



Введение
Представленное сообщение посвящено обобщенному анализу полученных в процессе стационарных и экспедиционных исследований многолетних данных о распространении и численности лесной мышовки на северной периферии ареала. Здесь, на таежном Северо-Западе России, этот вид служит единственным представителем надсемейства тушканчикообразных, обитающим значительно севернее своих более теплолюбивых сородичей. В исследуемой области, куда, помимо Карелии, относится юго-восточная часть Финляндии и Карельский перешеек Ленинградской области, проходит северная граница распространения мышовки, что во многом определяет проявляющиеся здесь особенности ее образа жизни. В этом регионе лесная мышовка распространена только в южной его части до широты г. Беломорска (64º30′), причем в большинстве пунктов наблюдений она немногочисленна. Средний многолетний показатель учетов в южной Карелии – 0.02 экз. на 100 ловушко-суток и 0.72 на 10 канавко-суток. Таким образом, на территории, расположенной у северо-западных границ распространения лесной мышовки, численность этого вида ниже и гораздо менее устойчива, чем в большинстве других частей ареала (Кулик и др., 1968; Шенброт и др., 1995).
Материалы
В основу работы положены исследования, сочетавшие учеты численности зверьков на полевых стационарах (Ладожский, Каскеснаволоцкий, «Кивач») с широкими экспедиционными обследованиями всей территории региона.
Методы
Количественные учеты заключались в расстановке параллельных (на расстоянии 25 м одна от другой) линий давилок (капканчиков Геро) по 25–50 шт. в каждой. Они равномерно распределялись по всем обследуемым биотопам (и модельным участкам) и действовали по 2–4 суток. Приманкой служили кусочки смоченной растительным маслом корочки ржаного хлеба. За показатель обилия принимали число зверьков, попавших за одни сутки работы 100 ловушек (на 100 ловушко-суток), и выраженную в процентах долю данного вида в общем улове ловушками (относительное обилие в населении мелких млекопитающих, или индекс доминирования). Учет и отлов канавками проводился с помощью 30-метровых траншей, имевших по три металлических конуса, сужающихся к горловине и врытых таким образом, чтобы верхний их край находился вровень с дном канавки. Показатель обилия – число зверьков, попавших в конусы за 10 суток работы одной канавки (на 10 канавко-суток) и относительное число зверьков данного вида, выраженное в долях (индекс доминирования, %). При этом общий объем проведенных учетов превышает 360 тыс. ловушко-суток и более 9 тыс. канавко-суток. В итоге с помощью ловушко-линий было добыто 110 мышовок, а канавками – 657.
Для изучения размножения лесных мышовок популяцию этого вида подразделяли на возрастные группы. С этой целью использовался разработанный нами на основе рекомендаций Г. А. Клевезаль (1988) метод возрастной диагностики мышовок (Ивантер, 1972, 1973), заключающийся в подсчетах годовых слоев (линий склеивания) на окрашенных гемотоксилином Эрлиха поперечных срезах нижней челюсти в периостальной зоне кости.
В качестве дополнительных возрастных критериев применялось строение зубов, конфигурация черепа, размерные и весовые показатели, а также размеры тимуса и состояние мехового покрова.
Результаты
Распространение и численность
Лесная мышовка – западный палеарктический вид, более или менее обычный и широко распространенный на Европейском Севере и на Урале. На северо-востоке европейской части России по интразональным местообитаниям проникает до устья реки Печоры и заходит в южные тундры (Колоскова, 1984; Бобрецов, 2004). Распространение этого вида по Уральскому хребту ограничивается на севере Приполярным Уралом (Балахонов, Лобанова, 1986).
Как уже указывалось, в исследуемом регионе лесная мышовка распространена только в южной его половине, при этом в большинстве пунктов наблюдений немногочисленна. В среднем в наших сборах мелких млекопитающих на ее долю приходится 5.9 % (шестое место по численности), но эта величина зависит от способа отлова: при учетах канавками она значительно возрастает (5.2 %), а при учетах ловушками – падает (0.2 %). Средний многолетний показатель учетов в южной Карелии (данные 1948–2017 гг.) составляет 0.02 экз. на 100 ловушко-суток (доля в отловах 0.07 %) и 0.72 на 10 канавко-суток (1.3 %). Таким образом, в Карелии, расположенной у северо-западных границ распространения лесной мышовки, численность этого вида ниже, чем в большинстве других мест ареала (Кулик и др., 1968; Шенброт и др., 1995).
Различается численность мышовки и в пределах изучаемого региона. Так, в северных районах Карелии в отловах ловушками этот вид вообще отсутствует, в центральных показатель его численности составляет 0.01 экз. на 100 ловушко-суток (доля в отловах 0.07 %) и 0.2 на 10 канавко-суток (1.3 %), а в южных – 0.03 (0.9 %) и 0.75 (6.2 %) соответственно. Показатели обилия вида изменяются и в зависимости от ландшафтных условий района и года исследований. Например, в учетах мелких млекопитающих в северо-восточном Приладожье доля лесной мышовки колебалась в сборах канавками от 0 в 1967, 1989 и 2004 гг. до 18.3, 21.4 и 34.3 % в 1980, 2000 и 2002 гг. соответственно. В отловах ловушко-линиями показатель относительной численности мышовок варьировал в том же регионе от 0 до 5.9 %. В Пудожском районе Карелии индекс обилия зверьков менялся при учетах канавками от 0 до 2.7 %, ловушками – от 0 до 0.9 %. В Лахденпохском районе в июне – сентябре 1951 г. С. А. Орлова поймала в ловушки 155 лесных мышовок, что от общего обилия отловленных грызунов составило 21.8 %. Таким образом, по направлению к югу и юго-востоку Карелии плотность популяции лесной мышовки заметно увеличивается.
Биотопическое размещение
В исследуемом регионе лесная мышовка обитает в разных биотопах (табл. 1), но тяготеет к лиственным насаждениям, придерживаясь разреженных участков с богатым травяным ярусом и достаточным количеством убежищ в виде старых пней, куч валежника, упавших стволов и т. д. Не избегает она и хвойных, в особенности сосновых, лесов, но концентрируется здесь по опушкам, долинам ручьев и другим увлажненным участкам с богатым лиственным подлеском и хорошим травостоем. Явное предпочтение, судя по коэффициенту верности биотопу, оказывает этот вид черничным, багульниково-черничным и зеленомошным соснякам, хотя оптимальными для лесных грызунов эти биотопы не назовешь, прежде всего вследствие однообразной и неустойчивой кормовой базы. Правда, это относится в основном к растительным кормовым ресурсам, менее важным для мышовок, предпочитающих животный корм, а он находится здесь в избытке. К тому же представители данного вида почти не сталкиваются в этих местообитаниях с пищевой конкуренцией со стороны других видов мелких млекопитающих, в той или иной мере избегающих биотопы данного типа. При этом мышовки отлавливаются в сосновых лесах не только в годы относительно высокой, но и низкой численности, что нехарактерно для других типов местообитаний.
Свежие вырубки мышовки заселяют неохотно, но со второго – третьего года по мере зарастания лиственными молодняками на лесосеках складываются более благоприятные, хотя и нестабильные защитные и кормовые условия, и эти биотопы становятся для зверьков более привлекательными. По данным наших учетов (Курхинен и др., 2006), численность мышовок на зарастающих вырубках в несколько раз выше, чем в спелых сосняках (0.3 на 10 канавко-суток и 0.07 на 100 ловушко-суток против 0.06 и 0.03 соответственно). При этом индекс доминирования возрастает соответственно в 6 и 2 раза. Как и следовало ожидать, особенно сильное увеличение численности и доли участия в уловах зафиксировано во вторичных лиственных и смешанных лесах.
Открытые биотопы (пашни, окраины сфагновых болот и т. п.) мышовки посещают преимущественно в период расселения молодняка. На полях они ловятся главным образом во время созревания урожая, но иногда живут здесь постоянно, выбирая участки около поросших кустарником и молодыми деревцами межей и каменных гряд. В июне – сентябре 1951 г. С. А. Орлова отловила на овсяных и гороховых полях 125 мышовок из 155 зверьков разных видов.
Таблица 1. Биотопическое размещение лесной мышовки в Карелии (сводные данные за 1959–2017 гг.)
| Биотоп | Число ловушко-суток /канавко-суток | Добыто зверьков | На 100 ловушко- суток / на 10 канавко-суток | % от улова | Коэффициент верности биотопу |
| Учет ловушками | |||||
| Сосняки лишайниковые | 9390 | 4 | 0.04 | 3.42 | +0.33 |
| Сосняки зеленомошные | 52788 | 22 | 0.04 | 1.69 | +0.44 |
| Ельники зеленомошные и травяно-зеленомошные | 104521 | 24 | 0.02 | 0.78 | -0.64 |
| Лиственные и смешанные леса | 96064 | 27 | 0.03 | 0.81 | -0.18 |
| Лиственное мелколесье | 34150 | 2 | 0.01 | 0.15 | -0.73 |
| Зарастающие вырубки | 24494 | 7 | 0.03 | 0.53 | +0.65 |
| Луга и другие сельскохозяйственные угодья | 30916 | 8 | 0.03 | 0.68 | -0.02 |
| Всего | 352323 | 94 | |||
| Учет канавками | |||||
| Сосняки лишайниковые | 110 | 3 | 0.27 | 2.90 | -0.13 |
| Сосняки зеленомошные | 3990 | 311 | 0.78 | 7.77 | +0.05 |
| Ельники зеленомошные и травяно-зеленомошные | 1104 | 41 | 0.37 | 2.70 | -0.5 |
| Лиственные и смешанные леса | 478 | 12 | 0.25 | 1.78 | -0.19 |
| Лиственное мелколесье | 1670 | 56 | 0.34 | 3.34 | -0.05 |
| Зарастающие вырубки | 86 | 1 | 0.12 | 2.00 | -0.36 |
| Луга и другие сельскохозяйственные угодья | 402 | 12 | 0.30 | 2.50 | -0.13 |
| Всего | 7840 | 436 | – | ||
Согласно нашим данным по Карелии (см. табл. 1), к лиственным и смешанным лесам, по крайней мере в условиях северной периферии ареала, мышовка достаточно индифферентна, и это несмотря на богатый травяной покров, разнообразие растительной и животной пищи, обилие валежника и старых пней, что для других видов мелких млекопитающих делает эти формации более привлекательными, чем коренные древостои (Ивантер, Макаров, 2001). Не привлекают мышовок и зеленомошные ельники, хотя именно этот биотоп занимает по численности других мелких млекопитающих одно из первых мест. Половина пойманных в ельниках лесных мышовок оказались сеголетками, тогда как в других типах местообитаний доля последних не превышала 30 %. И если для большинства других представителей Micromammalia зеленомошные и травяно-зеленомошные ельники вполне могут рассматриваться в качестве «стаций переживания» (Ивантер, 1975; Ивантер, Макаров, 2001), то для лесной мышовки они служат скорее стацией расселения молодняка после периода массового размножения вида.
На лугах, пашнях и других сельскохозяйственных угодьях представители данного вида отлавливаются в основном во второй половине лета, во время расселения молодняка, но иногда живут здесь постоянно на участках возле межей и каменных гряд, поросших кустарником и молодыми деревцами.
Тяготение лесных мышовок к зарастающим вырубкам и гарям, лесным опушкам, сенокосным лугам, кустарникам с полянами трав по берегам ручьев и речек, а также к разнообразным лиственным и смешанным лесным насаждениям, где они преимущественно придерживаются разреженных участков с богатым травяным ярусом на влажной, но не слишком сырой почве, с достаточным количеством убежищ в виде старых пней, куч валежника и пр., отмечается по всему ареалу (Заблоцкая, 1957; Попов, 1960; Даргольц, 1964; Кулик и др., 1968; Айрапетьянц, 1970; Ивантер, 1972, 1975; Фокин, 1978; Юдин и др., 1979; Куприянова, 1994; Шенброт и др., 1995; Kubik, 1952 и др.). Именно эти биотопы обеспечивают наилучшие для зверьков кормовые и защитные условия и характеризуются более высокой и устойчивой численностью популяций.
По данным учетов ловушко-линиями, численность мышовки на вырубках примерно в 1.5 раза выше, чем в спелых сосняках, а по учетам канавками – в 3 раза (см. табл. 1). При этом индекс доминирования возрастает соответственно в 2 и 6 раз. Как и следовало ожидать, особенно сильное увеличение численности и доли участия в уловах этого вида зафиксировано во вторичных лиственных и смешанных лесах.
Как показали исследования реакции лесной мышовки на последствия рубок, в частности на лесовосстановление и фрагментацию лесных насаждений, особенно охотно она заселяет увлажненные участки злаковых вырубок, а также хорошо прогреваемые лесосеки на грядах (сельгах) с многочисленными выходами скальных пород, но избегает заболоченных осоково-сфагновых вырубок. Из молодняков 16–20 лет она отдает предпочтение как неоднородным, смешанным насаждениям, так и однородному лиственному и хвойному мелколесью. Гетерогенность биоценоза, появление осветленных участков в результате рубок ухода и собранные в кучи порубочные остатки явно привлекают зверьков.
Активное заселение мышовками вырубок и вторичных лиственных и сосново-лиственных лесов характерно и для других регионов таежной России (Заблоцкая, 1957; Попов, 1960; Реймерс, 1966; Айрапетьянц, 1969). Таким образом, омоложение лесов в результате их интенсивной эксплуатации, как и формирование на вырубках лиственных и смешанных молодняков, способствует росту численности леной мышовки, показатель учета которой в трансформированных рубками лесонасаждениях выше, чем в спелых хвойных лесах.
Суточная и сезонная активность, зимняя спячка
Согласно данным, полученным нами в Карелии, в хорошую погоду мышовки ловятся в любое время суток, но особенно часто в дневные часы. Так, из 34 зверьков, для которых было известно время поимки, 20 (т. е. 58.8 %) было добыто между 9 и 18 ч, 6 (17.6 %) – в утренние и вечерние сумерки и только 8 (23.6 %) – в ночные часы (с 24 до 4 ч). Сравнение этих данных с наблюдениями в других регионах (Снигиревская, 1954; Попов, 1960; Тупикова, 1960; Телегин, 1972; Фокин, 1978; Шенброт и др., 1995) подтверждает вывод Айрапетьянц (1969) о сдвиге активности лесных мышовок в северных частях ареала на светлое время суток как адаптации вида с несовершенной терморегуляцией к суровым северным условиям. Что же касается сезонного изменения ритма суточной активности, о котором пишут Попов (1960) и Айрапетьянц (1969), то на нашем материале он не прослеживается. Преимущественно дневная активность характерна для карельских мышовок не только в начале и конце лета, как в Татарии, Ленинградской области и Западной Сибири, но и в июле.
Иначе ведут себя мышовки в условиях неволи. Здесь, независимо от географической широты места наблюдений, они бывают активны в ночное время суток, с 18–20 и до 1–3 ч ночи. Так, Н. В. Тупикова (1960) пишет, что в конце лета содержащиеся в лаборатории зверьки покидали гнездо примерно с 17 ч (за 2–3 ч до захода солнца, когда в лесу уже нет яркого освещения) и возвращались в него ко второй половине ночи, когда начинала остывать земля. Дневной перерыв в активности составлял 13 ч. Аналогичные сведения приводят П. К. Смирнов (1954), В. А. Попов (1960), В. И. Телегин (1972), А. Э. Айрапетьянц (1969) и др. Наши наблюдения также свидетельствуют о преимущественно ночной активности мышовок, содержащихся в неволе. При этом, как и другие авторы, мы наблюдали тесную связь ритма активности зверьков с температурой. В холодную погоду, при падении температуры воздуха ниже 10 °С, мышовки резко снижают активность, прячутся в гнездо и впадают в своеобразное оцепенение, принимая «позу спячки»: зверек свертывается тугим клубком, уткнув мордочку в брюшко, подогнув лапки и «обвязавшись» хвостом. Потепление вызывает отогревание зверька (при этом он часто зевает и попискивает), и он снова становится деятельным. Изучение ритма суточной активности лесной мышовки в иных экспериментальных условиях (Ердаков и др., 1981) также показало ее монофазный характер при периоде непрерывного покоя, равном 6–12 ч.
На активность лесной мышовки оказывают влияние осадки: при дожде мы ловили в 2–3 раза больше мышовок, чем за «сухие» сутки (Ивантер, 1975). Вместе с тем реакция на выпадение осадков зависит и от температуры. Например, на фоне высоких летних температур стимулирующее влияние осадков проявлялось более отчетливо (за «дождливые» сутки поймано в среднем 1.0 экз., за «сухие» же – всего 0.2). В годы с холодным летом оно было либо мало заметно, либо вовсе отсутствовало, или же осадки оказывали даже отрицательное воздействие: среднесуточный отлов в период затяжных дождей составил 0.8 экз., а в ясную погоду – 0.7.
На зиму лесная мышовка залегает в спячку, которая длится несколько месяцев. Самый поздний случай ее поимки зафиксирован в Карелии 4 октября 1951 г., самый ранний – 7 мая 1999 г. Однако в массе эти зверьки попадают в ловушки со второй декады мая по конец сентября. Следовательно, в зимней спячке лесные мышовки проводят у нас 7–7.5 мес., т. е. столько же, сколько и в других регионах (Снигиревская, 1954; Попов, 1960; Юрлов и др., 1965; Кулик и др., 1968; Айрапетьянц, 1969; Фокин, 1978; Шенброт и др., 1995). Сроки пробуждения и начала спячки зависят от погодных условий. В годы с ранней и теплой весной мышовки пробуждаются раньше, а поздняя, теплая и сухая осень может задержать наступление спячки почти на месяц. Обычно начало спячки совпадает с понижением температуры воздуха до +6°...+10°. Зверьки к этому времени сильно жиреют, удваивая или даже утраивая свою массу. Как показали эксперименты в лаборатории (Айрапетьянц, 1969), в неволе успешнее перезимовывают мышовки, устраивавшиеся на зимовку на глубине 40–50 см. Именно поэтому в условиях севера свои зимние гнезда они предпочитают помещать не в дуплах и пустотах за корой старых пней, а под землей.
Пробуждение лесных мышовок от зимнего сна происходит обычно после окончательного схода снега: у нас это обычно бывает в конце апреля – начале мая. Однако конкретные сроки их пробуждения зависят, как уже упоминалось, от условий года. В Карелии и других районах таежного Северо-Запада, по средним многолетним данным, лесные мышовки становятся обычными приблизительно с середины мая, а в средней полосе России – с конца апреля.
Питание
Как показывает анализ содержимого желудков добытых зверьков (табл. 2), основу питания лесной мышовки в Карелии составляют насекомые (в среднем 94.3 % встреч), на втором месте паукообразные (65.2 %), на третьем – растительные корма (31.2 %). Насекомые поедаются на различных стадиях развития, соотношение их изменяется по годам и сезонам, но в целом в питании преобладают личинки (79 % против 63 % имаго), представленные в основном листоедами. Куколки и яйца насекомых в бесснежный период потребляются редко. В пищевой рацион вида входят также моллюски и дождевые черви (1.4 и 3.5 % соответственно), но по количеству и частоте потребления эти корма значительно уступают основным и не относятся к массовым пищевым объектам лесной мышовки.
Что же касается насекомых, то чаще других потребляются гусеницы бабочек (обнаружены в 70 % исследованных желудков) и жесткокрылые (50 %), причем последние поедаются преимущественно на стадии имаго (49 % против 5 % личинок), а также равнокрылые (11 %), двукрылые (7 %) и перепончатокрылые (9 %), в основном личинки пилильщиков. При этом, в отличие от землероек, поедающих в основном напочвенных насекомых (Ивантер, Макаров, 2001), в рационе мышовок преобладают жуки, живущие на растительности (долгоносики, щелкуны и листоеды), а также обитатели цветов – трипсы (из всех насекомоядных зверьков их поедают только мышовки). Из паукообразных наибольшее кормовое значение для мышовок имеют сенокосцы (41 %), мелкие виды пауков (35 %) и лесные клещики (7 %).
Растительные корма, представленные зелеными частями растений (36 %), семенами (43 %) и ягодами (9 %), хотя и несколько уступают по частоте потребления животному корму (43 % против 100 %), но количественно заметно превосходят его, нередко составляя более половины всего содержимого желудков.
При общем сходстве питания лесных мышовок из разных районов, местообитаний и по годам некоторый отпечаток на их рацион накладывает кормовой спектр биоценоза и урожай кормов. Например, в питании лесных мышовок, отловленных в Карелии в 1951 г. преимущественно на овсяных и гороховых полях и клеверище, преобладали семена и зеленая масса, а в годы урожая ягод черники (1957, 1958, 1961) – именно этот корм. Наличие характерных годовых различий в питании лесной мышовки в Карелии иллюстрирует и рисунок.
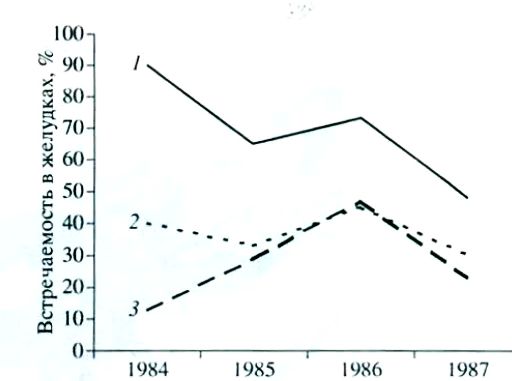
Встречаемость личинок чешуекрылых (1), пауков (2) и сенокосцев (3) в желудках лесных мышовок по годам
Frequency of lepidoptera larvae (1), spiders (2) and harvestmen (3) in the stomachs of birch mice by years
Как показали наблюдения за подопытными зверьками (Ивантер, 1975), в неволе мышовки особенно охотно поедали листья мокреца, семена подсолнуха, клевера, салата, ячменя, овса, ели, сосны, ягоды черники, земляники, малины, гусениц, куколок муравьев и мух. К этому списку предпочитаемых кормов, по наблюдениям за содержащимися в клетках лесными мышовками, отловленными в Ленинградской области (Айрапетьянц, 1970), можно добавить мякоть плодов дикой яблони, семена ржи и пшеницы. Хорошо поедали они плоды шиповника, ягоды костяники, семена лугового и горного клевера, семена и стебли лесного вейника, цветы одуванчика и подземные части купальницы. 23 вида из предлагаемых им растений лесные мышовки использовали слабо: листья, почки и побеги ивы, березы, серой ольхи, семена и хвою сосны и ели, плоды, листья и стебли борщевика, одуванчика, латука, стебли и семена ежи сборной, овсяницы луговой, тимофеевки и др. Перед спячкой зверьки выбирают наиболее калорийные части растений. Из беспозвоночных они охотно поедали прямокрылых, ос, шмелей, куколок лесных муравьев и пауков, отдавая им предпочтение перед ночными бабочками и различными жуками. При этом живой корм мышовки явно предпочитают и при возможности выбора пищи начинают именно с него. Суточный рацион зверьков состоял из 1.5–2 г семенного корма (подсолнуха, зерен злаков), 0.5 г ягод или плодов, 3–4 г воды или молока и живого корма: 7–8 мучных червей, 40–50 муравьиных яиц, 1–2 кузнечиков и т. д. Наблюдения Айрапетьянц (1970) и Фокина (1978) выявили и значительную потребность мышовок в воде. В природе они, возможно, могут довольствоваться и росой, в неволе же охотно и регулярно пьют воду или молоко, а лишенные жидкости и при отсутствии сочного корма погибают через сутки.
По данным сибирского зоолога В. И. Телегина (1972), в мае на долю животной пищи приходится 67 % поедаемого мышовкой пищевого рациона, в июне – июле потребление животного корма снижается, а растительного, наоборот, увеличивается, и в августе на его долю приходится уже 86 %. Отмеченная В. И. Телегиным сезонная изменчивость в питании лесных мышовок в основном обусловлена доступностью того или иного вида корма. В конце лета и осенью созревают семена многих растений, которые не только легко доступны, но и весьма калорийны, обеспечивая мышовке быстрое накопление жировых запасов для зимовки.
Таблица 2. Состав пищи лесных мышовок (встречаемость, % от числа исследованных желудков)
| Вид корма | 1951 г. | 1959–1968 гг. | 1969–1971 гг. | 1984–1987 гг. |
| Насекомые | 78.9 | 50.0 | 75.0 | 94.3 |
| имаго | 63.1 | |||
| личинки | 79.4 | |||
| яйца | – | 2.5 | 6.2 | 0.7 |
| Отряд не определен | 63.1 | 7.5 | 25.0 | |
| Жесткокрылые | – | 25.0 | 50.0 | 49.6 |
| имаго | 48.9 | |||
| личинки | 5.0 | |||
| Двукрылые | 7.1 | |||
| имаго | 6.4 | |||
| личинки | 1.4 | |||
| Перепончатокрылые | 8.5 | |||
| имаго | 2.8 | |||
| личинки | 5.7 | |||
| Равнокрылые | 11.3 | |||
| Чешуекрылые (личинки) Прямокрылые | 2.5 | 68.8 | ||
| Клопы | 5.7 | |||
| Трипсы | 2.1 | |||
| Тараканы | – | 2.5 | 6.2 | |
| Паукообразные | 65.2 | |||
| пауки | 34.8 | |||
| клещи | 7.1 | |||
| сенокосцы | 41.1 | |||
| Дождевые черви | 10.5 | 7.5 | 12.5 | — |
| Моллюски | – | 2.5 | 6.2 | 0.7 |
| Растительные остатки | 73.7 | 75.5 | 37.5 | 31.2 |
| Зеленая масса | 21.0 | 47.5 | 25.0 | |
| Семена | 68.4 | 40.0 | 18.7 | |
| Ягоды | – | 15.0 | 6.2 | |
| Число исследованных желудков | 19 | 40 | 16 | 142 |
Заключение
Отмеченные выше экологические особенности, свойственные лесной мышовке в специфических условиях северо-западной периферии ареала, отличаются значительной лабильностью и гармоничной согласованностью как с плотностью и состоянием популяции, так и с внешними факторами среды. Разнообразие экологических приспособлений, их трансформация и смена во времени и пространстве, адаптивный динамизм и ориентированность на конкретную экологическую ситуацию, складывающуюся из взаимодействия экзогенных и эндогенных факторов, обуславливают относительную стабильность и оптимальное состояние популяций рассматриваемого вида в экстремальных для него приграничных условиях. Вместо узких специальных приспособлений индивидуального ранга в периферических его населениях успешно реализуются широкие структурно-популяционные адаптивные комплексы, отличающиеся динамичностью и высокой скоростью компенсаторной перестройки. Все это вплотную приближает нас к разработке общей концепции периферических популяций мегаареальных политипических видов животных. Проведенный нами анализ закономерных географических отличий, характеризующих популяционную специфику репродукции зверьков этого вида, обитающих на северных пределах области распространения, подтвердил выдвинутое нами ранее положение, согласно которому в экологическом центре (оптимуме) видового ареала население вида благодаря функционированию внутрипопуляционных компенсаторных механизмов отличается не только относительно высоким уровнем численности, но и большей устойчивостью, тогда как у северных границ распространения эти стабилизирующие механизмы практически отсутствуют, и организация населения изменяется в широком диапазоне и с большей амплитудой. Соответственно более выражены и гораздо рельефнее проявляются близ северных границ ареала и специфические структурно-популяционные адаптации, направленные на максимально эффективное воспроизводство видового населения, способное обеспечить виду и его популяциям необходимое преодоление экстремальных условий экологического пессимума и в конечном счете содействовать как более широкой их экспансии за границы ареала, так и успешному закреплению в новых областях.
Библиография
Айрапетьянц А. Э. Насекомоядные и грызуны Ленинградской области // Звери Ленинградской области. Л.: Изд-во ЛГУ, 1970. С. 47–165.
Балахонов В. С., Лобанова Н. А. Лесная мышовка в антропогенных биотопах Полярного Урала // Горные экосистемы Урала и проблемы рационального природопользования. Информ. материалы ИЭРиЖ УНЦ АН СССР. Свердловск, 1986.
Бобрецов А. В. Насекомоядные. Мышевидные грызуны // Млекопитающие Печоро-Илычского заповедника. Сыктывкар: Коми книжн. изд-во, 2004. С. 38–109, 206–301.
Даргольц В. Г. Особенности динамики численности лесной мышовки в подзоне южной тайги // Современные проблемы изучения динамики численности популяций животных. М.: Наука, 1964. С. 76–84.
Ердаков А. Н. Организация ритмов активности грызунов . Новосибирск: Наука, 1984. 181 с.
Заблоцкая Л. В. Материалы по экологии основных видов мышевидных грызунов Приокско-Террасного заповедника и смежных территорий // Труды Приокско-Террасного заповедника. 1957. Вып. 1. С. 170–240.
Ивантер Э. В. К экологии лесной мышовки (Sicista betulina Pall.) // Aquiloo. Ser. Zool. 1972. Vol. 13. P. 103–108.
Ивантер Э. В. Методика определения возраста лесной мышовки Sicista betulina (Rodentia, Dipodoidea) // Зоологический журнал. 1973. Т. 52. Вып. 2. С. 255–257.
Ивантер Э. В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР. Л.: Наука, 1975. 246 с.
Ивантер Э. В., Макаров А. М. Территориальная экология землероек-бурозубок (Insectivora, Sorex) . Петрозаводск: Изд-во ПетрГУ, 2001. 272 с.
Клевезаль Г. А. Определение возраста млекопитающих . М.: Наука, 1967. 142 с.
Клевезаль Г. А. Регистрирующие структуры млекопитающих в зоологических исследованиях . М.: Наука, 1988. 285 с.
Колоскова Н. И. О северной границе ареала лесной мышовки Sicista betulina (Rodentia, Dipodidae) в Европейской части СССР // Зоологический журнал. 1984. Т. 63. Вып. 5. С. 784–788.
Кулик Н. Л., Тупикова Н. В., Никитина Н. А., Карасева Е. В., Суворова Л. Г. Материалы по экологии лесной мышовки (Sicista betulina Pallas) // Cборник трудов Зоологического музея МГУ. 1968. Т. 10. С. 146–159.
Куприянова И. Ф. Мышевидные грызуны // Фауна европейского Северо-Востока России. Млекопитающие. СПб.: Наука, 1994. Т. 2. Ч. 1. С. 68–99.
Курхинен Ю. П., Данилов П. И., Ивантер Э. В. Млекопитающие Восточной Фенноскандии в условиях антропогенной трансформации таежных экосистем . М.: Наука, 2006. 208 с.
Попов В. А. Млекопитающие Волжско-Камского края . Казань, 1960. 466 с.
Реймерс Н. Ф. Птицы и млекопитающие Южной тайги Средней Сибири . М.; Л.: Наука, 1966. 419 с.
Смирнов П. К. Наблюдения по экологии грызунов Ленинградской области // Ученые записки ЛГУ. 1954. Вып. 181 (Биол. 38). С. 45–129.
Снигиревская Е. М. Экология и хозяйственное значение мышевидных грызунов в широколиственных лесах Жигулевской возвышенности : Автореф. дис. … канд. биол. наук. Л., 1954. 24 с.
Телегин В. И. Териофауна лесопарка Новосибирского научного центра // Вопросы лесопаркового хозяйства и озеленения Новосибирского научного центра. Новосибирск, 1972. С. 24–41.
Тупикова Н. В. Суточный ритм активнoсти лесной мышовки // Зоологический журнал. 1960. Т. 39. Вып. 6. С. 561–572.
Фокин И. М. Тушканчики. Жизнь наших птиц и зверей . Л.: Изд-во Ленингр. ун-та, 1978. Вып. 2. 184 с.
Шенброт Г. И., Соколов В. Е., Гептнер В. Г., Ковальская Ю. М. Млекопитающие России и сопредельных регионов. Тушканчикообразные . М.: Наука, 1995. 576 с.
Юдин Б. С., Галкина Л. И., Потапкина А. Ф. Млекопитающие Алтае-Саянской горной страны . Новосибирск: Наука. Сиб. отд-ние, 1979. 293 с.
Юрлов К. Т., Юдин Б. С., Потапкина А. Ф. и др. К характеристике фауны мелких млекопитающих северной степи Барабинской низменности // Животный мир Барбы. Новосибирск: Наука, 1965. С. 184–207.
Kubik I. Badania nad morrfologia I bioljgia smuzki (Sicista betulina Pall.) z Bialoweiskirgo Parku Narodowego // Ann. Univ. M. Cure-Skilod., Lublin. 1952. Vol. 11. № 1. S. 47–61.

 © 2011 - 2026
© 2011 - 2026