||
| Маршрут | 0.31 | 0.44 | 0.49 | 0.42 | 0.37 | 0.33 | 0.40 | 0.36 | 0.28 | 0.40 | 0.29 | 0.16 | -0.01 | -0.04 | -0.13 |
| Участок | 0.37 | 0.57 | 0.53 | 0.44 | 0.40 | 0.42 | 0.38 | 0.34 | 0.33 | 0.29 | 0.17 | 0.11 | -0.03 | -0.02 | -0.09 |
| До конца осени | |||||||||||||||
| Маршрут | 0.23 | 0.46 | 0.48 | 0.46 | 0.41 | 0.32 | 0.35 | 0.38 | 0.32 | 0.33 | 0.31 | 0.19 | 0.09 | 0.06 | -0.11 |
| Участок | 0.25 | 0.59 | 0.56 | 0.48 | 0.44 | 0.44 | 0.39 | 0.37 | 0.36 | 0.29 | 0.21 | 0.13 | 0.03 | 0.02 | -0.10 |
| До конца года | ГТК | ||||||||||||||
| Маршрут | - | - | - | - | - | 0.27 | 0.38 | 0.32 | 0.27 | 0.41 | 0.27 | 0.13 | -0.02 | -0.04 | -0.15 |
| Участок | - | - | - | - | - | 0.35 | 0.36 | 0.31 | 0.29 | 0.30 | 0.17 | 0.10 | -0.02 | -0.01 | -0.10 |
| До конца осени | |||||||||||||||
| Маршрут | - | - | - | - | - | 0.31 | 0.37 | 0.37 | 0.32 | 0.35 | 0.29 | 0.17 | 0.09 | 0.06 | -0.12 |
| Участок | - | - | - | - | - | 0.38 | 0.38 | 0.36 | 0.32 | 0.30 | 0.20 | 0.13 | 0.04 | 0.04 | -0.10 |
При рассмотрении связи средней температуры воздуха с датами встреч семейных групп до конца осени и года ключевым периодом был май – июнь. Повышенные средние температуры в мае – июне способствовали более ранней средней дате встреч куропаток на участке. И наоборот, более прохладный май – июнь приводил к смещению средней даты встреч на более поздние сроки (см. табл. 1). Для медианы получались более строгие значения (см. табл. 2).
Наиболее важным с позиции влияния осадков на средние и медианные даты встреч куропаток оказался период с середины весны до начала лета: апрель – июнь. Именно в этот период наблюдалась самая высокая и достоверная положительная корреляция сумм осадков со средней датой встреч семейных групп серых куропаток в Буртинской степи. Дождливые середина весны и начало лета приводили к смещению средней даты встреч семейных групп куропаток на более поздний период. Начиная с периода апрель – май в таблице значимость и значение корреляции снижаются слева направо. Хорошо заметно, что без весеннего периода летние суммы осадков перестают коррелировать со средней датой встреч (см. табл. 1). Особенно это заметно на примере медианы (см. табл. 2).
При рассмотрении связи гидротермического коэффициента с датами встреч семейных групп куропаток выявлен ключевой период май – июль. Как и в случае с осадками, повышенное увлажнение в мае – июле приводило к смещению средней даты встреч семейных групп на более поздний период (см. табл. 1). При анализе корреляции медианы даты встреч с гидротермическим коэффициентом значимым оказался только июнь. Майские значения корреляции лишь немного не достигли значимого уровня 0.05 (см. табл. 2).
Связь температур, осадков и гидротермического коэффициента с размахом встреч, а также с количеством встреч семейных групп в июне –июле и ноябре–декабре
На графике (см. рис. 3) заметно некоторое расширение размаха встреч в период с 2001 по 2013 г. за счет все большего наращивания ранних июньских и поздних ноябрьских встреч, появления декабрьских встреч. За исследуемый период 1992–2019 гг. первые несколько декабрьских встреч появились только в 2001 г., и до 2013 г. включительно птицы встречались в декабре регулярно. В дальнейшем декабрьские встречи были зарегистрированы только в 2016 г.
Прослеживается некоторая связь размаха с температурой. Размах встреч на маршруте до конца осени (июнь – ноябрь) положительно коррелирует со средней температурой воздуха за тот же период (R = 0.46, р = 0.021, n = 25). Достоверность связи между размахом встреч на всем участке до конца осени (июнь – ноябрь) и средней температурой воздуха за тот же период несколько не достигла минимального критического уровня (R = 0.38, р = 0.052, n = 27). На других отрезках сезона (июнь – декабрь, июнь – апрель) корелляция была низкой и недостоверной. Связь размаха встреч с осадками за различные периоды сезона также оказалась низкой и недостоверной.
В пользу увеличения размаха встреч в сезоны с повышенной средней температурой воздуха говорит и достоверная положительная связь количества встреч семейных групп в июне – июле с температурами в эти и предшествующие периоды, а также положительная связь количества встреч в ноябре – декабре с температурами в эти и предшествующие периоды (табл. 3). Для начала периода появления семейных групп наилучшим результатом оказалась положительная корреляция средних температур в мае – июне с количеством встреч в июне – июле на маршруте: R = 0.52, р = 0.0052, n = 27. В ноябре количество встреч также было выше, если осень была теплой. Наибольшая достоверная связь зафиксирована между средней температурой в сентябре – ноябре и количеством встреч на всем участке в ноябре – декабре: R = 0.44, р = 0.023, n = 27 (см. табл. 3).
Таблица 3. Значения корреляции Спирмена между средней температурой воздуха и количеством встреч семейных групп на краевых периодах размаха встреч (июнь – июль, ноябрь – декабрь) до конца года, на маршруте (n = 27) и на всем участке (n = 27). Данные учетов в Буртинской степи заповедника «Оренбургский», 1992–2019 гг.
| Температура | Количество встреч: маршрут/участок | ||||
| Июнь | Июнь – Июль | Ноябрь | Ноябрь – Декабрь | Декабрь | |
| Апрель | 0.07/0.14 | -0.02/0.11 | – | – | – |
| Апрель – Май | 0.26/0.31 | 0.28/0.42 | – | – | – |
| Апрель – Июнь | 0.29/0.20 | 0.39/0.42 | – | – | – |
| Апрель – Июль | – | 0.31/0.39 | – | – | – |
| Май | 0.26/0.20 | 0.41/0.45 | – | – | – |
| Май – Июнь | 0.32/0.12 | 0.52/0.44 | – | – | – |
| Май – Июль | – | 0.43/0.42 | – | – | – |
| Июнь | 0.24/0.01 | 0.42/0.31 | – | – | – |
| Июнь – Июль | – | 0.27/0.25 | – | – | – |
| Сентябрь | – | – | 0.36/0.38 | 0.33/0.37 | 0.16/0.25 |
| Сентябрь – Октябрь | – | – | 0.15/0.21 | 0.14/0.21 | 0.17/0.24 |
| Сентябрь – Ноябрь | – | – | 0.40/0.41 | 0.41/0.44 | 0.34/0.41 |
| Сентябрь–Декабрь | – | – | – | 0.20/0.22 | 0.04/0.10 |
| Октябрь | – | – | -0.15/-0.09 | -0.17/-0.12 | -0.02/-0.02 |
| Октябрь – Ноябрь | – | – | 0.26/0.27 | 0.27/0.29 | 0.25/0.30 |
| Октябрь – Декабрь | – | – | – | 0.24/0.23 | 0.10/0.15 |
| Ноябрь | – | – | 0.38/0.37 | 0.41/0.41 | 0.31/0.36 |
| Ноябрь – Декабрь | – | – | – | 0.23/0.21 | 0.03/0.07 |
| Декабрь | – | – | – | – | -0.17/-0.16 |
Примечание. Корреляции, отмеченные полужирным шрифтом, значимы на уровне р ˂ 0.05, а отмеченные полужирным шрифтом и курсивом значимы на уровне р ˂ 0.01. То же для табл. 4.
На фоне слабой связи размаха встреч с осадками за весь период наблюдений в 1992–2019 гг. сумма осадков отрицательно влияла на количество встреч куропаток только в начале сезона (таб.. 4). Так, выпадение повышенного количества осадков в апреле – мае приводило к снижению количества встреч куропаток в июне – июле на маршруте (R = -0.40, p = 0.037, n = 27) и на всем участке (R = -0.43, р = 0.023, n = 27). Количество выпавших осадков осенью значимо не влияло на количество встреч куропаток в данный период. Отрицательная связь осадков с количеством встреч повышается в декабре (R = -0.26 на маршруте и R = -0.23 на участке), но остается недостоверной (см. табл. 4).
Таблица 4. Значения корреляции Спирмена между суммой осадков и количеством встреч семейных групп на краевых периодах размаха встреч (июнь – июль, ноябрь – декабрь) до конца года, на маршруте (n = 27) и на всем участке (n = 27). Данные учетов в Буртинской степи заповедника «Оренбургский», 1992–2019 гг.
| Осадки | Количество встреч: маршрут/участок | ||||
| Июнь | Июнь – Июль | Ноябрь | Ноябрь – Декабрь | Декабрь | |
| Апрель | -0.01/-0.06 | -0.23/-0.26 | – | – | – |
| Апрель – Май | -0.30/-0.16 | -0.40/-0.43 | – | – | – |
| Апрель – Июнь | -0.31/-0.11 | -0.36/-0.32 | – | – | – |
| Апрель – Июль | – | -0.41/-0.33 | – | – | – |
| Май | -0.28/-0.13 | -0.22/-0.25 | – | – | – |
| Май – Июнь | 0.32/-0.12 | -0.26/-0.21 | – | – | – |
| Май – Июль | – | -0.35/-0.25 | – | – | – |
| Июнь | -0.34/-0.09 | -0.26/-0.16 | – | – | – |
| Июнь – Июль | – | -0.39/-0.24 | – | – | – |
| Сентябрь | – | – | 0.11/0.04 | 0.20/0.14 | 0.22/0.22 |
| Сентябрь – Октябрь | – | – | 0.18/0.12 | 0.21/0.17 | 0.12/0.15 |
| Сентябрь – Ноябрь | – | – | 0.21/0.19 | 0.24/0.21 | 0.13/0.18 |
| Сентябрь – Декабрь | – | – | – | -0.01/-0.04 | -0.06/-0.01 |
| Октябрь | – | – | 0.15/0.15 | 0.15/0.16 | 0.03/0.08 |
| Октябрь – Ноябрь | – | – | 0.15/0.18 | 0.14/0.15 | 0.01/0.06 |
| Октябрь – Декабрь | – | – | – | -0.17/-0.17 | -0.22/-0.17 |
| Ноябрь | – | – | -0.07/-0.01 | -0.01/0.01 | 0.06/0.08 |
| Ноябрь – Декабрь | – | – | – | -0.27/-0.25 | -0.18/-0.15 |
| Декабрь | – | – | – | – | -0.26/-0.23 |
Линейные зависимости средней даты, медианы, размаха встреч от времени и метеопараметров в различные периоды сезона
Следствием описанных выше явлений, в первую очередь, вероятно, устойчивого роста температуры, являются выявленные некоторые слабые линейные зависимости изучаемых параметров от времени и температуры.
Линейная зависимость средней и медианной даты встреч от времени усиливается при рассмотрении отрезков сезона в направлении: июнь – ноябрь (до конца осени), июнь – декабрь (до конца года) и становится достоверной в июне – апреле (до конца цикла). Коэффициент регрессии был достоверным только для средней даты встреч на всем участке: B = 0.43, R2 = 0.18, F1,23 = 5.20, p = 0.03, n = 25 (распределение остатков нормальное, DW = 1.96: автокорреляция отсутствует) и для медианной даты встреч на всем участке B = 0.46, R2 = 0.21, F1,23 = 6.07, p = 0.02, n = 25 (распределение остатков нормальное, DW = 2.10: автокорреляция отсутствует). Для других периодов сезона значимость коэффициента линейной регрессии немного не достигала уровня значимости 0.05 (рис. 4, табл. 5).
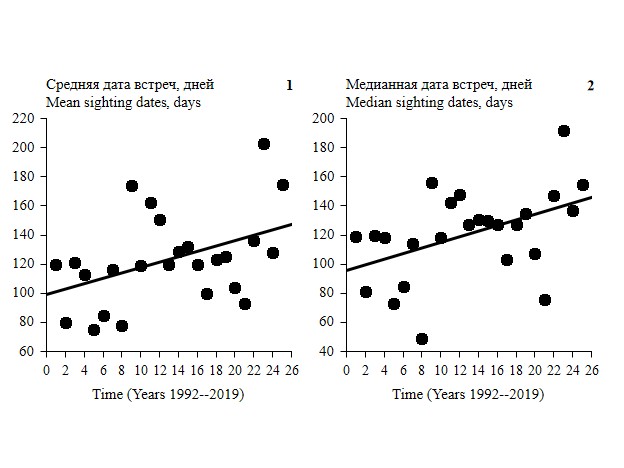 Рис. 4. Линейная зависимость средней (1) и медианной (2) даты встреч от времени в июне – апреле (до конца цикла) на всем участке. Средняя и медианная дата встреч рассчитывалась с 1 июня по 30 апреля следующего года (до конца встреч семейных групп). Данные учетов в Буртинской степи заповедника «Оренбургский», 1992–2019 гг.
Рис. 4. Линейная зависимость средней (1) и медианной (2) даты встреч от времени в июне – апреле (до конца цикла) на всем участке. Средняя и медианная дата встреч рассчитывалась с 1 июня по 30 апреля следующего года (до конца встреч семейных групп). Данные учетов в Буртинской степи заповедника «Оренбургский», 1992–2019 гг.
Fig. 4. Linear dependence of average (1) and median (2) sighting dates on time, June – April (until the end of the cycle) for the entire site. Average and median sighting dates were calculated from June 1 to April 30 of the following year (until the end of family group sightings). Census data in Burtinskaya Steppe, Orenburg Nature Reserve, 1992–2019
В то же время линейная зависимость размаха встреч от времени на различных отрезках сезона не подтверждается. Она оказалась слабой и недостоверной (см. табл. 5). Линейная связь между средней датой встреч и средней температурой воздуха также оказалась слабой и недостоверной; аналогичные результаты получены и для медианы. Линейная связь между размахом встреч и температурой снижается от осени к зиме и к концу цикла, т. е. влияние температуры на размах встреч резко падает к зиме и далее. При этом коэффициент линейной регрессии оказался значимым только для пары переменных «Размах встреч до конца года на маршруте & температура» (B = 0.44, R2 = 0.19, F1,23 = 5.51, p = 0.03, n = 25) (см. табл. 5).
Линейная связь между средней датой встреч и количеством осадков на различных отрезках сезона отсутствует (см. табл. 5). Для медианы получаются похожие значения. Линейная связь между количеством осадков и размахом встреч отсутствует на всех отрезках сезона. Как и в случае с осадками, линейная связь между ГТК и средней датой встреч, а также между ГТК и размахом встреч отсутствует (см. табл. 5).
Таблица 5. Некоторые линейные регрессии, описывающие влияние метеопараметров (средняя температура, осадки, ГТК) и времени Т (годы) на среднюю дату, медиану и размах встреч семейных групп серых куропаток на маршруте и на всем участке за различные периоды сезона. Данные учетов в Буртинской степи заповедника «Оренбургский», 1992–2019 гг.
| Линейная регрессия | Статистические параметры |
| (зависимая и независимая переменные) | |
| Средняя дата встреч до конца осени на маршруте & время Т | B = 0.27, R2 = 0.07, F1,24 = 1.91, p = 0.18, n = 26 |
| Средняя дата встреч до конца осени на участке & время Т | B = 0.31, R2 = 0.10, F1,25 = 2.72, p = 0.11, n = 27 |
| Средняя дата встреч до конца года на маршруте & время Т | B = 0.25, R2 = 0.06, F1,24 = 1.62, p = 0.22, n = 26 |
| Средняя дата встреч до конца года на участке & время Т | B = 0.33, R2 = 0.11, F1,25 = 3.08, p = 0.09, n = 27 |
| Средняя дата встреч до конца цикла на маршруте & время Т | B = 0.37, R2 = 0.14, F1,22 = 3.57, p = 0.07, n = 24 |
| Средняя дата встреч до конца цикла на участке & время Т | B = 0.43, R2 = 0.18, F1,23 = 5.20, p = 0.03, n = 25 |
| Медиана даты встреч до конца осени на маршруте & время Т | B = 0.26, R2 = 0.07, F1,24 = 1.69, p = 0.21, n = 26 |
| Медиана даты встреч до конца осени на участке & время Т | B = 0.30, R2 = 0.09, F1,25 = 2.52, p = 0.12, n = 27 |
| Медиана даты встреч до конца года на маршруте & время Т | B = 0.27, R2 = 0.07, F1,24 = 1.89, p = 0.18, n = 26 |
| Медиана даты встреч до конца года на участке & время Т | B = 0.33, R2 = 0.11, F1,25 = 3.06, p = 0.09, n = 27 |
| Медиана даты встреч до конца цикла на маршруте & время Т | B = 0.35, R2 = 0.13, F1,22 = 3.17, p = 0.09, n = 24 |
| Медиана даты встреч до конца цикла на участке & время Т | B = 0.46, R2 = 0.21, F1,23 = 6.07, p = 0.02, n = 25 |
| Размах встреч до конца осени на маршруте & время Т | B = 0.09, R2 = 0.01, F1,23 = 0.18, p = 0.68, n = 25 |
| Размах встреч до конца осени на участке & время Т | B = 0.17, R2 = 0.03, F1,25 = 0.75, p = 0.40, n = 27 |
| Размах встреч до конца года на маршруте & время Т | B = 0.10, R2 = 0.01, F1,23 = 0.23, p = 0.64, n = 25 |
| Размах встреч до конца года на участке & время Т | B = 0.15, R2 = 0.02, F1,25 = 0.55, p = 0.46, n = 27 |
| Размах встреч до конца цикла на маршруте & время Т | B = 0.21, R2 = 0.04, F1,22 = 0.99, p = 0.33, n = 24 |
| Размах встреч до конца цикла на участке & время Т | B = 0.16, R2 = 0.03, F1,24 = 0.66, p = 0.42, n = 26 |
| Средняя дата встреч до конца осени на маршруте & температура | B = -0.01, R2 = 0.00, F1,24 = 0.01, p = 0.94, n = 26 |
| Средняя дата встреч до конца осени на участке & температура | B = -0.01, R2 = 0.00, F1,25 = 0.00, p = 0.95, n = 27 |
| Средняя дата встреч до конца года на маршруте & температура | B = -0.15, R2 = 0.02, F1,24 = 0.58, p = 0.45, n = 26 |
| Средняя дата встреч до конца года на участке & температура | B = -0.16, R2 = 0.03, F1,25 = 0.69, p = 0.41, n = 27 |
| Средняя дата встреч до конца цикла на маршруте & температура | B = -0.17, R2 = 0.03, F1,22 = 0.64, p = 0.43, n = 24 |
| Средняя дата встреч до конца цикла на участке & температура | B = -0.19, R2 = 0.03, F1,23 = 0.82, p = 0.38, n = 25 |
| Размах встреч до конца осени на маршруте & температура | B = 0.39, R2 = 0.15, F1,23 = 4.03, p = 0.06, n = 25 |
| Размах встреч до конца осени на участке & температура | B = 0.33, R2 = 0.11, F1,25 = 3.07, p = 0.09, n = 27 |
| Размах встреч до конца года на маршруте & температура | B = 0.44, R2 = 0.19, F1,23 = 5.51, p = 0.03, n = 25 |
| Размах встреч до конца года на участке & температура | B = 0.18, R2 = 0.03, F1,25 = 0.80, p = 0.38, n = 27 |
| Размах встреч до конца цикла на маршруте & температура | B = 0.15, R2 = 0.02, F1,22 = 0.48, p = 0.49, n = 24 |
| Размах встреч до конца цикла на участке & температура | B = -0.12, R2 = 0.01, F1,23 = 0.33, p = 0.57, n = 25 |
| Средняя дата встреч до конца осени на маршруте & осадки | B = 0.20, R2 = 0.04, F1,24 = 1.05, p = 0.32, n = 26 |
| Средняя дата встреч до конца осени на участке & осадки | B = 0.15, R2 = 0.02, F1,25 = 0.59, p = 0.45, n = 27 |
| Средняя дата встреч до конца года на маршруте & осадки | B = 0.18, R2 = 0.03, F1,24 = 0.83, p = 0.37, n = 26 |
| Средняя дата встреч до конца года на участке & осадки | B = 0.10, R2 = 0.01, F1,25 = 0.27, p = 0.61, n = 27 |
| Средняя дата встреч до конца цикла на маршруте & осадки | B = 0.08, R2 = 0.01, F1,22 = 0.15, p = 0.71, n = 24 |
| Средняя дата встреч до конца цикла на участке & осадки | B = 0.06, R2 = 0.00, F1,23 = 0.08, p = 0.78, n = 25 |
| Размах встреч до конца осени на маршруте & осадки | B = -0.13, R2 = 0.02, F1,23 = 0.40, p = 0.53, n = 25 |
| Размах встреч до конца осени на участке & осадки | B = 0.08, R2 = 0.01, F1,25 = 0.18, p = 0.68, n = 27 |
| Размах встреч до конца года на маршруте & осадки | B = -0.18, R2 = 0.03, F1,23 = 0.79, p = 0.38, n = 25 |
| Размах встреч до конца года на участке & осадки | B = 0.00, R2 = 0.00, F1,25 = 0.00, p = 0.99, n = 27 |
| Размах встреч до конца цикла на маршруте & осадки | B = -0.05, R2 = 0.00, F1,22 = 0.06, p = 0.81, n = 24 |
| Размах встреч до конца цикла на участке & осадки | B = -0.02, R2 = 0.00, F1,23 = 0.01, p = 0.93, n = 25 |
| Средняя дата встреч до конца осени на маршруте & ГТК | B = 0.26, R2 = 0.07, F1,24 = 1.73, p = 0.20, n = 26 |
| Средняя дата встреч до конца осени на участке & ГТК | B = 0.30, R2 = 0.09, F1,25 = 2.40, p = 0.13, n = 27 |
| Размах встреч до конца осени на маршруте & ГТК | B = -0.17, R2 = 0.03, F1,23 = 0.69, p = 0.41, n = 25 |
| Размах встреч до конца осени на участке & ГТК | B = 0.03, R2 = 0.00, F1,25 = 0.02, p = 0.90, n = 27 |
Примечание. р – значения, отмеченные жирным шрифтом, близки к уровню р = 0.05, а отмеченные жирным шрифтом и курсивом соответствуют уровню р ˂ 0.05
Обсуждение
Полученные данные указывают на то, что серые куропатки подвида Perdix perdix robusta реагируют как на температуру, так и осадки в первой половине сезона размножения. Жаркая погода в мае – июне способствует более ранним встречам выводков в Буртинской степи, т. е. способствует ускорению всего процесса размножения: более ранней откладке яиц, более раннему вылуплению птенцов. В то время как повышенное количество осадков в апреле – июне и повышенное увлажнение в мае – июле приводят к смещению средней даты встреч семейных групп на более поздние сроки, т. е., другими словами, затягивают вылупление птенцов.
Предполагается, что холода в конце весны и начале лета приводят либо к повторным кладкам и затягиванию всего процесса размножения, либо птицы приступают к размножению позже, ожидая более комфортных условий. Полученные данные согласуются с общими тенденциями реакции птиц на повышение температуры в начале сезона размножения. Сейчас хорошо известно, что большинство птиц северного умеренного пояса приступают к откладке яиц раньше в теплые весны, чем в холодные, как следствие глобального изменения климата (Crick, Sparks, 1999; Both, Artemyev et al., 2004; Соколов, 2006; Carey, 2009; Dunn, Winkler, 2010). Серые куропатки проявляют чувствительность к эстремальным погодным условиям во время откладки яиц. В теплые годы скорость откладки яиц в сутки у серых куропаток увеличивается (Potts, 1980). Одним из объяснений возможной задержки гнездового сезона может быть то, что низкие температуры весной замедляют рост травы, что приводит к плохой маскировке гнезд с кладками и высокой доле их разорения хищниками (Meriggi et al., 1990).
Аналогичным образом дожди в середине весны и начале лета могут препятствовать cтадиям размножения, вызывать оставление кладок самками или приводить к гибели недавно вылупившихся птенцов, что, в свою очередь, также, возможно, затягивает появление выводков в районе исследования. Так, во Франции сильный дождь в летнее время иногда приводил к оставлению кладок самками серых куропаток, но лишь в отдельных случаях (Bro et al., 2014). На северо-востоке США у воробьинообразных большее количество дождевых осадков обычно было связано с более поздним появлением молодых и более поздней готовностью самок к размножению (McDermott, 2016).
Как показали исследования, средняя температура воздуха может оказывать и другое воздействие. Наметились некоторые тенденции расширения размаха встреч в период с июня по ноябрь в наиболее теплые годы. Раннее тепло в середине весны и начале лета способствует появлению выводков уже с начала июня, а теплые ноябрь и декабрь позволяют все большему количеству семейных групп задерживаться на заповедном участке и, очевидно, в целом в данной местности вплоть до конца года. Количество встреч семейных групп было достоверно выше весной и осенью в более теплые годы. Вместе с тем при рассмотрении линейной зависимости температуры и размаха встреч можно отметить, что она, как и в случае корелляции, резко снижается от осени к зиме и еще сильнее к концу цикла.
В настоящем исследовании выявлена также небольшая положительная линейная зависимость средней и медианы даты встреч от времени, которая усиливается при рассмотрении отрезков сезона в следующем направлении: июнь – ноябрь (до конца осени), июнь – декабрь (до конца года), июнь – апрель (до конца цикла). Другими словами, за исследуемый период средняя и медианная даты встреч семейных групп серых куропаток все больше смещаются к концу календарного года.
Отсутствие устойчивого снежного покрова при положительных температурах в октябре, ноябре и декабре, по-видимому, улучшает кормовые условия в районе исследования. Далее при сохранении благоприятных условий незначительная часть птиц остаются на участке до конца марта или апреля. В холодные годы период комфортного пребывания выводков на участке сокращается: появление первых семейных групп начинается только с конца июня, а в октябре птицы уже покидают участок из-за усиливающихся холодов в сочетании с обильными осадками в виде снега, из-за формирования устойчивого снежного покрова и, видимо, ухудшения условий питания. Подобные процессы наиболее ярко проявлялись в отдельные годы в начале исследуемого отрезка времени.
Что касается влияния осадков на количество встреч птиц в заповеднике, отрицательная связь двух этих параметров отмечена только в весенне-летний период. Отрицательная связь отсутствует осенью и начинает повышаться только к зиме, по-прежнему оставаясь недостоверной. В декабре – марте численность серых куропаток в Буртинской степи в значительной степени определяется выпавшими осадками в виде снега: в многоснежные зимы птицы либо полностью откочевывают, либо присутствуют в очень минимальном числе (Barbazyuk, Chibilyov, 2018). Все более длительные задержки и зимовки отдельных выводков на участке в условиях устойчивого потепления и как следствие снижения средней высоты снежного покрова (Barbazyuk, Chibilyov, 2018) в районе исследования могут объяснить и другую наметившуюся тенденцию: все большее смещение средней и медианной даты встреч к концу года со временем. Эта тенденция наиболее отчетливо проявляется при рассмотрении полного цикла встреч семейных групп куропаток, с момента их появления в июне и до полного распада выводков в апреле – мае следующего года. При этом, как указывалось выше, линейная связь между средней, а также медианной датой встреч и количеством осадков отсутствует. Очевидно также, что необходимо дальнейшее изучение связи размаха и средней даты встреч птиц в заповеднике с температурой и осадками на более длительном временном ряду.
Заключение
В районе исследования устойчиво растут весенне-летние температуры, наблюдается снижение количества осадков и ГТК за тот же период.
Температура, осадки и увлажнение в середине весны – начале лета значимо влияют на среднюю и медианную даты встреч семейных групп куропаток в Буртинской степи позже в сезон. Повышенные средние температуры в конце весны и начале лета сдвигают среднюю и медианную даты встреч на более ранний срок на отрезке сезона до конца календарного года, а повышенные количество осадков и увлажнение в апреле – июне, наоборот, способствуют сдвигу средней и медианной дат встреч семейных групп на более поздние сроки.
Отмечена некоторая тенденция расширения размаха встреч семейных групп в наиболее теплые годы с момента их появления в июне и до конца осени. Расширение происходит за счет все более ранних встреч в июне и увеличения количества встреч в ноябре, а также в декабре. Теплая погода в начале лета и в конце осени – начале зимы может способствовать расширению периода благоприятных условий нахождения птиц в данной местности. Раннее тепло в конце весны и начале лета способствует появлению выводков уже с начала июня, а теплые сентябрь – ноябрь и декабрь, отсутствие устойчивого снежного покрова как результат потепления позволяют все большему количеству семейных групп задерживаться на заповедном участке и в целом в данной местности вплоть до конца года.
В то же время связь размаха встреч куропаток с осадками на исследуемом отрезке времени отсутствует. Повышенные суммы осадков осенью значимо не влияли на количество встреч в этот период. Отрицательная связь количества встреч и осадков начинает усиливаться только в декабре, но остается недостоверной. Только весной дождливые апрель и май приводят к значимому снижению встреч куропаток в июне – июле, т. е. могут в какой-то степени влиять на размах встреч в целом.
Тенденция смещения средней даты и медианы встреч семейных групп куропаток на конец года за рассматриваемый многолетний период времени, по-видимому, также связана с улучшением благоприятных условий, в первую очередь кормовых. Потепление климата, снижение сумм осадков в осенне-зимнее время, снижение высоты снежного покрова, уменьшение продолжительности устойчивого снежного покрова повышают доступность кормовой базы куропаток. Однако прямой линейной зависимости между средней датой встреч и количеством осадков на различных отрезках сезона не выявлено. Для более уверенного подтверждения линейных тенденций связи размаха встреч, средней даты встреч с метеопараметрами и временем требуется более продолжительный временной ряд и больший объем данных.
Библиография
Гайдук В. Е., Абрамова И. В. Экология птиц юго-запада Беларуси. Неворобьинообразные. Брест: БрГУ, 2009. 300 с.
Грингоф И. Г., Павлова В. Н. Основы сельскохозяйственной метеорологии. Обнинск: ФГБУ ВНИИГМИ-МЦД, 2013. 384 с.
Потапов Р. Л. Серая куропатка // Птицы СССР. Курообразные, журавлеобразные. Л.: Наука, 1987. С. 24–39.
Соколов Л. В. Влияние глобального потепления климата на сроки миграции и гнездования воробьиных птиц в ХХ веке // Зоологический журнал. 2006. Т. 85, № 3. С. 317–341.
Черенкова Е. А., Золотокрылин А. Н. О сравнимости некоторых количественных показателей засухи // Фундаментальная и прикладная климатология. 2016. Т. 2. С. 79–94. DOI: 10.21513/2410-8758-2016-2-79-94.
Чибилев А. А. Заповедник «Оренбургский»: история создания и природное разнообразие. Екатеринбург: УИПЦ, 2014. 140 с.
Aebischer N. J., Ewald J. A. The grey partridge in the UK: population status, research, policy and prospects // Animal Biodiversity and Conservation. 2012. Vol. 35 (2). P. 353–362. DOI: 10.32800/abc.2012.35.0353.
Barbazyuk E. V., Chibilyov A. A. The Impact of Winter Meteorological Parameters on Abundance of the Grey Partridge Perdix perdix in the Southern Ural Steppe // Doklady Biological Sciences. 2018. Vol. 480. P. 103–106. DOI: 10.1134/S0012496618030080.
Bro E., Brillard J., Millot F. Impact of heavy rains and wet cold weather on Grey partridge Perdix perdix clutch desertion and breeding success // Alauda. 2014. Vol. 82 (1). P. 51–62.
Berthold P. Avian migration. A general survey, Oxford, UK: Oxford University Press, 2001. 266 р.
BirdLife International. Perdix perdix. The IUCN Red List of Threatened Species 2016. URL: https://dx.doi.org/10.2305/IUCN.UK.2016-3.RLTS.T22678911A85929015.en (дата обращения: 18.01.2021).
Both C., Artemyev A. V., Blaauw B., Cowie R. J., Dekhuijzen A. J., et al. Large-scale geographical variation confirms that climate change causes birds to lay earlier // Proceedings of the Royal Society of London. Series B. 2004. Vol. 271. Р. 1657–1662. DOI: 10.1098/rspb.2004.2770.
Böhning-Gaese K. & Lemoine N. Importance of climate change for ranges, communities and conservation of birds // Advances in Ecological Research. 2004. Vol. 35. Р. 211–236. DOI: 10.1016/S0065-2504(04)35010-5.
Carey C. The impacts of climate change on the annual cycles of birds // Philosophical Transactions of the Royal Society B. 2009. Vol. 364. Р. 3321–3330. DOI: 10.1098/rstb.2009.0182.
Cotton P. A. Avian migration phenology and global climate change // Proceedings of the National Academy of Sciences of the United States of America. 2003. Vol. 100 (21). Р. 12 219–12 222. DOI: 10.1073/pnas.1930548100.
Crick H. Q. P., Sparks T. H. Climate change related to egg-laying trends // Nature. 1999. Vol. 399. P. 423. DOI: 10.1038/20839.
Dunn P., Winkler D. W. Effects of climate change on timing of breeding and reproductive success in birds // Møller A. P., Fiedler W., Berthold P (eds.). Effects of climate change on birds, Oxford, UK: Oxford University Press, 2010. Р. 113–128.
Faragó S., Dittrich G., Horváth-Hangya K., Winkler D. Twenty years of the grey partridge population in the LAJTA Project (Western Hungary) // Animal Biodiversity and Conservation. 2012. Vol. 35 (2). P. 311–319. DOI: 10.32800/abc.2012.35.0311.
Gates J. M. Gray partridge ecology in southeast-central Wisconsin. Madison: Wisconsin Department of Natural Resources. Technical Bulletin 70, 1973. 8 p.
Gruychev G. V., Angelov E. E. Density of Grey Partridge (Perdix perdix Linnaeus, 1785) population in Sakar Mountain (SE Bulgaria) and the effect of weather and habitats // Ecologia Balkanica. 2019. Vol. 11 (1). P. 51–62.
Halupka L, Halupka K. The effect of climate change on the duration of avian breeding seasons: a meta-analysis // Proceedings of the Royal Society B. 2017. Vol. 284. P. 2017–1710. DOI: dx.doi.org/10.1098/rspb.2017.1710.
Johnsgard P. A. The North American Quails, Partridges, and Pheasants. Lincoln: Zea E-Books. 58, 2017. 131 p. DOI: 10.13014/K2G15Z1P.
Kuijper D. P. J., Oosterveld E., Wymenga E. Decline and potential recovery of the European grey partridge (Perdix perdix) population – a review // European Journal of Wildlife Research. 2009. Vol. 55. P. 455–463. DOI: 10.1007/s10344-009-0311-2.
McCrow V. P. Gray partridge habitat use and nesting biology in north-central Iowa: Retrospective Theses and Dissertations. 7516. Ames: Iowa State University, 1982. 239 p. DOI: 10.31274/rtd-180813-5931.
McDermott M. E., DeGroote L. W. Long-term climate impacts on breeding bird phenology in Pennsylvania, USA // Global Change Biology. 2016. Vol. 22. P. 3304–3319. DOI: 10.1111/gcb.13363.
Meriggi A., Montagna D., Zacchetti D., Matteucci C., Toso S. Population dynamics of the gray partridge in relation to agriculture and weather in northern Italy // Church K. E., Warner R. E., Brady S. J. (eds.). Perdix V: gray partridge and ring-necked pheasant workshop. Kansas Dept. of Wildlife and Parks, Emporia, 1990. Р. 241–256.
Montagna D., Meriggi A. Population dynamics of grey partridge (Perdix perdix) in northern Italy // Italian Journal of Zoology. 1991. Vol. 58 (2). P. 151–155. DOI: 10.1080/11250009109355746.
Panek M. The effect of environmental factors on survival of grey partridge (Perdix perdix) chicks in Poland during 1987–89 // Journal of Applied Ecology. 1992. Vol. 29 (3). P. 745–750. DOI: 10.2307/2404484.
Parmesan C. Influences of species, latitudes and methodologies on estimates of phenological response to global warming // Global Change Biology. 2007. Vol. 13. P. 1860–1872. DOI: 10.1111/j.1365-2486.2007.01404.x.
Potts G. R. Recent changes in the farmland fauna with special reference to the decline of the Grey Partridge // Bird Study. 1970. Vol. 17 (2). P. 145–166. DOI: 10.1080/00063657009476266.
Potts G. R. The effects of modern agriculture, nest predation and game management on the population ecology of partridges (Perdix perdix and Alectoris rufa) // Advances in Ecological Research. 1980. Vol. 11. P. 1–79. DOI: 10.1016/S0065-2504(08)60266-4.
Potts G. R. Partridges – countryside barometer. London: The new naturalist library. Collins, 2012. 465 p.
Potts G. R., Aebischer N. J. Population dynamics of the grey partridge Perdix perdix 1793–1993: monitoring, modelling and management // Ibis. 1995. Vol. 137. P. 29–37. DOI: 10.1111/j.1474-919X.1995.tb08454.x.
Tryjanowski P., Hartel T., Báldi A., Szymański P., Tobolka M., et al. Can research in Western Europe identify the fate of farmland bird biodiversity in Eastern Europe? // Acta Ornithologica. 2011. Vol. 46 (1). P. 79–90. DOI: 10.3161/000164511X589857.
References
Aebischer N. J., Ewald J. A. The grey partridge in the UK: population status, research, policy and prospects // Animal Biodiversity and Conservation. 2012. V. 35 (2). P. 353–362. DOI: 10.32800/abc.2012.35.0353
Barbazyuk E. V., Chibilyov A. A. The Impact of Winter Meteorological Parameters on Abundance of the Grey Partridge Perdix perdix in the Southern Ural Steppe // Doklady Biological Sciences. 2018. V. 480. P. 103–106. DOI: 10.1134/S0012496618030080
Bro E., Brillard J., Millot F. Impact of heavy rains and wet cold weather on Grey partridge Perdix perdix clutch desertion and breeding success // Alauda. 2014. V. 82 (1). P. 51–62.
Berthold P. Avian migration. A general survey, Oxford, UK: Oxford University Press, 2001. 266 р.
BirdLife International. 2016. Perdix perdix. The IUCN Red List of Threatened Species 2016. URL: https://dx.doi.org/10.2305/IUCN.UK.2016-3.RLTS.T22678911A85929015.en (дата обращения 18 января 2021).
Both C., Artemyev A. V., Blaauw B., Cowie R. J., Dekhuijzen A. J., et al. Large-scale geographical variation confirms that climate change causes birds to lay earlier // Proceedings of the Royal Society of London. Series B. 2004. V. 271. Р. 1657–1662. DOI: 10.1098/rspb.2004.2770
Böhning-Gaese K. & Lemoine N. Importance of climate change for ranges, communities and conservation of birds // Advances in Ecological Research. 2004. V. 35. Р. 211–236. DOI: 10.1016/S0065-2504(04)35010-5
Carey C. The impacts of climate change on the annual cycles of birds // Philosophical Transactions of the Royal Society B. 2009. V. 364. Р. 3321–3330. DOI:10.1098/rstb.2009.0182
Cherenkova E.A., Zolotokrylin A.N. // On the comparability of some quantitative indicators of drought. Fundamental and Applied Climatology. 2016. V. 2. P. 79–94. DOI:10.21513/2410-8758-2016-2-79-94
Chibilyov А.А. Orenburg Reserve: History of Creation and Natural Diversity. Yekaterinburg: UIPC, 2014. 140 p.
Cotton P. A. Avian migration phenology and global climate change // Proceedings of the National Academy of Sciences of the United States of America. 2003. V. 100 (21). Р. 12 219–12 222. DOI: 10.1073/pnas.1930548100
Crick H. Q. P., Sparks T. H. Climate change related to egg-laying trends // Nature. 1999. V. 399. P. 423. DOI:10.1038/20839
Dunn P., Winkler D. W. Effects of climate change on timing of breeding and reproductive success in birds // Møller A. P., Fiedler W., Berthold P (eds). Effects of climate change on birds, Oxford, UK: Oxford University Press, 2010. Р. 113–128.
Faragó S., Dittrich G., Horváth–Hangya K., Winkler D. Twenty years of the grey partridge population in the LAJTA Project (Western Hungary) // Animal Biodiversity and Conservation. 2012. V. 35 (2). P. 311–319. DOI: 10.32800/abc.2012.35.0311
Gaiduk V.Е., Abramova I.V. Ecology of birds of the south-west of Belarus. Non-passerine birds. Brest: Brest State University, 2009. 300 p.
Gates J. M. Gray partridge ecology in southeast-central Wisconsin. Madison: Wisconsin Department of Natural Resources. Technical Bulletin 70, 1973. 8 p.
Gringof I.G., Pavlova V.N. Fundamentals of Agricultural Meteorology. Obninsk: FGBU VNIIGMI-MCD, 2013. 384 p.
Gruychev G. V., Angelov E. E. Density of Grey Partridge (Perdix perdix Linnaeus, 1785) population in Sakar Mountain (SE Bulgaria) and the effect of weather and habitats // Ecologia Balkanica. 2019. V. 11 (1). P. 51–62.
Halupka L, Halupka K. The effect of climate change on the duration of avian breeding seasons: a meta-analysis // Proceedings of the Royal Society B. 2017. V. 284. P. 20171710. DOI: dx.doi.org/10.1098/rspb.2017.1710
Johnsgard P. A. The North American Quails, Partridges, and Pheasants. Lincoln: Zea E-Books. 58, 2017. 131 p. DOI: 10.13014/K2G15Z1P
Kuijper D. P. J. , Oosterveld E., Wymenga E. Decline and potential recovery of the European grey partridge (Perdix perdix) population – a review // European Journal of Wildlife Research. 2009. V. 55. P. 455–463. DOI: 10.1007/s10344-009-0311-2
McCrow V. P. Gray partridge habitat use and nesting biology in north-central Iowa: Retrospective Theses and Dissertations. 7516. Ames: Iowa State University, 1982. 239 p. DOI: 10.31274/rtd-180813-5931
McDermott M. E., DeGroote L. W. Long-term climate impacts on breeding bird phenology in Pennsylvania, USA // Global Change Biology. 2016. V. 22. P. 3304–3319. DOI: 10.1111/gcb.13363
Meriggi A., Montagna D., Zacchetti D., Matteucci C., Toso S. Population dynamics of the gray partridge in relation to agriculture and weather in northern Italy // Church K. E., Warner R. E., Brady S. J. (eds.). Perdix V: gray partridge and ring-necked pheasant workshop. Kansas Dept. of Wildlife and Parks, Emporia, 1990. Р. 241–256.
Montagna D., Meriggi A. Population dynamics of grey partridge (Perdix perdix) in northern Italy // Italian Journal of Zoology. 1991. V. 58 (2). P. 151–155. DOI: 10.1080/11250009109355746
Panek M. The effect of environmental factors on survival of grey partridge (Perdix perdix) chicks in Poland during 1987–89 // Journal of Applied Ecology. 1992. V. 29 (3). P. 745–750. DOI: 10.2307/2404484
Parmesan C. Influences of species, latitudes and methodologies on estimates of phenological response to global warming // Global Change Biology. 2007. V. 13. P. 1860–1872.
DOI:10.1111/j.1365-2486.2007.01404.x
Potapov R.L. The Grey Partridge. In: Birds of the USSR. Galliformes, Gruiformes. Leningrad: Nauka, 1987. P. 24–39.
Potts G. R. Recent changes in the farmland fauna with special reference to the decline of the Grey Partridge // Bird Study. 1970. V. 17 (2). P. 145–166. DOI: 10.1080/00063657009476266
Potts G. R. The effects of modern agriculture, nest predation and game management on the population ecology of partridges (Perdix perdix and Alectoris rufa) // Advances in Ecological Research. 1980. V. 11. P. 1–79. DOI: 10.1016/S0065-2504(08)60266-4
Potts G. R. Partridges – countryside barometer. London: The new naturalist library. Collins, 2012. 465 p.
Potts G. R., Aebischer N. J. Population dynamics of the grey partridge Perdix perdix 1793–1993: monitoring, modelling and management // Ibis. 1995. V. 137. P. 29–37. DOI: 10.1111/j.1474-919X.1995.tb08454.x
Sokolov L.V. The impact of global warming on the timing of migration and breeding of passerine birds in the 20th century // Russian Zoological Journal. 2006. V. 85 (3). P. 317–341.
Tryjanowski P., Hartel T., Báldi A., Szymański P., Tobolka M., et al. Can research in Western Europe identify the fate of farmland bird biodiversity in Eastern Europe? // Acta Ornithologica. 2011. V. 46 (1). P. 79–90. DOI: 10.3161/000164511X589857
Благодарности
Автор выражает благодарность коллективу инспекторов участка "Буртинская степь" заповедника "Оренбургский" В. Ф. Шпанагелю, В. В. Шпанагелю, Н. А. Савоскину, В. И. Ершову за сбор полевого материала и техническое содействие при проведении собственных полевых исследований. Статья подготовлена в рамках темы государственного задания Института степи ОФИЦ УрО РАН № ГР АААА-А21-121011190016-1.

 © 2011 - 2026
© 2011 - 2026