



Введение
Зарянка является многочисленным и популярным видом в Европе для исследования миграции (Cramp, 1992; Gyimóthy et al., 2011). Она относится к группе видов, летящих на средние дистанции (Цвей, 2008), пролет которых осуществляется ночью (Зимин, Носков, 2020). Известно, что события, происходящие в этот период, оказывают существенное влияние на будущий успех размножения птиц (Newton, 2006). Иногда уровень смертности во время пролета на порядок выше, чем во время зимовок и размножения (Sillett & Holmes, 2002). Установлено, что миграционная стратегия зарянок в различных частях гнездового ареала отличается. Если в южных регионах Европы этот вид является мигрантом на короткие дистанции (Remisiewicz et al., 1997), который затрачивает минимальное время для пролета, то в северных широтах, напротив, это типичный мигрант на средние дистанции, требующий определенных затрат энергии для броска (Gyimóthy et al., 2011). На территории европейской части России стратегия миграции зарянки достаточно хорошо изучена на Куршской косе (Цвей, 2008), где рассмотрены вопросы миграционной экологии, начиная от фенологии и заканчивая скоростью жиронакопления. Показано, что зарянки мигрируют с небольшими скоростями, ненадолго задерживаясь на остановках всего 1–2 дня. Незначительные жировые накопления приводят к коротким миграционным броскам, дающим птицам возможность пролететь около 6 часов (Цвей, 2008). Изменение жирности в течение сезона у зарянок имеет волнообразную природу, при этом в пик пролета мигрируют птицы с наивысшей степенью жирности (Блюменталь, 1967). Характер такого изменения в период миграции связан с пролетной активностью. Средняя продолжительность остановки и факторы, определяющие начало миграционного броска молодых зарянок, указаны в работе В. Булюка и А. Цвея (Buluk, Tsvey, 2006). Ими был определен механизм феномена ночных мигрантов, которые по-разному принимают решение о начале миграционного броска. Основным фактором, определяющим решение о длительности остановки и начале ночного полета, являются индивидуальные эндогенные циркадные ритмы, связанные с окружающей средой. Имеются данные о летних миграциях зарянок в Ленинградской области (Резвый, Савинич, 2005; Резвый, 2011) и Карелии (Зимин, 2012).
Однако анализ стратегии поведения молодых зарянок на миграционной остановке исследован недостаточно. Имеются данные по моделям осенних миграций близкородственного зарянкам вида варакушки. Показаны достоверные различия в выбираемых стратегиях поведения молодых и взрослых особей на миграционных остановках в восточной Фенноскандии (Панов, Чернецов, 2010а, б). Установлено, что взрослые птицы при достаточно коротких остановках успевают значительно увеличивать свои энергетические резервы в отличие от молодых варакушек. Предполагается, что молодые особи имеют более низкую скорость жиронакопления и делают продолжительные остановки чаще, чем взрослые. Это может быть связано с ландшафтной обстановкой, которая усиливает возрастные различия в миграционных стратегиях (Панов, Чернецов, 2010а). Поэтому нами было выдвинуто предположение о том, что модель осенних миграций молодых зарянок будет подобна таковой варакушек.
Цель данной работы – изучить особенности миграционной стратегии зарянок-первогодок в условиях речных долин средней тайги в популяции из восточной части Русской равнины.
Материалы
Исследования проведены в долине среднего течения р. Сысола (Республика Коми), которая определена как ключевая орнитологическая территория международного значения для многих гнездящихся и мигрирующих птиц (Ануфриев, Кочанов, 2000). Приблизительно 50 видов воробьинообразных пролетает осенью через эту территорию (Накул, 2018). Бассейн реки расположен в северо-восточной части Восточно-Европейской равнины, примерно в 450 км к западу от Северного Урала в подзоне средней тайги. Река протекает с юга на север и является одним из главных притоков р. Вычегда, которая входит в речной бассейн Северной Двины. Птиц отлавливали в с. Межадор (61°08 с. ш. 50°19 в. д). Территория отловов характеризуется высоким разнообразием биотопов: сенокосные луга и ивовые заросли граничат с крапивными пустошами, зарослями борщевика Сосновского, частными картофельными огородами, зарослями плодово-ягодных кустарников и деревьями (береза, черемуха, сосна обыкновенная), как отдельно стоящими, так и растущими группами. Эти условия являются благоприятными как для насекомоядных птиц, так и для видов, питающихся ягодами и семенами различных культурных и диких растений.
Материал собран в августе – сентябре 2015–2017 и в сентябре 2019 года. Всего было проанализировано 188 особей молодых зарянок. Из них переловлено в течение сезона 17 птиц, что составило 9 % от общего объема отловленных зарянок. Отлов проводили стандартными паутинными сетями 5–12 м, общая протяженность которых составила 120 м. Сети расставляли в местах массовых скоплений и кормления птиц. Проверка сетей проходила ежедневно в светлое время суток с интервалом в 1–2 часа. Ночью птицы в сети не попадались. В 2017 и 2019 гг. для привлечения зарянок применяли «звуковую ловушку» (Панов, Чернецов, 2010б) – акустическую систему Supra PAS-6255, которая использовалась в ночное время (с 22:00 до 4:00 следующего дня). Запись голоса зарянки была представлена видовой песнью и позывками из фондов Фонотеки голосов животных им. проф. Б. Н. Вепринцева (Вепринцев и др., 2007). Определение возраста осуществляли согласно методике, предложенной Н. В. Виноградовой с соавторами (1976). Стадии линьки описывали по схеме, принятой для зарянки (Савинич, 1990). Массу тела измеряли электронными весами с точностью до 0.01 г. Количество жировых запасов определяли визуально по количеству подкожных жировых отложений на межключичной и брюшной артериях, где расположены основные депо подкожного жира (Блюменталь, 1967): 1 балл – «нежирные» птицы, 2 балла – «маложирные», 3 балла – «среднежирные», 4 балла – «жирные». Для расчетов продолжительности миграционной остановки зарянок использованы данные пойманных птиц, начиная с первого пика повышения численности вида в отловах. Для исключения местных зарянок-первогодок из выборок для анализа скорости жиронакопления использованы данные особей, которые впервые были отловлены ближе к медианной дате и имели стадию линьки не ниже четвертой.
Методы
Для каждого параметра (длина крыла, начальная и конечная масса тела, начальная и конечная жирность) рассчитывали среднее и стандартную ошибку. Для оценки значимости различий между выборками использовали непараметрический критерий Манна – Уитни (значение z) при p < 0.05. «Тощая» масса тела рассчитывалась согласно уравнению: Lbm = 2.248178 + 0.172748 × w, где Lbm – «тощая» масса тела, w – длина крыла в мм (Цвей, 2008), а определение продолжительности миграционного полета – согласно предложенной методике исходя из массы их энергетических резервов (Цвей, 2008). За 1 г энергетических резервов принималось 21.6 кДж энергии (Klaassen et al., 2000), а цена полета в неподвижном воздухе составляет 7 BMR (уровней базального метаболизма) (Klaassen et al., 2000). BMR рассчитывался на основании зависимостей, приведенных в работе В. Р. Дольника (1995). Длительность миграционной остановки проведена на основе средних минимальных промежутков между повторными отловами (разницу между датами первого и последнего отловов). Моделирование изменения массы тела и степени жирности у мигрантов на остановках оценивали методом множественной пошаговой регрессии. В исходную модель были включены начальная масса тела, дата, посуточное изменение массы тела птицы, а также изменение массы тела в течение суток (Schaub, Jenni, 2000). Корреляционные связи по Спирмену установлены для непараметрических данных (включенные факторы в модель) обоих выборок. В выборку по расчету изменения массы и показателей жирности через одни сутки входили птицы, которых перелавливали на следующий день после начального отлова. Для анализа второй выборки птиц отбирались данные повторных отловов тех особей, которые попадали в сети повторно более чем через сутки после начального отлова. В расчет не вошли данные по изменению массы особей, которые были отловлены в первый день вечером и на следующий день ранним утром, чтобы избежать заниженных результатов ночных потерь масс тела птицы. Для расчета всех упомянутых коэффициентов использовали программные пакеты STATUSTICA 6.0, PAST 3.13 и Excel 2010.
Результаты
Динамика пролета и продолжительность остановки. Динамика и сроки пролета в разные годы исследований отличались незначительно (табл. 1). Продолжительность миграционной остановки в среднем для данного вида составила 3.04 ± 0.57 дня (n = 23). Наибольший интервал между первым и вторым отловом, отмеченный во время первой и второй волн миграции в I и II декадах сентября, у некоторых особей составил 9 дней.
Таблица 1. Динамика осенней миграции молодых зарянок
| Год исследования | Сроки миграционных волн | Медиана пролета | ||
| 1-я волна | 2-я волна | 3-я волна | ||
| 2015 | 7.09–10.09 | 15.09–17.09 | 23.09–29.09 | 15.09 |
| 2016 | 30.08–1.09 | 27.09–28.09 | – | 16.09 |
| 2017 | 9.09–10.09 | 13.09–15.09 | 20.09–22.09 | 14.09 |
| 2019 | 8.09–9.09 | 11.09–13.09 | 24.09–25.09 | 12.09 |
Примечание. Прочерк означает отсутствие миграционной волны (массового пролета в этот период).
Совмещение линьки и миграции. В общей выборке преобладали птицы на V стадии линьки, на долю которых приходилось до 50 %. В первую миграционную волну доминировали птицы на IV–V стадиях линьки, во вторую и третью волны – на последних стадиях постювенильной линьки и полностью перелинявшие особи (92 %).
Таблица 2. Распределение молодых зарянок в отловах по стадиям линьки
| Период миграции | Стадии линьки и доля птиц в отловах | |||||
| I | II | III | IV | V | Полная | |
| 1-я волна | 0.061 | 0.182 | 0.030 | – | 0.364 | 0.364 |
| 2-я волна | – | 0.095 | 0.079 | 0.079 | 0.492 | 0.254 |
| 3-я волна | – | 0.040 | – | 0.240 | 0.340 | 0.380 |
Примечание. Прочерк означает отсутствие птиц на данной стадии линьки в отловах.
Скорость жиронакопления. Для птиц, переловленных через сутки, отмечена корреляционная зависимость между их конечной и исходной массой тела (r = 0.76 при р < 0.01), а также конечной массой тела и скоростью жиронакопления (r = 0.79 при р < 0.04). Для зарянок, интервал переловов которых был более суток, достоверных корреляций не обнаружено. У особей, оставшихся на остановке дольше двух суток, отмечено более высокое накопление массы перед отлетом по сравнению с группой птиц, прибывших на остановку в начале сезона миграции (рис. 1А). При длительных остановках успешное увеличение массы тела у зарянок проходило в начале миграционного сезона, а в конце выявлена тенденция к уменьшению этого показателя (рис. 1В).
После первых и вторых суток пребывания зарянок на остановке отмечены максимальные потери их массы тела (около 8 % от среднего значения массы тела зарянок), в то время как на шестые сутки после первичного отлова, наоборот, максимальный прирост (около 11 % от среднего значения массы тела). Несмотря на энергетические потери в первые сутки, общий тренд изменения массы тела молодых зарянок оставался положительным (рис. 2). Средний прирост в весе тела при средней длительности остановки в 3.04 ± 0.57 дня составил около 0.31 г в сутки. Расчетная модель скорости жиронакопления у зарянок на миграционной остановке показала положительную динамику увеличения веса тела к моменту отлета (рис. 3). В течение суток масса тела имела значительные колебания, особенно в середине светлого времени суток, но в целом общий тренд динамики был положительным к концу дня (рис. 4).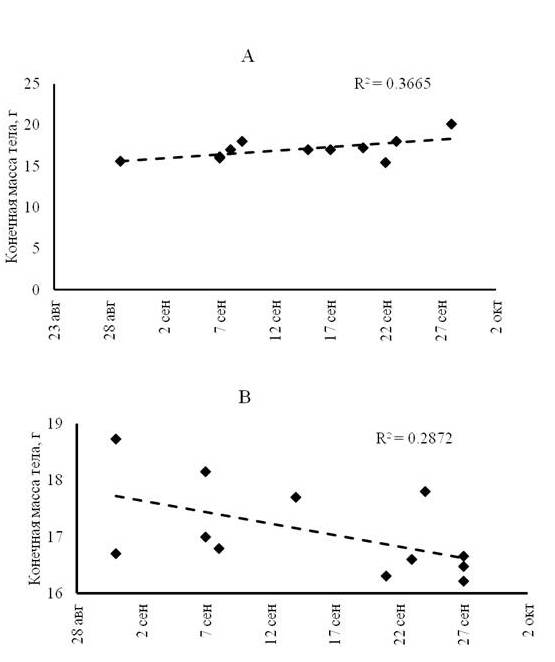
Рис. 1. Конечная масса тела мигрирующих зарянок, остановившихся на сутки (А) и на длительный срок (B)
Fig. 1. The final body weight of migrating Robins stopped for a day (A) and for a long period (B)
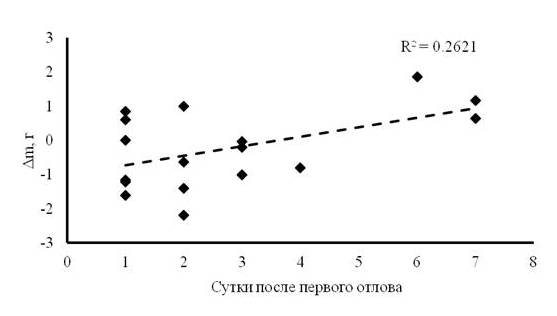
Рис. 2. Динамика отношения начальной и конечной массы тела зарянок на миграционной остановке (n = 17), где Δm – разница между первым и последним взвешиванием тела птицы
Fig. 2. The dynamics of the ratio of the initial and final body mass of Robins at the migration stopover site (n = 17), where Δm is the difference between the first and last weighing of the bird's body
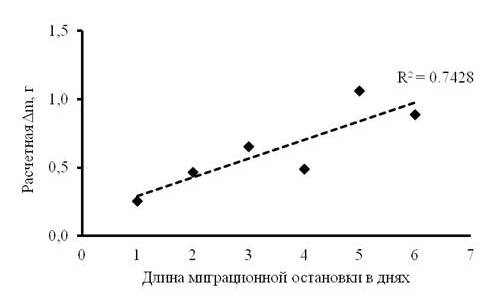
Рис. 3. Расчетная модель жиронакопления зарянок в течение миграционной остановки, где Δm – расчетная масса тела молодой зарянки, накопленная за период остановки
Fig. 3. The calculated model of fat accumulation of Robins during migration stopover, where Δm is the estimated body mass of a young Robin accumulated during the stopover period
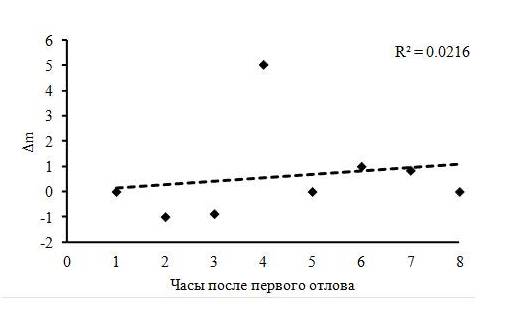
Рис. 4. Суточные изменения массы тела у молодых зарянок
Fig. 4. Daily change of body weight in young Robins
Степень жировых резервов у птиц в общей выборке варьировала, но подавляющее число птиц (67 %) оказались «нежирными», доля «жирных» особей составила 4 %. В начальных отловах в обеих выборках преобладали «тощие» птицы (81 и 92 % соответственно). Уровень жира во всех группах на миграционной остановке не изменялся. Индивидуальные наблюдения за отдельными птицами показали, что жирность не изменялась и оставалась на прежнем уровне через сутки, а также у переловленных на третьи, пятые и восьмые сутки относительно номинального уровня. В то же время на вторые и девятые сутки отмечено снижение, а на четвертые и шестые сутки, наоборот, увеличение уровня показателя жирности на один балл. Однако при сравнении выборок достоверных изменений жирности птиц не обнаружено. Общая средняя разность между начальным и конечным уровнем жировых резервов оказалась ничтожно малой – 0.08 ± 0.42 балла.
Масса энергетических резервов (разница между фактической массой тела и «тощей» массой) транзитных зарянок в средней тайге востока Русской равнины составила от -0.2 до 5.8 г (2.17 ± 0.10 г в среднем) и была меньше, чем у птиц, которые совершали остановку (табл. 3). Однако различия в энергетических резервах между транзитными особями и зарянками, оставшимися на остановке, незначимы (z = -1.4; p = 0.15). При последнем измерении перед отлетом показатель энергетических резервов в среднем уменьшался до 2.37 ± 0.33 г, что в пересчете на длительность беспересадочного полета составило 5.8 ± 0.74 часа.
Таблица 3. Сравнительная характеристика энергетических резервов у мигрирующих молодых зарянок
| Группа | «Тощая» масса в среднем | Энергетические резервы | Часы полеты |
| Транзитные | 14.51 ± 0.03 | 2.17 ± 0.10 | 5.3 ± 0.2 |
| Совершившие остановку | 14.48 ± 0.09 | 2.6 ± 0.26 | 6.4 ± 0.58 |
Обсуждение
Мигрирующих птиц условно можно разделить на две группы. Транзитная группа включает в себя особей, которые не задерживаются на остановке. Вторая группа молодых зярянок остается на более длительный временной срок, их доля в общем потоке не более 10 %. Такое соотношение не является чем-то особенным и встречается у многих других воробьиных видов птиц (Chernetsov et al., 2007; Salewski et al., 2007; Панов, Чернецов, 2010а; Чернецов, 2010). Наши исследования показали, что динамика отлова зарянок дает возможность четко отличить местных птиц от мигрирующих. Особенно хорошо это прослеживается при отловах с помощью аудиоловушек. Местные молодые птицы попадают в сети в последней декаде августа, после чего наступает период отсутствия в отловах новых птиц, несмотря на привлечение видовыми песнями. С начала сентября число пролетающих новых птиц резко возрастает, вследствие чего пролет проходит несколькими волнами до октября (см. табл. 1). Такая схема разделения потоков отмечалась ранее в популяциях зарянки на западе России и Европы (Цвей, 2008). Кроме того, подобная модель выявлена и у других видов воробьинообразных (Панов, Чернецов, 2010а; Kanja, 1981), что указывает на широкое применение этой стратегии внутри отряда. Полученные данные по совмещению линьки и миграции у молодых зарянок свидетельствуют о преобладании в первой волне миграции особей из ближних гнездовых территорий. В последующих волнах пролета по численности преобладают поздние мигранты, уже получившие опыт пролета. Схожая стратегия отмечена для варакушки, родственного зарянкам вида, в Карелии и в нижнем Приобье, где на первых этапах подавляющее число особей находилось на завершающих стадиях линьки (Блюменталь, 1967; Рыжановский, 1988, 2014; Панов, Чернецов, 2010а).
В целом длительность пребывания нетранзитных особей в местах остановки может быть связана с разными факторами. Ряд авторов отмечает, что на длительность остановки влияет качество местообитаний, протяженность предстоящего экологического барьера (Lindström, 2003; Ktitorov et al., 2010; Bayly et al., 2019), а также социальная «напряженность» или конкуренция (Блюменталь, 1967) на миграционных остановках. На исследуемой территории вопрос факторов, определяющих длительность пребывания птиц на остановках, остается открытым. Наличие значительных экологических барьеров в этой части Русской равнины не просматривается, особенно в связи с учетом развитых речных систем Печорского, Северодвинского и Мезенского бассейнов. Также кажется сомнительным предположение о бедности кормом местных биотопов, учитывая их мозаичность и разнообразие экологических ниш. Определение конкуренции между остановившимися особями в рамках этого исследования не проводились. А влияние погодных факторов на длительность остановки молодых зарянок не подтвердилось (Buluk, Tsvey, 2006).
Известно, что перед первым миграционным броском зарянки накапливают значительные жировые резервы (Блюменталь, 1967; Рыжановский, 2014). Наши результаты показали незначительное увеличение веса тела птиц во время остановки перед миграционным броском (см. рис. 1, 2). Зарянки, как правило, следуют без значительных накоплений жира, преобладающее большинство птиц имеют массу тела, близкую к теоретической «тощей». В этой группе особи обладают достаточным энергетическим резервом для дальнейшей миграции, но не более чем на одну ночь (в среднем на 5–6 часов, а максимальный запас – на 12 часов беспрерывного полета). Для мигрирующих через Куршскую косу зарянок данный показатель чуть меньше и равен 3.7 часа (Цвей, 2008). Особи, решившие остаться на более длительный срок, значительно теряют в весе в первые сутки. Восполнение потраченной энергии осуществляется на третьи сутки, а к завершению остановки потерянный уровень массы полностью восстанавливается и, реже, увеличивается на 1 % относительно средней «тощей» массы. Очень похожие результаты были получены на молодых варакушках (Ellegreen, 1991; Панов, Чернецов, 2010а). Одной из причин низкой скорости жиронакопления в первые сутки может быть ориентация в незнакомой местности (оценка биотопа, где остановилась птица) и поиск кормных мест, что отнимает много времени и энергии у прилетевших птиц (Чернецов, 2010). Другой причиной служит появление ночных заморозков, увеличение количества дней с осадками, снижение видового разнообразия и обилия корма. Так, наши данные свидетельствуют о том, что пик интенсивности питания приходится на середину дня и в предвечерние часы перед миграционным броском (см. рис. 4). Они подтверждают, что за среднюю миграционную остановку молодые зарянки способны как восполнить потери, так и преувеличить их. Однако в период ночевок на снижение массы могут повлиять низкие температуры воздуха, достигающие минусовых значений и способствующие увеличению терморегуляционных потерь. Следовательно, птицы после ночевки пытаются восполнить энергетические потери, которые могут достигать до 8 % от «тощей массы» тела, что согласуется с данными по взрослым зарянкам из западных регионов России (Zimin, 2003) и Европы (Alerstam and Lindstrom, 1990). В результате влияние всех этих факторов снижает скорость накопления массы тела птиц, в итоге жировые накопления меняются незначительно. Стоит также отметить, что птицы, прилетающие позднее и совершающие длительные остановки, успевают накопить жира меньше, чем ранние мигранты. Это, на наш взгляд, связано с тем, что в общих отловах в начале сезона миграции преобладают особи, подлетевшие из соседних районов и их уровень жира еще не потрачен на миграционные броски. Также причиной низких скоростей накопления жира в завершении сезона миграции может служить обеднение кормовой базы биотопов на месте остановки. Исследования экологии массовых грибных комаров и мокрецов в таежной зоне показали снижение летной активности и численности значительного количества этих видов к концу сентября (Глухова, 1989; Субботина, Максимов, 2013). Таким образом, общая картина энергетического поведения зарянок на исследуемой территории совпадает с той, что происходит в других регионах Европейского континента (Блюменталь, 1967; Цвей, 2008; Pettersson & Hasselquist, 1985; Karlsson et al., 1988; Cramp, 1992; Ehnbom et al., 1993; Därnhardt & Lindström, 2001; Zimin, 2003).
Заключение
Таким образом, независимо от места расположения стоянки в широтном и долготном направлении, для молодых зярянок сохраняется единая стратегия поведения на миграционных остановках в Европе (Shaub, Jenni, 2000). Молодые птицы не увеличивают свои энергетические резервы перед следующим миграционным броском. Часть популяции осуществляет многодневные остановки, где скорость жиронакопления незначительная, а накопления за весь период не превышают 1 % от средней массы тела в популяции за средний период остановки. Каждые сутки пребывания на остановке приводят к временным всплескам и потерям в весе тела, но накопления жировых отложений не происходит. В наиболее оптимальных условиях оказывается та часть мигрирующих птиц, которые принимают решение о миграционной остановке в медианную дату или раньше. Эта немногочисленная группа является основным резервом для сохранения популяции в сложный период миграции молодых птиц.
Библиография
Ануфриев В. М., Кочанов С. К. Республика Коми // Ключевые орнитологические территории России. М., 2000. Т. 1. С. 633.
Блюменталь Т. И. Изменение энергетических запасов (жирности) у некоторых воробьиных птиц Куршской косы в связи с участием их в миграции // Миграции птиц Прибалтики / Под ред. акад. Б. Е. Быховского. Л.: Наука, 1967. С. 164–202. (Труды Зоол. ин-та АН СССР. Т. 40).
Вепринцев Б. Н., Вепринцева О. Д., Рябицев В. К., Дмитренок М. Г., Букреев С. А. и Гашков С. И. Голоса птиц России. Ч. 1: Европейская Россия, Урал и Западная Сибирь: Звуковой справочник-определитель. Сопроводительный буклет . Екатеринбург: Изд-во Урал. ун-та, 2007. 76 с.
Виноградова Н. В., Дольник В. Р., Ефремов В. Д., Паевский В. А. Определение пола и возраста воробьиных птиц фауны СССР: Справочник / Под ред. В. Д. Ильичева. М.: Наука, 1976. 189 с.
Глухова В. М. Фауна СССР. Насекомые двукрылые. Т. III, вып. 5а. Н. с. № 139. Кровососущие мокрецы родов Culicoides и Forcipomyia (Ceratopogonidae) . Л.: Наука, 1989. 408 с.
Дольник В. Р. Ресурсы энергии и времени у птиц в природе . СПб.: Наука, 1995. 360 с. (Труды Зоол. ин-та РАН. Т. 179).
Зимин В. Б. Зарянка на севере ареала. Т. 2: Линька и миграция . Петрозаводск, 2012. 408 с.
Зимин В. Б., Носков Г. А. Зарянка Erithacus rubecula // Миграции птиц Северо-Запада России. Воробьиные / Под ред. Г. А. Носкова, Т. А. Рымкевич, А. Р. Гагинской. СПб., 2020. С. 271–278.
Накул Г. Л. Об осенней миграции воробьиных птиц в долине р. Сысола в 2015–2017 гг. // Русский орнитологический журнал. 2018. Т. 27. С. 5287–5299.
Панов И. Н., Чернецов Н. С. Миграционная стратегия варакушки (Luscinia svecica) в Восточной Фенноскандии. Сообщение 1: Основные параметры миграционных остановок // Труды Зоол. ин-та РАН. 2010а. Т. 314, № 1. С. 93–104.
Панов И. Н., Чернецов Н. С. Миграционная стратегия варакушки (Luscinia svecica) в Восточной Фенноскандии. Сообщение 2: Реакция на акустические маркёры и выбор биотопа на миграционной остановке // Труды Зоол. ин-та РАН. 2010б. Т. 314, № 2. С. 173–183.
Резвый С. П. Летние миграции зарянки Erithacus rubecula на северо-востоке Ленинградской области // Русский орнитологический журнал. 2011. Т. 20. С. 931–942.
Резвый С. П., Савинич И. Б. О летних миграциях зарянки Erithacus rubecula на юго-восточном берегу Ладожского озера // Русский орнитологический журнал. 2005. Т. 14. С. 935–936.
Рыжановский В. Н. Послегнездовой период жизни варакушки (Luscinia svecica). Распадение выводков и постювенальная линька // Зоологический журнал. 1988. Т. 67, № 1. С. 68–78.
Рыжановский В. Н. Годовой цикл линьки северной варакушки (Luscinia svecica svecica): сроки, полнота и фотопериодические интервалы // Зоологический журнал. 2014. Т. 93, №11. С. 1340–1344. DOI: DOI: 10.7868/S0044513414110087
Савинич И. Б. Зарянка – Erithacus rubecula (L.) // Линька воробьиных птиц северо-запада СССР / Т. А. Рымкевич (ред.). Л.: Изд-во ЛГУ, 1990. С. 47–52.
Субботина Е. Ю., Максимов Ю. В. Особенности сезонной активности представителей массовых родов грибных комаров (Diptera, Sciaroidea) подтаежной зоны Западной Сибири // BioClimLand (Biota, Climate, Landscapes). 2013. № 2. С. 14–16.
Цвей А. Л. Стратегии миграции зарянки (Erithacus rubecula) в восточной Прибалтике : Автореф. дис. … канд. биол. наук. СПб., 2008. 25 с.
Чернецов Н. С. Миграция воробьиных птиц: остановки и полет . М.: Товарищество научных изданий КМК, 2010. 173 с.
Чернецов Н. С., Булюк В. Н., Ктиторов П. С. Роль Джаныбекского оазиса как места миграционных остановок дендрофильных видов воробьиных птиц // Поволжский экологический журнал. 2010. № 2. С. 204–216.
Alerstam T., Lindstrom A. Optimal bird migration, the relative importance of time, energy, and safety // Gwinner E. (Ed.). Bird Migration, Physiology and Ecophysiology. Springer, Heidelberg, 1990. Р. 331–351. DOI: 10.1007/978-3-642-74542-3_22
Bayly N. J., Rosenberg K. V., Gómez C., Hobson K. Habitat choice shapes the spring stopover behaviour of a Nearctic-Neotropical migratory songbird // J. Ornithol. 2019. Vol. 160. P. 377–388. DOI: 10.1007/s10336-019-01624-3
Bulyuk V., Tsvey A. Timing of nocturnal autumn migratory departures in juvenile European robins (Erithacus rubecula) and endogenous and external factors // J. Ornithology. 2006. Vol. 147. P. 298–309. DOI: 10.1007/s10336-005-0046-0
Chernetsov N., Buluk V. N., Ktitorov P. Migration stopovers of passerines in an oasis at the crossroad of the African and Indian flywas // Ringing and Migration. 2007. Vol. 23, № 1. P. 243–251.
Cramp S. The birds of the Western Palaearctic. Vol. 6. Cambridge: Cambridge University Press, 1992. 735 p.
Dänhardt J., Lindström Å. Optimal departure decisions of songbirds from an experimental stopover site and the significance of weather // Anim. Behav. 2001. Vol. 62, No 2. P. 235–243. DOI: 10.1006/anbe.2001.1749
Ehnbom S., Karlsson L., Ylvén R., Åkesson S. A comparison of autumn migration strategies in robins erithacus rubecula at a coastal and an inland site in southern Sweden // Ringing & Migration. 1993. Vol. 14. P. 84–93. DOI: 10.1080/03078698.1993.9674049
Ellegreen H. Stopover ecology of autumn migrating Bluethroats Luscinia s. svecica in relation to age and sex // Ornis Scandinavia. 1991. Vol. 22. P. 340–348. DOI: 10.2307/3676506
Gyimóthy Z., Gyurácz J., Bank L., Bánhidi P., Farkas R., Németh A., Csörgő T. Autumn migration of robins (Erithacus rubecula) in Hungary // Biologia. 2011. Vol. 66. P. 548–555. DOI: 10.2478/s11756-011-0039-9
Kanja W. The autumn migration of the chaffinch Fringilla coelebs over the Baltic coast in Poland // Acta ornithologica. 1981. Vol. XVIII, № 7. Р. 375–418.
Karlsson L., Persson K., Pettersson J. & Walinder G. Fat‐weight relationships and migratory strategies in the Robin Erithacus rubecula at two stop‐over sites in south Sweden // Ringing & Migration. 1988. Vol. 9. P. 160–168. DOI: 10.1080/03078698.1988.9673940
Klaassen M., Kvist A., Lindström Å. Flight costs and fuel composition of a bird migrating in a wind tunnel // Condor. 2000. Vol. 102, No 2. P. 444–451. DOI: 10.1093/condor/102.2.444
Ktitorov P., Tsvey A., Mukhin A. The good and the bad stopover: behaviours of migrant reed warblers at two contrasting sites // Behav. Ecol. Sociobiol. 2010. Vol. 64. P. 1135–1143. DOI: 10.1007/s00265-010-0929-9
Lindström Ǻ. Fuel deposition rates in migrating birds: causes, constraints and consequences // Berthold P., Gwinner E., Sonnenschein E. (Eds.). Bird Migration. Berlin: Springer, 2003. Р. 307–320. DOI: 10.1007/978-3-662-05957-9_21
Newton I. Can conditions experienced during migration limit the population levels of birds? // J. Ornithol. 2006. Vol. 147. P. 146–166. DOI: 10.1007/s10336-006-0058-4.
Pettersson J. & Hasselquist D. Fat deposition and migrationcapacity of Robins Erithacus rebecula and Goldcrests Regulus regulus at Ottenby, Sweden // Ringing & Migration. 1985. Vol. 6. P. 66–76. DOI: 10.1080/03078698.1985.9673859
Remisiewicz M., Nowakowski J. K., Busse P. Migration pattern of Robin (Erithacus rubecula) on the basis of Polish ringing recoveries // Ring. 1997. Vol. 19. P. 3–40.
Salevski V., Bruderer B. The evolution of bird migration – a synthesis // Naturwissenschaften. 2007. Vol. 94 (4). P. 268–279. DOI: 10.1007/s00114-006-0186-y
Schaub M., Jenni L. Body-mass of six long-distance migrant passerine species along the autumn migration route // J. Ornithol. 2000. Vol. 141. P. 441–460. DOI: 10.1007/BF01651574
Sillett T., Holmes R. Variation in survivorship of a migratory songbird throughout its annual cycle // Journal of Animal Ecology. 2002. Vol. 71. P. 296–308. DOI: 10.1046/j.1365-2656.2002.00599.x
Zimin V. B. Body mass variability in juvenile Robins Erithacus rubecula in the Ladoga area // Avian Ecol. Behav. 2003. Vol. 10. P. 1–31.
Благодарности
Автор выражает благодарность сотрудникам Института биологии Коми НЦ УрО РАН А. Б. Наваковскому и А. А. Таскаевой за помощь в освоении статистических методов и обсуждении полученных результатов. Работа выполнена в рамках государственного задания по теме «Распространение, систематика и пространственная организация фауны и населения наземных и водных животных таежных и тундровых экосистем европейского северо-востока России», № гос. регистрации АААА-А17-117112850235-2.

 © 2011 - 2026
© 2011 - 2026