



Введение
Проникновение водных беспозвоночных в несвойственные им ранее местообитания является одной из важнейших экологических проблем (Корнева, 2005). Одной из причин распространения чужеродных видов водных беспозвоночных в самых разнообразных водоемах и водотоках стало развитие аквариумистики (Винарский и др., 2015). Как правило, чужеродные виды, занесенные благодаря любителям-аквариумистам, относятся к пресноводным обитателям тропических водоемов. Некоторые виды-вселенцы, такие как Melanoides sp., Ampullaria sp., отмечены в водоемах севера-западного Приазовья и Европы, Украины (Дегтяренко, Анистратенко, 2011; Сон, 2007). Одним из наиболее значимых экологических факторов, определяющих пространственное распределение и сезонную смену видов, является температура. Температурный диапазон в водоемах-охладителях обеспечивает обитание в них различных в отношении терморезистентности гидробионтов. Известны случаи нахождения жизнеспособных особей тропических моллюсков в зообентосе прудов отстойников АЭС и в сбросных каналах ГРЭС (Яныгина и др., 2009; Нехаев, Палатов, 2016; Силаева, 2017; Bespalaya et al., 2018a, 2018b). В области сброса отработанных вод температуры воды повышены в течение всего года, что и является возможной причиной сохранения жизнедеятельности теплолюбивых моллюсков.
В связи с изменением климатического режима, а также с увеличением антропогенной термальной нагрузки на водоемы значительное число работ посвящено изучению термоустойчивости и возможности температурной адаптации рыб (Капшай, Голованов, 2013). Вопрос о возможности инвазии теплолюбивых моллюсков в водоемы средней полосы России изучен недостаточно, и полученные экспериментальные данные имеют не только теоретический, но и практический интерес. Изучение температурного адаптационного потенциала пресноводного брюхоного моллюска Melanoides granifera к пониженным температурам обусловлено тем, что данный вид обладает рядом качеств, благоприятствующих успешному занятию свободных территорий. Моллюски данного рода имеют широкий ареал обитания – встречаются в Африке, Азии, Австралии. Мелании – грунтовые моллюски, в естественных условиях могут заселять прибрежные участки глубиной до метра и обитать во временных водоемах. Они являются типичными детритофагами, успешно размножаются в диапазоне температур от +18 до 28 ˚С, относительно устойчивы к изменению солености и жесткости воды, дышат жабрами, способны к половому и бесполому размножению, для них характерно живорождение (Leng at el., 1999; Ben-Ami, Heller, 2005; Сафронов, 2001). Предполагается, что за счет способности к партеногенетическому размножению колонизация новых мест обитания моллюсками данного вида при благоприятных условиях среды происходит с достаточно большой скоростью (Pointier at el., 1998). Поэтому целью исследования являлось изучение устойчивости Melanoides granifera к пониженным температурам. Задачи исследования включали определение нижней летальной температуры, выживаемости моллюсков при действии пониженных температур и при снижении влажности грунта.
Материалы
Брюхоногие моллюски Melanoides granifera являются типичными обитателями декоративных аквариумов. Материнскую культуру содержали в аквариуме объемом 20 литров с грунтом, высшей водной растительностью и системой принудительной аэрации и фильтрации. Кормление моллюсков проводили через день сухим кормом для рыб «Тетра». В эксперименте использовали особей одной размерной категории: длина раковины 8.08 ± 0.04 мм, масса моллюска 0.08 ± 0.001 г. Для каждого варианта эксперимента использовали трехкратную повторность. В пластиковые емкости объемом 250 мл добавляли 150 г грунта, доливали отстоянную аэрированную воду и помещали туда по пять особей. Далее емкости с моллюсками переносили в климатическую камеру для поддержания выбранных температур. В контрольном варианте моллюсков помещали в аналогичные емкости при заданной температуре акклимации на необходимый период экспозиции.
Методы
Испытания проводили с использованием климатической камеры (камера постоянной температуры и влажности, модель TH-ME-025). Время экспозиции и скорость понижения температуры в климатической камере задавали автоматически − 0.1 ˚С в минуту до выбранного значения температуры. Скорость понижения температуры выбрана как наиболее оптимальная скорость изменения температуры на основе работ, посвященных изучению верхней летальной температуры (ВЛТ) (Голованов, 2013). Определение нижней летальной температуры (НЛТ) проводили на основе метода хронического летального минимума с последующим расчетом НЛТ. Летальной считают температуру, при которой 50 % животных погибают, а 50 % выживают (Шмидт-Ниельсен, 1982). Определение нижней летальной температуры проводили при температурах акклимации +20˚С и +18˚С, период экспозиции 8 и 168 часов. Для этого на первом этапе находили диапазон летальных температур от LT100 до LT0, а затем графическим методом с помощью пробит-анализа определяли LT50 (Проссер, 1977). Акклимацию моллюсков к соответствующим температурам проводили в течение 7–10 дней.
Изучение изменения выживаемости моллюсков при длительном воздействии пониженных температур проводили при температурах +10 ˚С и +12 ˚С как температурах, приближенных к среднему значению температуры воды в холодный период в водоемах-охладителях АЭС и ТЭС (Яныгина и др., 2009; Гуржапов и др., 2017). Температура акклимации составляла +20 ˚С. При температуре +12 ˚С период экспозиции составлял 48, 72, 96, 120 часов. При температуре +10 ˚С – 8, 24, 48, 72 часа.
Изучение выживаемости Melanoides granifera при снижении влажности грунта проводили при температуре +20 ˚С и относительной влажности воздуха 36–40 %. Влажность и температуру воздуха измеряли термогигрометром «ТКА-ПКМ». В испытаниях использовали 18 емкостей по 5 моллюсков для каждого варианта. Осушение грунта происходило путем естественного испарения воды из открытых емкостей. По мере осушения проводили измерение влажности грунта и оценку выживаемости моллюсков. Влажность грунта рассчитывали по стандартной методике определения влажности грунта весовым методом (Ващенко, 1982). В контрольном варианте объем воды поддерживался на начальном уровне путем добавления отстоянной аэрированной воды. Наблюдения проводили в течение 27 суток.
Проверку выживаемости моллюсков во всех сериях экспериментов проводили по наличию двигательной активности после помещения особей в воду. Гибель особей рассчитывали по среднему значению из трех повторностей по сравнению с контролем.
Статистическая обработка данных проведена в программе Statistica 6.1. Принятый в работе критический уровень значимости р = 0.05.
Результаты
На первом этапе определяли диапазон пессимальных температур от точки холодовой смерти до нижнего значения температуры, при котором выживали все особи. Выявлено, что при температуре акклимации +20 ˚С и 8-часовом воздействии пониженных температур диапазон летальных температур находится от +10 ˚С (LT0, гибель 0 % особей) до +8 ˚С (LT100, гибель 100 % особей, точка холодовой смерти). При акклимации к температуре +18˚С диапазон летальных температур – от +8.5 ˚С (LT0) до +6 ˚С (LT100) (рис. 1).
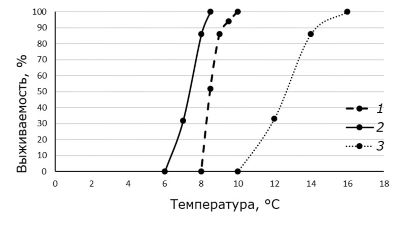
Рис. 1. Диапазон выживаемости моллюсков Melanoides granifera при разных температурах в зависимости от времени воздействия и температуры акклимации: 1 – Твозд. 8 ч, tаккл. +20 °С; 2 – Твозд. 8 ч, tаккл. +18 °С; 3 – Твозд. 168 ч, tаккл.+20 °С
Fig. 1. The range of survival of the mollusk Melanoides granifera at different temperatures depending on the exposure time and acclimation temperature: 1 – Тinf. 8 ч, taccl. +20 °С; 2 – Тinf. 8 ч, taccl. +18 °С; 3 – Тinf. 168 ч, taccl. +20 °С
Как видно из графика, снижение температуры акклимации на 2 °С приводит к увеличению холодоустойчивости моллюсков, смещению значения точки холодовой смерти и увеличению «зоны толерантности». На основе полученных результатов, используя пробит-анализ, установлены значения НЛТ при 8-часовой экспозиции: при температуре акклимации +20 ˚С значение нижней летальной температуры составило +8.4 ˚С, при температуре акклимации +18 ˚С – +7.3 ˚С.
Известно, что выживание организма в зоне сублетальных значений температуры у границ жизнедеятельности зависит и от продолжительности воздействия (Шмидт-Ниельсен, 1982). При экспозиции моллюсков в течение 168 часов диапазон летальных температур определен от +16˚ (LT0) до +10˚C (LT100) Определение НЛТ при более длительном воздействии показало, что моллюски выживали в течение 168 часов при более высоких значениях температуры (см. рис. 1), а нижняя летальная температура составила +13.3 ˚С.
Изучение динамики выживаемости моллюсков при пониженных температурах показало значительное уменьшение числа особей, сохраняющих жизнеспособность по мере увеличения периода воздействия (рис. 2).
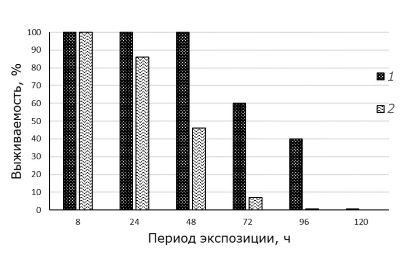
Рис. 2. Выживаемость моллюсков при действии пониженных температур +12 °С (1) и +10 °С (2) в зависимости от периода экспозиции
Fig. 2. Survival of mollusks under the action of low temperatures +12 °С (1) and +10 °С (2) depending on the exposure time
При температуре +12 ˚С через 24 и 48 часов экспозиции сохранялась 100 % выживаемость особей. Гибель 40 % особей отмечена через 72 часа экспозиции (статистически значимые отличия, критерий Mann – Whitney, р = 0.025), 60 % – через 96 часов (статистически значимые отличия, критерий Mann – Whitney, р = 0.036), а через 120 часов при температуре +12 ˚С жизнеспособных особей не обнаружено.
При температуре воздействия +10 ˚C уже через 24 часа отмечена гибель 14 % моллюсков, однако разница в выживаемости не является статистически значимой (критерий Mann – Whitney, р = 0.317). Через 48 часов погибло 54 % особей, а уже через 72 часа отмечена гибель 93 % особей, что является статистически значимыми отличиями (критерий Mann – Whitney, р = 0.036 и р = 0.033 соответственно).
Мелании могут обитать на мелководье в прибрежном грунте (Leng, 1999), где существует вероятность выноса моллюсков в зону выше уреза воды. Поэтому изучение их выживаемости при пересыхании грунта имеет не только теоретический, но и практический интерес. В нашем исследовании выявлено, что при осушении грунта путем естественного испарения воды выживаемость моллюсков достоверно снижается только при влажности грунта менее 2 % (критерий Mann – Whitney, р = 0.046) (рис. 3).
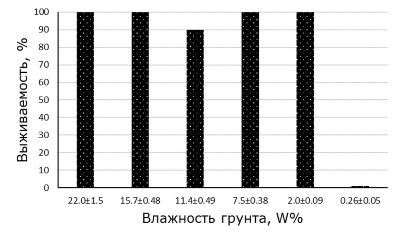
Рис. 3. Выживаемость моллюсков Melanoides granifera при разной влажности грунта
Fig. 3 Survival of mollusks Melanoides granifera at different soil moisture
В вариантах с влажностью грунта 7.5 ± 0.38, 11.4 ± 0.48 и 15.7 ± 0.48 после его обводнения, помимо взрослых особей, отмечено наличие жизнеспособной молоди моллюсков.
С учетом результатов предыдущих исследований по определению выживаемости моллюсков при понижении влажности грунта и воздействии пониженных температур были выбраны температура +10 ˚С и значение влажности грунта, соответствующее 20 %, для изучения возможности выживания моллюсков в зоне выше уреза воды. Выявлено, что в осушенном грунте при воздействии пониженных температур в течение 168 часов выживаемость моллюсков в опыте снижается на 34 %, что является статистически значимым отличием по отношению к контролю (критерий Mann – Whitney, р = 0.033) (рис. 4).
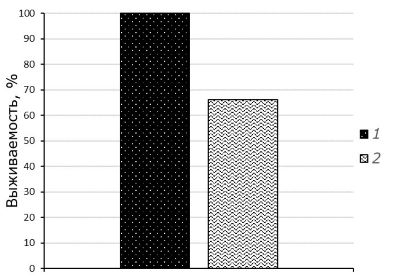
Рис. 4. Выживаемость Melanoides granifera при влажности грунта 20 % при температуре среды +20 °С (1) и +10 °С (2) в течение 168 часов
Fig. 4. Survival of Melanoides granifera at soil moisture of 20 % and an ambient temperature of + 20 °C (1) and + 10 °C (2) for 168 hours
Обсуждение
НЛТ при разных температурах акклимации и времени воздействия определяли для оценки устойчивости Melanoides granifera к пониженным температурам, как к основному, определяющему их выживаемость фактору. Летальная температура является одним из важнейших температурных критериев жизнедеятельности пойкилотермных животных. По значению НЛТ можно судить о нижней границе существования вида и его адаптационных возможностях при действии естественных и антропогенных факторов среды (Голованов и др., 2012; Капшай, Голованов, 2013). Для инвазионных видов этот фактор может стать определяющим экспансию.
Полученные значения НЛТ показывают, что в естественных водоемах северо-западных и центральных регионов Европейской части России выживание и распространение меланий будет ограничиваться температурным фактором, т. к. температура воды в холодный период в них снижается до 0 °С. Даже с учетом понижения температуры акклимации уже кратковременное воздействие приводит к гибели 50 % особей при температурах, не являющихся летальными для беспозвоночных водоемов нашего региона. С увеличением времени воздействия до 168 часов значения НЛТ возрастают до температур, свойственных летне-осенним показаниям. Кроме того, при длительном воздействии пониженных температур отмечены изменения и в поведении моллюсков. Так, по окончании экспозиции обнаружено, что в контрольной группе моллюски зарывались в грунт, а в опытных группах большинство особей находились на поверхности грунта. Возможно, это связано с нарушением цепи поведенческих актов, присущих моллюскам в оптимальных условиях. Изучение динамики выживаемости моллюсков при действии пониженных температур показало, что количество жизнеспособных особей Melanoides granifera снижается по мере уменьшения температуры среды, и чем ниже температура среды, тем меньший процент особей сохраняет жизнеспособность за один и тот же промежуток времени. Причем моллюски погибали уже через 120 часов при температурах, значительно превышающих значения температуры воды в естественных водоемах в зимний период. Таким образом, для сохранения популяции данного вида моллюсков даже в водоемах-охладителях требуются относительно высокие значения температур, которые достигаются, как правило, только в зоне сброса отработанных вод.
Изучение выживаемости меланий при осушении грунта показало, что они сохраняют жизнеспособность при снижении влажности грунта до 2 %. Возможно, при осушении грунта моллюски, зарывшись в грунт, переходят в состояние ангидробиоза, что способствует их выживанию. Известно, что в состоянии биологического покоя, в частности ангидробиоза, беспозвоночные способны переносить неблагоприятные условия среды на протяжении длительного периода (Алексеев, 2010). Интересным фактом стало наличие в осушенном грунте после его обводнения жизнеспособной молоди меланий. Вероятно, молодь может сохранять жизнеспособность в условиях пониженной влажности грунта, находясь в выводковой камере материнской особи. Это может являться важной приспособительной реакцией для сохранения численности популяции. Выживание именно молодых особей в периоды безводной зимовки или летнего пересыхания известно для пресноводных моллюсков наших широт (Митропольский, 1978).
Изучение выживаемости моллюсков в осушенном грунте при действии пониженных температур показало, что снижение температуры среды выступает дополнительной функциональной нагрузкой, способствующей снижению жизнеспособности особей. Однако ранее нами было показано, что при данной температуре в обводненном грунте моллюски погибали через 168 часов. Следовательно, при действии пониженных температур вероятность выживания моллюсков в обсыхающем грунте выше, чем в обводненном, что, вероятно, связано с их переходом в состояние биологического покоя. Таким образом, показанная ранее способность моллюсков данного вида сохранять жизнеспособность в осушенном грунте при нормальных температурах может послужить одной из преадаптаций, способствующей их распространению в водоемах с благоприятным терморежимом.
Заключение
В осенний период устойчивый переход среднесуточных температур воздуха через 0 ºC влечет за собой снижение температуры воды до 0 ºC и начало ледовых явлений на водоемах и водотоках (Бузин, Зиновьев, 2009). В водоемах-охладителях ГРЭС и прудах-отстойниках АЭС температура воды в зонах постоянного сильного подогрева и зонах сброса может превышать естественные значения в зимний период на 14–16 ˚С, а в летний – на 11–14 ˚С (Болдаков, 2003; Гуржапов и др., 2017), что создает вероятность выживания и размножения в них данного вида моллюсков. Для теплолюбивых моллюсков зоны сброса подогретых вод ГРЭС и АЭС, имеющих специфический гидротермический режим, могут стать своеобразным температурным рефугиумом. Интродукция данного вида моллюсков может представлять опасность для биологического разнообразия коренных видов в водных объектах и привести к появлению новых паразитарных заболеваний, поскольку представители данного рода являются промежуточными хозяевами для ряда паразитов (López-López et al., 2009; Pointier et al., 1998; McKoy et al., 2011). Полученные данные указывают на примерное расположение границы нижнего термального полигона, и, учитывая термофильность вида, широкое распространение его в естественных водоемах северных районов маловероятно.
Выводы
- Нижняя летальная температура для моллюсков Melanoides granifera при 8-часовой экспозиции и температуре акклимации +20 ˚С составила +8.4 ˚С, при температуре акклимации +18 ˚С − +7.3 ˚С.
- Нижняя летальная температура для моллюсков Melanoides granifera при 168-часовой экспозиции и температуре акклимации +20 ˚С составила +13.3 ˚С.
- Выживаемость Melanoides granifera при осушении грунта зависит от степени снижения влажности грунта. Моллюски погибают при влажности грунта менее 2 %.
- Выживаемость моллюсков зависит от температуры среды и периода воздействия. Чем дольше моллюски находятся при пониженных температурах, тем меньше их выживаемость. При более высоких значениях температуры среды увеличивается время сохранения жизнеспособности моллюсками.
- При влажности грунта 20 % выживаемость моллюсков снижается при действии пониженных температур.
Библиография
Алексеев В. Р. Физиологические и молекулярно-генетические основы механизма биологического покоя у водных беспозвоночных // Онтогенез. 2010. Т. 41. № 2. С. 83–93.
Болдаков А. М. Влияние подогретых вод Костромской ГРЭС на поведение и пространственное распределение рыб : Дис. ... канд. биол. наук. Кострома, 2003. 197 с.
Бузин В. А., Зиновьев А. Т. Ледовые процессы и явления на реках и водохранилищах. Методы математического моделирования и опыт их реализации для практических целей . Барнаул: Изд-во ООО «Пять плюс», 2009. 168 с.
Ващенко И. М. Практикум по основам сельского хозяйства . М.: Просвещение, 1982. 399 с.
Винарский М. В., Андреев Н. И., Андреева С. И., Казанцев И. Е., Каримов А. В., Лазуткина Е. А. Чужеродные виды моллюсков в водных экосистемах Западной Сибири: Обзор // Российский журнал биологических инвазий. 2015. № 2. С. 2–18.
Голованов В. К., Смирнов А. К., Капшай Д. С. Окончательно избираемые температуры и верхние летальные температуры молоди некоторых видов пресноводных рыб // Труды Карельского научного центра РАН. 2012. № 2. С. 70–75.
Голованов В. К. Температурные критерии жизнедеятельности пресноводных рыб . М.: Полиграф-Плюс, 2013. 300 с.
Гуржапов Б. Ю., Цыдыпов Б. З., Андреев С. Г., Аюржанаев А. А., Содномов Б. В., Батоцыренов Э. А., Павлов И. А., Ширеторова В. Г., Ульзетуева И. Д., Раднаева Л. Д., Гармаев Е. Ж. Оценка влияния сбросов Гусиноозерской ГРЭС на термический и гидрохимический режим оз. Гусиное // Экология водоемов-охладителей энергетических станций: Сб. материалов Всерос. научно-практ. конф. с междунар. участием. Чита: ЗабГУ, 2017. С. 57–65.
Дегтяренко Е. В., Анистратенко В. В. Моллюски континентальных водоемов Северо-западного Приазовья: фаунистический обзор с замечаниями по распространению и экологии // Збįрникпраць Зоологичįчного музею. 2011. № 42. С. 13–57.
Капшай Д. С., Голованов В. К. Верхняя летальная температура у молоди теплолюбивых видов рыб в зависимости от температуры акклимации // Труды Карельского научного центра РАН. 2013. № 3. С. 185–189.
Корнева Л. Г. Современные инвазии планктонных водорослей // Чужеродные виды в Голарктике (Борок-2). Рыбинск: ОАО «Рыбинский печатный дом», 2005. С. 47–49.
Митропольский В. И. Наблюдения над способностью моллюсков к перенесению высыхания и промерзания в прибрежье Рыбинского водохранилища // Фауна беспозвоночных и условия воспроизводства рыб в прибрежной зоне Верхне-Волжских водохранилищ: Сб. тр. 1978. Вып. 39 (42). С. 46–48.
Нехаев И. О., Палатов Д. М. От моря Черного к морю Белому: первая находка инвазийного моллюска Physella acuta на крайнем севере Европы // Российский журнал биологических инвазий. 2016. № 3. С. 61–65.
Проссер Л. Сравнительная физиология животных . М.: Мир, 1977. Т. 2. 571 с.
Сафронов В. Грунтовые улитки – мелании // Аквариум. 2001. № 3. С. 37–40.
Силаева А. А. Особенности структуры зообентоса техноэкосистем АЭС и ТЭС // Экология водоемов-охладителей энергетических станций: Сб. материалов Всерос. научно-практ. конф. с междунар. участием. Чита: ЗабГУ, 2017. С. 243–251.
Сон М. О. Моллюски-вселенцы в пресных и солоноватых водах Северного Причерноморья . Одесса: Друк, 2007. 132 с.
Шмидт-Ниельсен К. Физиология животных. Приспособление и среда . Книга 1 / Пер. с англ. М. Д. Гроздовой, Г. И. Рожковой; Под ред. Е. М. Крепса. М.: Мир, 1982. 426 с.
Яныгина Л. В., Кириллов В. В., Зарубина Е. Ю. Виды-вселенцы в биоценозе водоема-охладителя Беловской ГРЭС (юг Западной Сибири) // Российский журнал биологических инвазий. 2009. Т. 2. № 2. С. 60–68.
Ben-Ami F., Heller J. Spatial and temporal patterns of parthenogenesis and parasitism in the freshwater snail Melanoides tuberculata // J. Evol. Biol. 2005. Vol. 15. P. 138–146.
Bespalaya Y. V., Bolotov I. N., Aksenova O. V., Kondakov A. V., Gofarov M. Y., Laenko T. M., Sokolova S. E., Shevchenko A. R., Travina O. V. Aliens are moving to the Arctic frontiers: an integrative approach reveals selective expansion of androgenic hybrid Corbicula lineages towards the North of Russia // Biol. Invasions. 2018a. Vol. 20. P. 2227–2243.
Bespalaya Yu. V., Bolotov I. N., Aksenova O. V., Gofarov M. Yu., Kondakov A. V., Vikhrev I. V., Vinarski M. V. DNA barcoding reveals joint invasion of two cryptic Sinanodonta mussel species (Bivalvia: Unionidae) into the largest Siberian river. Limnologica (in press) // Limnologica. 2018b. Vol. 69. P. 94–102.
Leng M. J., Lamb A. L., Lam H. F., Telford R. J. Palaeoclimatic implications of isotopic data from modern and early Holocene shells of the freshwater snail Melanoides tuberculata, fromlakes in the Ethiopian Rift Valley // Journal of Paleolimnology. 1999. Vol. 21. Р. 97–106.
López-López E., Sedeño-Díaz J. E., Vega P. T., Oliveros E. Invasive mollusks Tarebia granifera Lamarck, 1822 and Corbicula fluminea Müller, 1774 in the Tuxpam and Tecolutla rivers, Mexico: spatial and seasonal distribution patterns // Aquatic Invasions. 2009. Vol. 4. P. 435–450.
McKoy S. A., Hyslop E. J., Robinson R. D. Associations between two trematode parasites, an ectosymbiotic annelid, and Thiara (Taberia) granifera (Gastropoda) in Jamaica // Journal of parasitology. 2011. Vol. 97. P. 828–832.
Pointier J. P., Samadi S., Jarne P., Delay B. Introduction and spread of Thiara granifera (Lamarck, 1822) in Martinique, French West Indies // Biodiversity and Conservation. 1998. Vol. 7. P. 1277–1290.

 © 2011 - 2026
© 2011 - 2026