



Введение
Глобальное изменение климата и техногенное загрязнение входят в число основных факторов снижения продуктивности, устойчивости и гибели лесов (Grey, Deneke, 1986; Allen et al., 2010; Gauthier et al., 2015; Schou et al., 2015; Brandt et al., 2016; Kuznetsova et al., 2019 и др.). Среднерусская лесостепь до ХХ в. характеризовалась благоприятными природно-климатическими условиями и довольно однородным лесным покровом, которые поддерживались климатом и лесами. Снижение лесистости и глобальное изменение климата существенно затронули лесостепные экосистемы. Повысилась вероятность их перехода в более низкое жизненное состояние и гибели (Richardson et al., 2007; Reyer et al., 2015; Johnstone et al., 2016). Развитие лесостепной биоклиматической системы ускорилось в направлении остепнения. Снизилась устойчивость лесов, конкурентоспособность со степной растительностью, зафиксирована дестабилизация сосновых лесов (Kuznetsova et al., 2019).
Развитие сложных систем, к числу которых относятся лесные экосистемы, находится под контролем мощных регуляторных и адаптивных механизмов (Bak, 1996). Считается, что морфологическим изменениям предшествуют биохимические и физиологические перестройки, вызванные воздействием антропогенных и климатических факторов (Yordanov et al., 2000; Popec et al., 2018). Показано, что в местах техногенного загрязнения у сосны смещается соотношение белкового и небелкового азота в пользу последнего (Mikhailova, 2000), изменяется метаболический статус насаждений (Придача и др., 2011). В засуху, когда экологические проблемы наиболее обострены, изменения водного статуса и метаболических процессов способствуют адаптации и выживанию вида (Чиркова, 2002). Однако данные уровней изменчивости нормы реакции и особенностей взаимодействия вегетативной и генеративной сферы в зависимости от места произрастания популяций сосны и напряженности засухи отсутствуют.
Цель исследования – изучить изменчивость морфологических и физиолого-биохимических признаков и механизмов выживания деревьев природных и урбанизированных экосистем сосны обыкновенной во время умеренной и сильной засухи ex situ и ex vivo. Данное сообщение является продолжением предыдущей работы (Кузнецова, Клушевская, 2020). Новым является моделирование в лабораторном эксперименте условий умеренной и сильной «засухи», описание преобразований адаптивной нормы реакции сосны в зависимости от напряженности засухи и жизненного состояния насаждения.
Материалы
Объектами исследования служили два 55–57-летних насаждения сосны обыкновенной (Pinus sylvestris L.) Воронежской области. Контролем служил Ступинский тест-объект (Рамонский район, лесные культуры, 30 деревьев) – типичное по вегетативной и семенной продуктивности насаждение, произрастающее рядом с Воронежским биосферным заповедником. Лесные культуры произрастают на территории, которая относится к числу эталонных земель с ненарушенным почвенным покровом (ТУМ А2, сосняк травяной). Опытный объект «Московский проспект» – лесная защитная полоса в черте г. Воронежа и вдоль крупной автомагистрали М4 «Дон». Коэффициент эмиссионной нагрузки в районе исследований десятикратно (К = 268) превышает средний уровень Воронежской области (Куролап и др., 2010). С 1987 г. в ЦЧР произошло 8 весенне-летних засух разной напряженности: 2001, 2014, 2019 гг. – слабые, 1995 г. – умеренная, 1991, 2007, 2012 гг. – сильные, 2010 г. – экологической катастрофы (Кузнецова, 2019). Их характеристика приведена в предыдущей работе (Кузнецова, 2019).
Методы
Мониторинг систем семенного размножения ступинской популяции проводили в условиях полевого эксперимента. Урожайность сосны оценивали по двум признакам семенной продуктивности: полнозернистость семян (%) и число семян на шишку (шт.) в оптимальные годы (2008, 2013, 2019 гг.), умеренную 1995 г. и сильную засуху 2007 г. Полнозернистость вычисляли как соотношение числа полных семян к общему их числу (полные + пустые).
Для физиолого-биохимических исследований в мае 2013 г. отобраны побеги 2-го года жизни и проведен анализ хвои 20 деревьев в эксперименте: 1) свежесобранные пробы; 2) умеренный стресс «засуха» (10 суток при +23 °С); 3) сильный гидротермический стресс «засуха + температура» (7 суток при 45 оС) (Клушевская, Кузнецова, 2016). Все растения лабораторного эксперимента являлись частью выборки 30 модельных деревьев опытного и контрольного объектов. Погодные условия на момент сбора образцов соответствовали региональной норме. Для имитации засухи в лабораторных условиях использовали 0.6 М раствор маннита, который растения не метаболизируют (Деменко и др., 2010). Исследования водного режима (дефицит влаги, общее количество влаги, количество коллоидно-связанной воды) проводили по методике Х. Н. Починка (1976). Содержание свободного пролина определяли методом Bates et al. (1973) в воздушно-сухих пробах. Для статистической обработки данных использовали программу Statistica 6. Сравнение выборок по физиолого-биохимическим показателям хвои проводились с использованием непараметрического Х-критерия Ван-дер-Вардена.
Результаты
Ступинский тест-объект
Генеративная сфера сосны обыкновенной очень чувствительна к состоянию среды обитания, часто используется как биоиндикатор мест экологического неблагополучия (Лесные экосистемы…, 1990; Micieta, Murin, 1998; Alaquori et al., 2020). В предыдущей работе показано, что ступинская популяция сосны представляет равновесную систему (Кузнецова, Клушевская, 2020). В ходе мониторинга систем семенного размножения 8 засушливых лет определено, что признаки семенной продуктивности имеют разную природу, внутрипопуляционную структуру и чувствительность к засухе (Кузнецова, 2019). Данные показателей биопродуктивности шишек в засушливые 1995 и 2007 гг. по сравнению с региональной нормой приведены в таблице. Установлено, что порог чувствительности признака полнозернистости – слабая засуха. В оптимальные годы распределение деревьев по классам продуктивности представляет асимметричную кривую с модой в области 80 %. В умеренную засуху 1995 г. уровень полнозернистости снизился вдвое (с 81.2 до 44.3 %), коэффициент корреляции (CV) повысился в 5 раз. Различия с оптимальным годом статистически достоверны (p < 0.01). Произошла внутрипопуляционная дифференциация деревьев на устойчивые, средние и чувствительные к засухе формы. В сильную засуху 2007 г. наблюдалось почти 3-кратное (32.1 %) снижение урожайности сосны. Достоверность различий между годами достигла более высокого уровня значимости (p < 0.001). Распределение деревьев стало асимметричным по отношению к оптимальным годам. Число семян на шишку, как более устойчивый признак, отреагировало только на сильную засуху (см. таблицу). В умеренную засуху его величина оставалась на уровне оптимальных лет – 21.2 шт. В сильную засуху наблюдалось существенное (p < 0.05) снижение количества семян в шишках (с 24.1 до 15.2 шт.), вариабельность признака повысилась с 27.1 до 60.8 %.
Показатели биопродуктивности шишек сосны обыкновенной Ступинского тест-объекта и Московского проспекта в оптимальном 2020 г., умеренную 1995 г. и сильную засуху 2007 г.
| Категория года | Полнозернистость семян, % | CV, % | Число семян на шишку, шт. | CV, % |
| Ступинский тест-объект | ||||
| Среднее за оптимальные годы | 81.2 ± 1.0 | 9.2 | 24.1 ± 1.2 | 27.1 |
| 1995 г., умеренная засуха | 44.3 ± 3.4** | 46 | 21.2 ± 1.4 | 36.5 |
| 2007 г., сильная засуха | 32.1 ± 3.5*** | 67.7 | 15.2 ± 1.5* | 60.8 |
| Московский проспект | ||||
| 2020 г., оптимальный год | 69.1 ± 3.0 | 19.6 | 16.4 ± 1.4 | 45.8 |
| 1995 г., умеренная засуха | 67.1 ± 3.6 | 27.9 | 16.0 ± 1.3 | 43 |
| 2007 г., сильная засуха | 58.4 ± 4.4* | 36.8 | 14.9 ± 1.4 | 45.8 |
* Примечание. Различия между годами достоверны при * – p < 0.05; ** – p < 0.01; *** – p < 0.001.
Моделирование умеренной и сильной «засухи» в лабораторном эксперименте ex vivo продиктовано тем, что за 34 года мониторинга в регионе произошла всего одна умеренная засуха (1995 г.). Анализируемые признаки по-разному отреагировали на смену гидротермического режима (Клушевская, Кузнецова, 2016). Так, средний показатель содержания влаги в образцах хвои оптимального года составляет 55.3 %. Индуцированный стресс снизил его уровень почти вдвое. Величина дефицита влаги при умеренном стрессе повысилась в два раза (с 8.6 до 15.3 %) и стала еще выше – 24.7 % при сильном гидротермическом. Умеренный стресс вызвал 3-кратное повышение содержания коллоидно-связанной воды (с 19.1 до 56.6 %), а при сильном ее количество увеличилось в 1.5 раза. Коэффициенты вариации признака варьировали в очень узких пределах 27.2, 23.3, 27.9 % (оптимальный год, умеренная, сильная засуха соответственно). Различия с контролем в данных вариантах эксперимента статистически достоверны (p < 0.05). Аналогичный отклик имел место по содержанию пролина. При умеренном стрессе его количество повысилось в 4–6 раз, при сильном – только в 1.4 раза. Из-за сильной изменчивости признака (55.9 %) различия в варианте сильная «засуха» недостоверны.
При сравнении пределов размаха нормы реакции признаков ступинской популяции по шкале «норма – умеренная – сильная засуха», несмотря на различия их природы и специфику условий ex vivo и ex situ, выявлен однотипный отклик на стрессовую нагрузку (рисунок). Установлено, что размах варьирования показателей полнозернистости представляет следующий ряд: 46.5–85.0–70.9 %, минимальный уровень – оптимальные годы, максимальный – умеренная засуха (см. рисунок, а). Анализ амплитуды второго признака показал, что значения по числу семян у деревьев колеблются в относительно небольших пределах: 64.0, 65.8, 4.0 % (см. рисунок, б). При этом максимальная изменчивость наблюдается в оптимальные годы, минимальная – в сильную засуху (см. рисунок, б). Показатели содержания влаги обнаруживают аналогичную, как и по полнозернистости, последовательность: 16.0–52.8–22.2 % (см. рисунок, в). Контроль (свежесобранная хвоя) характеризуется минимальным уровнем изменчивости, достигая максимума при умеренном стрессе. Другие показатели имеют тот же алгоритм (см. рисунок, г, д, е): дефицит влаги 7.3–15.8–12.5 %; содержание коллоидно-связанной воды 16.9–52.8–22.2 %; пролин 2.7–23.1–4.7 мкМ/г.
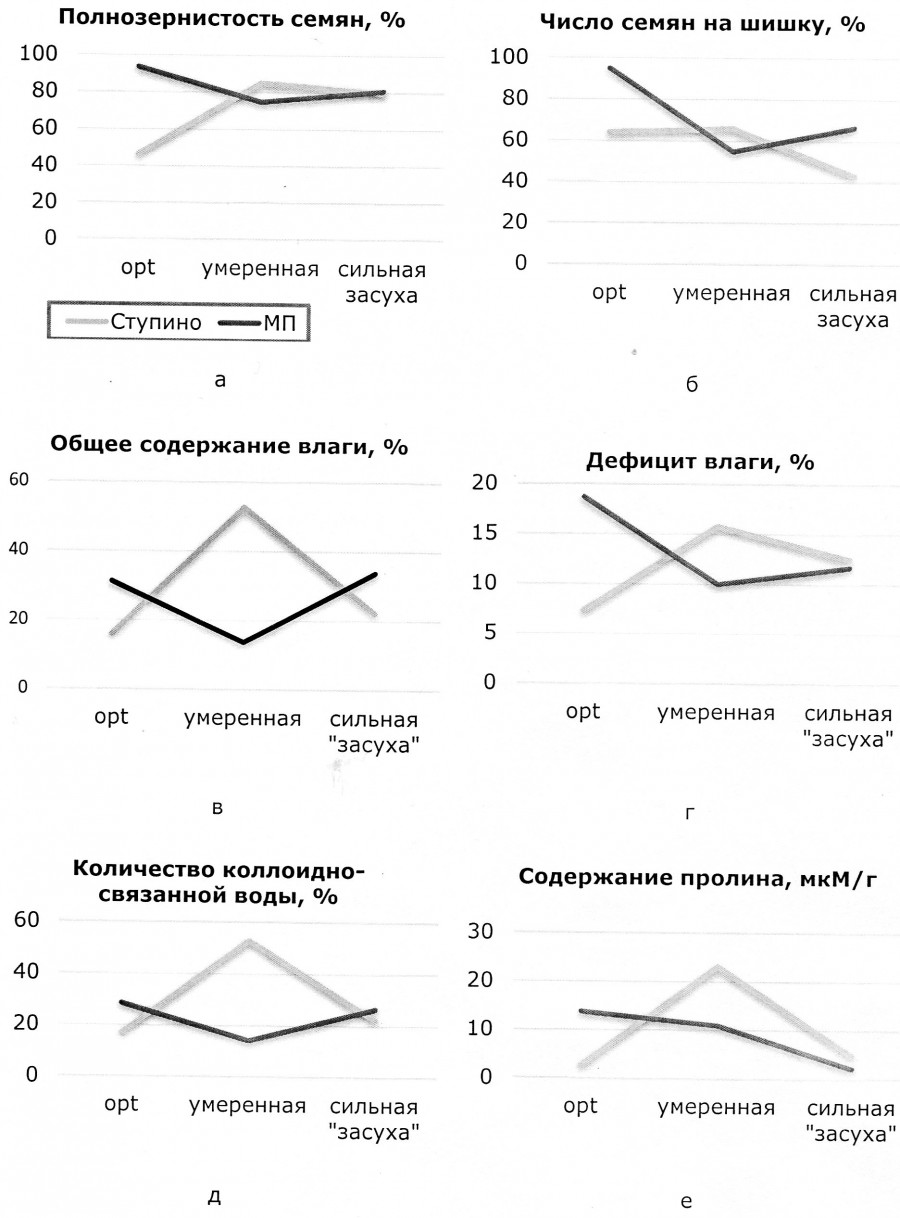
Амплитуда изменчивости нормы реакции у сосны обыкновенной Ступинского тест-объекта в оптимальные годы, умеренную и сильную засуху по признакам: а – полнозернистость семян; б – число семян на шишку; в – общее содержание влаги; г – дефицит влаги; д – количество коллоидно-связанной воды, е – содержание пролина
Amplitude of reaction norm variability in Scots pine of the Stupinsky test-object in optimal years, moderate and strong drought by the traits: a – seed fullness; б – total (full + empty) number of seeds per cone; в – total moisture content; г – moisture deficiency; д – amount of colloid-bound water; е – proline content
Анализ амплитуды изменчивости шести генеративных и вегетативных признаков сосны Ступинского тест-объекта в условиях ex situ и ex vivo показал, что общей реакцией пяти из них является максимальное расширение нормы реакции при умеренном стрессе и сужение при сильном. Исключение составляет показатель «число семян на шишку» (см. рисунок, б), что обусловлено его биологией: широкая норма реакции в оптимальные годы, низкая чувствительность к гидротермическому стрессу и однонаправленный сдвиг реализованной нормы реакции в засуху.
Объект «Московский проспект»
Модельный объект произрастает на техногенно загрязненной территории и находится в более низком жизненном состоянии – слабо неравновесной системы (Кузнецова, Клушевская, 2020). При изучении стресс-реакции установлено, что его деревья иначе реагируют на смену гидротермического режима в условиях ex situ и ex vivo. Отклик на засуху сильно сглажен. В засуху 2007 г. модальное значение полнозернистости семян снизилось всего на 15.5 % – с 69.1 % (0.0–93.7 %) до 58.4 % (5.5–86.1 %) (см. таблицу). Число семян уменьшилось на 9.2 % – с 16.4 шт. (0.0–37.4 семян) до 14.9 шт. (4.2–37.1 семян). По признакам семенной продуктивности достоверных различий между оптимальным 2020 г. и засушливым 2007 г. не выявлено. В умеренную засуху 1995 г. оба показателя остались на уровне оптимальных лет. Одной из особенностей техногенной популяции являлась повышенная изменчивость признаков как в оптимальные, так и в засушливые годы. Внутрипопуляционная дифференциация деревьев на контрастные по засухоустойчивости группы отсутствовала.
Анализ содержания влаги в хвое при умеренном и сильном стрессе показал, что изначально высокие значения признака снижаются в 4–5 раз (с 60.5 до 11.2 и 13.82 % соответственно). Уровень коллоидно-связанной воды в варианте умеренного стресса уменьшается более чем вдвое (с 75.3 до 33.3 %) и опускается до 30.4 % после сильного. Дефицит влаги возрастает вдвое, и его величина сопоставима при воздействии двух видов стресса (29.2 и 29.7 %). Повышенное содержание пролина в свежесобранных образцах при умеренном стрессе снижается с 7.4 до 5.1 мкМ/г, что в два раза меньше, чем у деревьев контроля. При сильном стрессе его количество уменьшается до 2.3 мкМ/г. Исходно достаточно широкая область распределения признака (1.5–15.4) сужается за счет понижения границы верхних значений при сильном стрессе в 3.2 раза (1.1–3.3), при умеренном в 1.5 раза (1.5–12.4). Различия между данными показателями эксперимента достоверны по всем физиолого-биохимическим признакам.
У опытной популяции выявлен противоположный контролю алгоритм изменчивости признаков. Минимальный размах приходится на умеренный погодный стресс: полнозернистость 93.7–74.0–80.6 %; число семян 74.8–54.8–66.4 % (см. рисунок, а, б). Аналогичная реакция наблюдалась по физиолого-биохимическим показателям (см. рисунок, в–е): содержание влаги 31.3–13.8–33.9 %; дефицит влаги 18.7–10.0–11.7 %; коллоидно-связанной воды 28.5–14.2–26.6 % (см. рисунок, в, г, д). Отличия выявлены только по реакции пролина на индуцированную «засуху» – 13.9, 11.0, 2.2 мкМ/г (см. рисунок, е). По-видимому, его образование возможно лишь до определенной стрессовой нагрузки, за пределами которой тождественность биосинтеза силе воздействия нарушается.
Единый для вегетативной и генеративной сферы тип изменчивости морфологических и физиолого-биохимических признаков подтверждает, что ступинская популяция и популяция Московского проспекта находятся в разном жизненном состоянии. По-видимому, повышенный внутренний дисбаланс деревьев опытной популяции, который присущ всем неравновесным системам, в засушливые годы предохраняет генетический материал формирующихся семян от разрушения.
Обсуждение
На классических моделях показано, что во время формирования популяций устанавливается состояние равновесия, которому соответствует наиболее оптимальная структура распределения растений по их микросостояниям (Bak, 1996). В зоне видового оптимума большинство сосновых лесов на экологически благоприятной территории представляют равновесные системы. У ступинской популяции равновесие в оптимальные годы обеспечивает устойчивость и сбалансированное развитие вегетативной и генеративной сфер большинства деревьев.
В засуху стабильность равновесных процессов нарушается: уменьшается годовой прирост, интенсивность семеношения, гомеостаз внутренней среды и т. д. (Чиркова, 2002; Чудинова, Орлова, 2006; Vacchiano et al., 2012; Кузнецова, 2019 и др.). Если погодный стресс не превышает допустимую нагрузку, большинство растений способно перестроиться и подключить защитные механизмы. Поскольку в равновесных системах стресс-реакция может протекать в прямом и обратном направлениях, то на следующий год после засухи они возвращаются к региональной норме.
Общей закономерностью проявления стресса является повышенный уровень изменчивости большинства экологически зависимых количественных признаков, что подтвердилось и в нашем эксперименте ex situ и ex vivo. Самый большой размах изменчивости ступинской популяции наблюдался в умеренную засуху. По-видимому, сосновые леса в состоянии равновесия имеют максимальный выбор возможностей для адаптации при предельном расширении нормы реакции. Сильная засуха ведет к более глубокой разбалансировке процессов, редукции урожая и нарушению генетического материала семян. Поскольку в равновесных системах стресс-реакция может протекать в прямом и обратном направлении, то на следующий год после засухи урожайность всегда возвращалась к региональной норме (≈ 80 %).
Согласно литературным данным, лесные экосистемы достаточно устойчивы к набору основных региональных стрессоров и после их воздействия способны восстанавливать свою структуру и функции (Richardson et al., 2007; Johnstone et al., 2016). Выживание растений в аномальные годы происходит с участием двух механизмов – чуткого реагирования на стресс или автономизации (нечувствительности к нему) (Суходолец, 1998). В засуху создается новая внутрипопуляционная структура, при которой на первое место выходят групповые связи и отношения между генотипами. В равновесных системах направление и сдвиг нормы реакции представляет упорядоченный процесс, который протекает на базе неспецифической и специфической реакции, зависит от категории засухи, природы и чувствительности генотипа к погодному стрессу. Происходит дифференциация на засухоустойчивые, средние и чувствительные формы. Однотипная групповая трансформация наблюдалась нами в разных популяциях сосны (зона видового оптимума, экологически благоприятная территория), что свидетельствует об экологической и генетической обусловленности процесса.
Иная ситуация наблюдается на техногенно загрязненной территории, где биология вида Pinus sylvestris и среда его обитания совпадают лишь частично и лесная экосистема представляет слабо неравновесную систему. Прежде чем перейти к рассмотрению особенностей реакции опытного объекта на засуху, отметим, что неравновесные популяции часто встречаются на территории ареала (болота, высокогорья, антропогенно нарушенные территории, северная и южная границы ареала). В засуху сосновые леса испытывают одновременное воздействие нескольких стрессоров. В оптимальные годы отличительной чертой сосны Московского проспекта является широкая норма реакции морфологических и физиолого-биохимических признаков (Кузнецова, Клушевская, 2020). Онтогенез протекает в условиях постоянных энергетических потерь, связанных с детоксикацией внутренней среды и поддержанием устойчивости неравновесных процессов, что, в свою очередь, снижает жизненный статус, устойчивость и продолжительность жизни растений.
По-видимому, в засуху у опытной популяции, в целом слабо неравновесной системы, существуют состояния локального равновесия, обеспечивающие ее выживание в силу специфики неравновесного состояния. Считается, что дополнительный стресс на фоне уже действующего техногенного прессинга ведет к развитию комплекса приспособительных реакций, трансформации физиолого-биохимических и других процессов, обеспечивающих функционирование и выживание растений в крайне неблагоприятных условиях (Безель и др., 1994; Sakugawa, Cape, 2007; Hamanishi, Campbell, 2011; Масленников и др., 2018; Popec et al., 2018). Согласно полученным данным, генеративная сфера техногенной популяции не отреагировала на умеренную засуху, и наблюдался слабый (15.5 %) отклик на сильную. По сравнению со Ступино выявлен противоположный тип изменчивости признаков по шкале «оптимальные годы – умеренная – сильная засуха». По-видимому, исходно широкие индивидуальные нормы реакции, в засуху сужаясь и перекрываясь, стабилизируют состав и свойства внутренней среды, благодаря которым нивелируется отклик растений на погодный стресс, поддерживаются основные функции их жизнеобеспечения. Это дало нам основание предположить, что механизм выживания техногенной популяции в засуху связан со стабилизацией внутренней среды деревьев, что помогает им пережить не только погодный стресс, но и сохранить способность к половой репродукции на уровне, близком к оптимальным годам.
Заключение
Морфологические и физиолого-биохимические показатели объектов из экологически благоприятных (Ступинский тест-объект) и техногенно загрязненных (Московский проспект) мест обитания достоверно отличаются по реакции на засуху в условиях ex situ и in vivo.
Общей закономерностью реакции вегетативной и генеративной сфер сосны Ступинского тест-объекта является увеличение изменчивости признаков: максимальное расширение реализованной нормы реакции при умеренном гидротермическом стрессе и сужение при сильном. Стресс-реакция представляет упорядоченный процесс, который протекает в рамках равновесного состояния популяционной экосистемы и направлен на адаптацию генотипического состава семян к погодному стрессу.
Выживание сосны Московского проспекта во многом обеспечивается спецификой слабо неравновесного состояния, которое характеризуется пониженной вегетативной и семенной продуктивностью в оптимальные годы, определенной стрессоустойчивостью и набором приспособительных физиолого-биохимических реакций в засуху. Отклик генеративной сферы деревьев на погодный стресс или отсутствует (умеренная засуха), или слабый (сильная засуха). Отличительной особенностью физиолого-биохимических показателей хвои является пониженная изменчивость при умеренном гидротермическом стрессе и близкая к оптимальным годам вариабельность признаков при его усилении. По-видимому, адаптационные механизмы в засуху реализуются в двух направлениях: у вегетативной сферы их действие направлено на поддержание гомеостаза внутренней среды деревьев, а ее стабильность, в свою очередь, предохраняет генетический материал формирующихся семян от полного разрушения.
Библиография
Безель В. С., Большаков В. Н., Воробейчик Е. Л. Популяционная экотоксикология . М.: Наука, 1994. 80 с.
Деменко В. И., Шестибратов К. А., Лебедев В. Г. Укоренение – ключевой этап размножения растений in vitro // Известия ТСХА. 2010. № 1. С. 73–85.
Клушевская Е. С., Кузнецова Н. Ф. Оценка устойчивости сосны обыкновенной к засухе по физиологическим характеристикам хвои // Лесоведение. 2016. № 3. C. 216–222.
Кузнецова Н. Ф. Засухи в лесостепной зоне Центрально-Черноземного района и критерии оценки их интенсивности // Известия Саратовского университета. Новая серия. Серия: Науки о Земле. 2019. Т. 19, вып. 3. С. 142–148. DOI: 10.18500/1819-7663-2019-19-3-142-148.
Кузнецова Н. Ф., Клушевская Е. С. Смена жизненного состояния как способ выживания Pinus sylvestris L. на техногенно загрязненной территории // Принципы экологии. 2020. № 2. С. 40–47. DOI: 10.15393/j1.art.2020.10322.
Куролап С. А., Нестеров Ю. А., Епринцев С. А. Типизация территории Воронежской области по уровню техногенного воздействия на среду обитания // Вестник Воронежского государственного университета. Серия: География. Геоэкология. 2010. № 1. С. 5–11.
Лесные экосистемы и атмосферное загрязнение / Под ред. А. К. Алексеева. Л.: Наука, 1990. 200 с.
Починок Х. Н. Методы биохимического анализа растений . Киев: Наукова думка, 1976. 334 с.
Придача Б. В., Сазонова Т. А., Таланова Т. Ю., Ольчев А. В. Морфофизиологическая реакция Pinus sylvestris L. и Picea obovata Ledeb. при техногенном воздействии в условиях северо-запада России // Экология. 2011. № 1. С. 25–33.
Суходолец В. В. Природа адаптивных эволюционных изменений: приспособленность и экологический потенциал // Генетика. 1998. Т. 34, № 12. С. 1589–1596.
Чиркова Т. В. Физиологические основы устойчивости растений . СПб.: СПбГУ, 2002. 244 с.
Чудинова Л. А., Орлова Н. В. Физиология устойчивости растений: Учеб. пособие к спецкурсу . Пермь: Перм. ун-т, 2006. 124 с.
Alaquori H. A. A., Ozer Genc C., Aricak B., Kuzmina N., Cetin M. The possibility of using Scots pine (Pinus sylvestris L.) as biomonitor in the determination of heavy metal accumulation // Applied Ecology and Environmental Research. 2020. № 2 (29). P. 3713–3727. DOI: 10.15666/aeer/1802_37133727.
Allen C. D., Macalady A. K., Chenchouni H., et al. A global overview of drought and heat induced tree mortality reveals emerging climate change risks for forests // Forest Ecology and Management. 2010. Vol. 259, Iss. 4. P. 660–684. DOI: 10.1016/j.foreco.2009.09.001.
Bak P. Complexity and criticality // How nature works: the science of self-organized criticality. New York, 1996. P. 1–32.
Bates L. S., Waldren R. P., Teare I. D. Rapid determination of free proline for water stress studies // Plant Soil. 1973. Vol. 39, № 1. P. 205–207. DOI: 10.1007/BF00018060.
Brandt L., Levis A. D., Fahey R., Scott L., Darling L., Swanston C. A framework for adapting urban forests to climate change // Environment Science and Policy. 2016. Vol. 66. P. 393–402. DOI: 10.1016/j.envsci.2016.06.005.
Grey G. W., Deneke F. J. Urban Forestry. 2nd Ed. New York: John Wiley and Sons, 1986. 299 p.
Johnstone J. F., Allen G. D., Franklin J. F., Frelich L. E., Harvey B. J., Higuera P. E., Mack M. C., Meentemeryer R. K., Metz M. R., Perry G. L. M., Schoennagel T., Turner M. G. Changing disturbance regimes, ecological memory, and forest resilience // Frontiers in Ecology and the Environmental. 2016. Vol. 14, Iss. 7. P. 369–378. DOI: 10.1002/fee.1311.
Kuznetsova N. F., Semenov M. A., Sautkina M. Yu. Pine forests of East European plain: distribution trends, functions and development problems // Pinus: growth, distribution and uses. New York: Nova Science Publ., 2019. P. 1–47.
Micieta K., Murin G. Three species of genus Pinus suitable as bioindicators of polluted environment // Water, Air & Soil Pollution. 1998. № 104. P. 413–422. DOI: 10.1023/A:1004984121831.
Mikhailova T. A. The physiological condition of pine trees in the Prebaikalia (East Siberia) // Forest Pathology. 2000. Vol. 30. P. 345–359. DOI: 10.1046/j.1439-0329.2000.00221.x.
Popek R., Przybysz A., Gawrońska H., Klamkowski K., Gawroński S. W. Impact of particulate matter accumulation on the photosynthetic apparatus of roadside woody plants growing in the urban conditions // Ecotoxicol. Environ. Safety. 2018. № 163. Р. 56–62. DOI: 10.1016/j.ecoenv.2018.07.051.
Reyer Ch., Brouwers N., Ramming A., Brook B. W., et al. Forest resilience and tipping points at different spatio-temporal scales: approaches and challenges // J. Ecology. 2015. Vol. 103, № 1. P. 5–15. DOI: 10.1111/1365-2745.12337.
Richardson D. M., Rundel P. W., Jackson S. T., Teskey R. O., Aronson J., Bytnerowicz A., Wingfield M. J., Proches S. Human impacts in pine forests: past, present and future // Ann. Rev. Ecology, Evolution and Systematics. 2007. Vol. 38. № 1. P. 275–297. DOI: 10.1146/annurev.ecolsys.38.091206.095650.
Sakugawa H., Cape J. N. Harmful effects of atmospheric nitrous acid on the physiological status of Scots pine trees // Environ. Pollution. 2007. № 3 (147). P. 532–534. DOI: 10.1016/j.envpol.2007.02.012.
Schou E., Thorsen B. J., Jacobsen J. B. Regeneration decision in forestry under climate change related uncertainties and risks: effects of three different aspects of uncertainty // Forest Policy and Economics. 2015. Vol. 50. P. 11–19. DOI: 10.1016/j.forpol.2014.09.006.
Tavoni M., Sohngen B., Bosetti V. Forestry and the carbon market response to stabilize climate // Energy policy. Elsevier. 2007. Vol. 35, № 11. P. 5346–5353. DOI: 10.1016/j.enpol.2006.01.036.
Vacchiano G., Garbarino M., Borgogno Mondino E., Motta R. Evidences of drought stress as a predisposing factor to Scots pine decline in Valle d’Aosta (Italy) // European Journal of Forest Research. 2012. № 131. P. 989–1000. DOI: 10.1007/s10342-011-0570-9.
Yordanov I., Velikova V., Tsonev T. Plant responses to drought, accumulation, and stress tolerance // Photosynthetica. 2000. Vol. 38, Iss. 2. P. 171–186. DOI: 10.1023/A:1007201411474.
Благодарности
Исследования выполнены в рамках госзадания Всероссийского института лесной генетики, селекции и биотехнологии.

 © 2011 - 2026
© 2011 - 2026